
Abstract
Field transfer experiments of periphytic diatom assemblages developed on artificial substrates were set up to assess the responses of those communities to environmental disturbances. The glass slides were positioned for colonization at the relatively unpolluted site (Red, in the Red River) and at the heavily polluted site (TL, in the To Lich River) in the beginning of the experiment. After a period of 2 weeks, the colonized glass slides were concomitantly transferred from the unpolluted Red site to the heavily polluted TL site and to the moderate polluted site (NT2, in the Nhue River) and, conversely, from the TL site to the Red site, and then to the NT2 site. The responses and the adapting capacity of periphytic diatom communities to environmental changes were assessed through the cell density, diversity index, species richness, taxonomic composition, and diatom indices after 2 and 4 weeks of transfer periods. For all transfers except for the transfer from the Red to the TL site in which the growth inhibition of diatom cells was found, the diatom density significantly increased until the end of the experiment. Thus, the diatom communities have expressed their pollution tolerance or sensitivities by changing their composition to adapt themselves to environmental changes. Characteristic species of the Red site (Gyrosigma scalproides, Navicula recens) were replaced by Nitzschia palea, Nitzschia umbonata, Aulacoseira granulate typical species of the NT2 site, in the biofilm transferred from the Red site to the NT2 site. The relative abundances of typical diatom species of the Red site proliferated in the biofilm transferred from the TL site to the Red site. The replacement of periphytic diatom communities appeared after the transfer from the second week at the different sites. The slow shift of the species towards the typical species at the TL site could result from the organized structure of diatoms within biofilm before the transfer from the Red site to the TL site. The shifts in values of the Index of Specific Polluosensitivity and Diatom Assemblage Index to organic pollution throughout the experiment indicated the clear sensitivity of these indices to water quality changes.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Anthropogenic activities play a major role in increasing nutrients and a large number of chemicals in surface water, causing unfavorable changes in aquatic ecological systems. The biological integrity of an aquatic ecosystem depends upon its physical and chemical components, which is also controlled by the local geology together with the land use activities within the watershed. In many streams and rivers, periphytic algae being primary producers, are considered to be a major source of energy to aquatic food webs (McCormick and Stevenson 1998). Because they are attached to substrates, their characteristics are affected by physical, chemical, and biological disturbances occurring in streams or rivers during the time in which they develop. Moreover, periphytic algae integrate ecological variations over time and can provide the information about changes in the water quality (Lowe and Pan 1996; Gold et al. 2002). The periphytic algae, in general, and diatoms, in particular, have been widely used as the organisms for monitoring and assessing the water quality owing to their wide distribution and well-studied ecology (Lange-Bertalot 1979; Potapova and Charles 2005; Centis et al. 2010). The diatoms are considered to be the excellent biological indicators for many types of pollution in aquatic systems such as organic pollution (Descy and Coste 1991), acidification, metallic pollution (Gold et al. 2003; Cattaneo et al. 2004; Morin et al. 2007), and eutrophication (Dixit et al. 1992). The analysis of diatom communities can be used to discriminate the impacts of organic pollution and inorganic nutrient enrichment such as treated urban wastewater and diffuse nutrient sources from farmland (Rott et al. 1998). The diatom indices based on selected sensitivities or tolerance of diatom species to organic pollution or acidity are being routinely and widely used in many European, North American, and Asian countries (Watanabe et al. 1986; Whitton and Rott 1996; Lavoie et al. 2006) for the assessment of the river water quality.
It has been well documented that multi-sources of pollution greatly affected the structure and function of periphytic algal communities. These effects included reduced photosynthetic ability (Soldo and Behra 2000; Barranguet et al. 2000), reduced growth rate (Gibson and Fitzsimons 1991; Genter 1996), the interruption of cell division, and deformation of diatom frustules (Gold et al. 2003; Cattaneo et al. 2004). At a cellular level, algae may tolerate pollutant stress by showing a decreased number of binding sites at the cell surface, the physiological development of exclusion mechanisms, genetic adaptation, morphological changes, and internal detoxifying mechanisms (Genter 1996). At a community level, the periphytic algae increase their tolerance by shifting their composition to more tolerant species when they are exposed to pollution (Kasai 1999), which can result in structural changes such as decreased species diversity and richness (Genter 1996). Many manipulative stream experiments have been extensively performed in both natural and laboratory conditions to assess the impacts of short- and long-term environmental changes on the periphytic algal communities. The effect of long-term copper exposure on the structure of periphytic communities was monitored in outdoor and indoor flows by Soldo and Behra (2000). Barranguet et al. (2000) used indoor experiment to assess the short-term metal effect on communities’ tolerance in photosynthetic process. Pan et al. (2000) used mesocosms to study the structural and functional changes of periphytic algal assemblages due to the increased phosphorus loading.
Up to now, few transfer experiments have been performed to assess the alteration of periphytic diatom assemblages caused by changes in water quality. Some earlier studies have suggested that the translocation of periphytic diatom communities developed on artificial substrates among different pollution sites was a suitable method to assess, in situ, the effects of metallic pollution (Ivorra et al. 1999; Gold et al. 2002; Tolcach and Gomez 2002). The translocation experiments showed variation in the response time of periphytic communities to simulated water quality improvement and degradation in running waters. Rimet et al. (2005) assessed the response of diatoms to water quality changes after the transfer of diatom-dominated biofilms from polluted sites to an unpolluted site. They noted that 40 to 60 days were necessary for the diatom indices calculated with the transferred communities to be the same as the indices calculated with the reference communities. Lavoie et al. (2008) suggested that the integration period of diatom indices varied from 2 to 5 weeks as a function of trophic status of the ecosystems. Differences or changes in the biomass, productivity, and structure of periphyton communities can provide a sensitive measure of the trophic and wholeness status of streams and rivers. In this study, we aimed (1) to investigate the response of the periphytic diatom communities on environmental changes via the transfers of early stages of periphytic diatom communities developed on artificial substrates from the reference site to the moderately and heavily polluted sites, and reversely (2) to estimate the time needed for diatom communities to adapt to new conditions.
Materials and methods
Study area and design
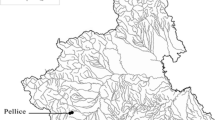
The sampling area is located in the Red River delta (North Vietnam) and includes three rivers: the Red, the Nhue, and the To Lich rivers (Fig. 1). The Red River is the largest one and plays an important role in the economic and social life in the North of Vietnam.

Location of sampling stations (TPd Thuy Phuong dam; TLd Thanh Liet dam)
The Nhue River, a tributary of the Red River, diverges from it at the Thuy Phuong dam, runs through the Hanoi and Hadong cities and Hanam province via several dams, and finally flows into the Day River. The length of Nhue River is 74 km and covers a total basin area of 107,503 ha with 75% cultivated area. The topography of the Nhue basin is sloping down from the North to the South. The soil textures of the Nhue River change with topography from sandy loam to clayey loam in its lower part (Kono and Doan 1995; Trinh 2003). There are two flow conditions in the Nhue River in relation with precipitations (the free-flow regime) and with opening or closing the dams (the close-dam regime), which regulate all over the year water current parameters in the Nhue River. During the dry season, dams are opened to get water from the Red River to irrigate winter/spring crops, and the currently measured value of the water inflow is 30 m3 s−1. During the rainy season, the water inflows are notably lower as the dams are closed to protect Hanoi and other areas of the Nhue basin from floods. At about 20 km downstream from its confluence with the Red River, the Nhue River receives the water discharge from the To Lich River through the Thanh Liet dam.
The To Lich River, which is 14 km long, originates from the West Lake (in Hanoi city) and flows through Hanoi city. Barely, the untreated wastewater of the Hanoi city is directly discharged into the To Lich River (via the Thanh Liet dam). The Nhue and To Lich rivers situated in the densely populated region (about 1,000 inhab/km2). The wastewater sources of the Nhue and To Lich systems originate from four main supplies: (1) 335,000 m3 d−1 of the industrial and municipal wastewater from Hanoi city; (2) disposal of the municipal wastewater along the river bank; (3) the wastewater from numerous traditional production villages along the river banks (craft villages producing papers, food, leather, texture, timber, etc.); (4) agricultural run-off (fertilizers and pesticides) and soil erosion from agricultural cultivated lands along the riverbank (Nguyen 2001). The water quality of the Nhue River is influenced by the agricultural wastewater from the upper basin and by the domestic and industrial wastewater from the downstream area (after receiving the To Lich River) whereas the To Lich River is strongly influenced by the domestic and industrial wastewater from the Hanoi city (Trinh 2003). However, the Nhue and To Lich rivers have been utilized as the major drainage and irrigation canals for the surroundings of the Hanoi areas.
The climate in the studied area is tropical and subtropical monsoon which leads to an average annual rainfall of 1,800–2,000 mm and 80% of which occur during the rainy season from June to October. The dry season lasts from November to April–May. The relative humidity is very high throughout the year with an annual value of 84.5% (Kurosawa et al. 2004). Within this area, the three study sites are: (1) the relatively unpolluted reference site (Red) in the Red River, which is situated at the Son Tay gauging station (Son Tay city, Hanoi area), about 8 km upstream from the Nhue River and Hanoi city, shows low nutrients and metal concentrations (Le 2005); (2) the heavily polluted site (TL) is located before the Thanh Liet dam (TL dam), downstream of the To Lich River and characterized by low dissolved oxygen, high nutrients and metal concentrations with black, foul-smelling water; (3) the moderately polluted site (NT2) is positioned in the Nhue River about 7 km downstream of its confluence with the To Lich River.
Field sampling
The experiment was performed during the dry season from (January to 20 February 2005. During this period, the rivers were connected; dams were opened to normally irrigate the downstream basin for agricultural purposes. However, in that dry season, the water level of the Red River was low, so the water supply to the Nhue River was quite limited. At both sites, Red and TL, one set of two plastic baskets equipped with floaters were immersed in water column parallel to the current at a depth of 15–20 cm under the surface and tied to the bank with a rope (Gold et al. 2002; Morin et al. 2009). In each plastic basket, 18 glass slides were placed separately and vertically to be used as the artificial substrata for algal attachment (30 × 18 cm; 1,080 cm2 surface for both sides). One set of plastic baskets was left at the Red site and the other one at the TL site in the beginning of the experiment for a period of 2 weeks to allow the development of biofilms on the glass slides. After 2 weeks of colonization at the Red site, one of the two plastic baskets containing glass slides was transferred to the heavily polluted site (TL); the other basket was moved to the moderate site (NT2). The same procedures were followed at the TL site, one plastic basket containing glass slides transferred to the reference site (Red), and one basket to the moderate site (NT2). During transportation, glass slides were kept immersed in their initial river water within a cool container box (1–2 h travel time). After transfer, the glass slides were left in their new locations for an additional period of 4 weeks. The periphytic diatoms developed on the glass slides were sampled before transfer at the second week of colonization (W2), after transfer at the fourth week (W4), and at the sixth week (W6). On the day of sampling at each site three glass slides, considered as the independent samples, were randomly removed from each plastic basket. Glass slides were then thoroughly rinsed with filtered river water. The biofilms were collected from the glass slides by using a nylon brush and then washed and diluted in a known volume (100 or 200 mL) of water depending on the biofilm thickness. In order to determine the dry weight of biofilms, a known volume of samples was stored in labeled polyethylene bottles and placed in a cool box (4–10°C) during the transportation to laboratory. The remainders of the natural biofilm samples were preserved in the labelled glass bottles with 5% formalin solution for the delayed identification of diatom composition.
Temperature, dissolved oxygen, conductivity, and pH of water were measured in situ during the whole experiment by using multi-parameter sensors (Model WQC-22 A, TOA). Water samples were taken near the river surface (30 cm below the surface) for nutrient determination and stored in an iced box (4–10°C) during sampling. The samples were filtered with Whatman GF/C filter (glass micro-fiber filters 0.45 μm) using hand pump filtration. The analysis of phosphate, ammonia, nitrate, and nitrite concentrations in water samples followed the methods of the American Public Health Association (APHA 1995). The analysis was performed at the Institute of Natural Product Chemistry (INPC, VAST).
Laboratory analysis
Diatom preparation
After homogenization, a 2-mL aliquot of each diatom sample from each site was digested in concentrated hydrogen peroxide (30%) and hydrochloric acid (35%) to remove organic matter and dissolved calcium carbonates, rinsed several times, and then diluted with deionized water. The cleaned diatom frustules were then mounted on a microscope glass slide using Naphrax, high-resolution mounting medium (Brunel Microscopes Ltd, UK; RI = 1.74; Gold et al. 2002; EN 13946 2003; EN 14407 2004). The diatoms were identified with a Leitz DMRD light microscope at ×1,000 magnification. Approximately 400 valves were identified at species level from each slide of the three replicates. The Süßwasserflora nomenclature (Krammer and Lange-Bertalot 1986–1991) was used as the references for the diatom taxonomy. The relative abundance of diatom species (in percentage) was estimated. The species richness (S) was calculated and the biological diversity was estimated through the Shannon–Weaver diversity index (H’). The diatom indices (IPS and DAIPo index) were applied in order to classify the water quality of each sampling site.
A Nageotte counting chamber (Marienfeld, Germany) was used to estimate the living diatom density (cell with chloroplast) in each sample by counting the total number of diatoms in 30 fields (1.25 μL each, 0.5 mm depth) under a light microscope (Olympus BX ×50) at ×200 magnification. Data were expressed in cells per area unit of glass slide (number diatom cells cm−²).
Biofilm dry weight determination
A known volume of homogenous biofilm suspension sample was filtered with a manual aspiration pump, through a previously weighed filter membrane (glass micro-fiber filters 0.45 μm) for determination of biofilm dry weight (dw) (values expressed as mg dw cm−2). The filters covered with drained biofilms were then thoroughly dried at 60°C for 48 h in incubation tubes, and dry weights were measured afterwards.
Data analysis
The diatom indices, IPS (Index of Specific Polluosensitivity; Cemagref 1982) and DAIPo (Diatom Assemblage Index to organic Pollution; Watanabe et al. 1986), were calculated using the OMNIDIA software (Lecointe et al. 1993). These diatom indices were transformed to range from 1 to 20 to be comparable. In order to study the significant differences in the diatom density, the structure of diatom communities (species richness, diversity index) and the diatom indices between the local and transferred diatom communities, we performed the ANOVA method using the STATISTICA software (StatSoft 2004). For statistical results, a probability of p < 0.05 was considered significant. The taxonomic differences between the sites were displayed using a principal component analysis (PCA) with PC-ORD Software (McCune and Mefford 1999). The PCA was performed based on the relative abundances of 53 species among 277 identified diatom species, which were selected because of their highest cumulative relative abundances within local and transferred diatom communities collected through out the experiment.
Results
Physical and chemical characteristics of sites
The physical and chemical characteristics of measured water at three sampling sites Red, NT2, and TL are shown in Table 1. Most of the parameters did not show significant variations at each site throughout the experiment. At all sites, pH varied from pH 6.4 to 7.5, showing quite neutral pH. The water temperature values ranged from 18.2°C to 21.0°C, with the lowest value observed at the Red site where reversely showing the highest dissolved oxygen value. The lower mean values of dissolved oxygen were recorded at the NT2 and TL sites (3.3 mg L−1 and 2.1 mg L−1, respectively). The mean conductivity value was approximately four times higher at the TL and NT2 sites than at the Red site. The higher nutrient (N, P) pollution levels of these two sites were also observed in comparison with those of the Red site. NH4–N and PO4–P were differentiated from other parameters by a sharp decrease of NH4–N values at the end of experiment at the NT2 and a clear increase of PO4-P at the TL, both associated with an increase of dissolved oxygen values. The water quality of the Red, the Nhue, and To Lich Rivers, which were observed monthly at the same sampling sites for the period 2002–2003, are well described in several studies (Trinh 2003; Le 2005). These results showed that the water quality of the To Lich River was strongly polluted while the Red River at the Red site (before entering the urbanized region of the delta) was much less impacted by the domestic and industrial activities. The Nhue River at NT2, downstream part after the confluence with the To Lich River, was in the middle pollution level in comparison with the Red and the To Lich Rivers.
Dry weight of biofilms
During the first 2-week colonization period, the mean value of biofilm dry weight was higher at the Red site than at the TL site by a factor of 29 (Fig. 2). However, the biofilms transferred from the Red to the TL site and from the Red to the NT2 site (Fig. 2a) did not follow the same pattern of development. A continuous increased development was observed in the biofilms transferred from the Red to the NT2 site, and the highest values of dry weights were then recorded with 6.2 mg cm−2 at W4 and 7.2 mg cm−2 at W6. In contrast, the biofilms transferred from the Red to the TL site maintained an average biomass of 2 mg cm−2 during the last 4-week period after transfer. The very thin biofilms initially constituted at the TL site grew thicker after the transfer to their new location (p < 0.05) with a stronger development at the NT2 site (dry weight average value, 5.3 ± 0.6 mg cm−2 at W4 and 4.6 ± 0.4 mg cm−2 at W6) than at the Red site (dry weight average value, 2.5 ± 0.26 mg cm−2 at W4 and 2.8 ± 0.5 mg cm−2 at W6) but without reaching the outstanding development observed at the NT2 after the transfer from the Red to the NT2 site (Fig. 2).

Dry weight of biofilms collected from the Red and TL at week 2 (W2) (during initial colonization site before transfer), then after transfer to their new locations (Red, NT2, or TL): week 4 (W4) and week 6 (W6). Box plot: mean value (n = 3) with its confidence interval at 95% denoted by the two horizontal lines outside the box
Diatom density
The quantitative characterization of diatom community assemblages was reflected by the numeration of diatom cells. The diatom densities developed on the glass substrates before (W2) and after the transfer (W4 and W6) are illustrated in Fig. 3 for the three sites. Figure 3a presents the diatom density of biofilms previously colonized in the Red site during 2 weeks, then transferred to either the TL site or the NT2 site for 4 extra weeks. Figure 3b shows the same development, but for the biofilms, previously developed at the TL site then transferred to the Red or the NT2 site. The total diatom density at the Red site showed 20 times more cells during the two initial week period of colonisation than at the TL site (4,928 ± 446 cells cm−2; 240 ± 19 cells cm−2, respectively). A yellow-brown-colored layer including detritus, algae, bacteria, and suspended particular matters covered glass substrates at the Red site after 2 weeks, whereas a thin black-colored layer was visible on its substrates at the TL site. After transfer of the glass substrata from the Red site to the NT2 and TL sites, the development of diatoms at these two sites were different (Fig. 3a). The densities at the TL site ranged then from 3,595 ± 351 to 6,248 ± 182 cells cm−2, showing low development of diatom communities. Meanwhile, at the NT2 site, the development of diatom densities after transfer from the Red site increased significantly (p < 0.05) until the end of the experiment. For the glass substrates transferred from the TL site to the Red and NT2 sites, the diatom densities on substrates grew a lot at their new locations and reached the maximum values of 31,539 ± 1511 cells cm−2 at the NT2 site and of 58,034 ± 3401 cells cm−2 at the Red site during the last week (Fig. 3b). Despite the difference in the diatom densities from the initial period of colonization in the Red or TL site, the diatom growth at the NT2 site after both translocations showed similar trend of the development, ranging from 11.8 ± 2.1 cells cm−2 at W4 to 33.8 ± 6.3 cells cm−2 at the W6 for the transferred diatom communities from the Red site to the NT2 site and from 15.2 ± 1.6 cells cm−2 at W4 and 31.5 ± 1.5 cells cm−2 at the W6 for the transferred diatom communities from the TL site to the NT2 site. Qualitative observations of the biofilm at NT2 after 4 weeks of transfer revealed the development of a thick, yellow-brown and mucilaginous coat growing over a black layer of material. Significant differences in the diatom densities between sites and colonization durations were clearly observed (p < 0.05).

Diatom densities on artificial substrates (cells cm−2) collected from the three stations Red and TL at the week 2 (W2) of the experiment (during initial colonization on site before transfer), then after transfer to their new locations (Red, NT2, or TL): at week 4 (W4) and week 6 (W6). Box plot: mean value (n = 3) with its confidence interval at 95% denoted by the two horizontal lines outside the box
Species richness, diversity of diatom communities, and diatom indices
The species richness (S), the Shannon–Weaver index (H’) (Shannon and Weaver 1963), and the diatom indices occurring during the experiment are shown in Table 2. A significant difference was noted between the communities previously developed at the Red and TL sites during the first 2 weeks, with 22% more species initially developed at the Red site than at the TL site. On the other hand, no significant difference was detected in S between the local diatom communities (Red, TL) at W2 and the transferred diatom communities during the last 4 weeks of the experiment, although S seemed to decrease for the environmental condition at the Red to the NT2 and to increase for both transfers from TL when compared with week-2 values. Concerning the Shannon diversity index (H’), only transferred diatom communities from the Red site to the NT2 site showed a significant difference during the course of the experiment, when compared with the local diatom communities (p < 0.05). Both diatom indices (IPS and DAIPo) revealed fairly similar profiles (Table 2). After transfer from the Red site, the IPS and DAIPo indices gradually and significantly (p < 0.05) decreased with exposition duration to reach the minimum values at W6 (4.5 and 7.3 at the NT2 site; 7.9 and 8.6 at the TL site, respectively). Table 2 also shows a considerable increase of the indices (mainly IPS) after transfer from the TL site to the Red site, although they did not reach the high values obtained at the original site (Red) during the first period of colonization. After transfer from the TL site to the NT2 site, diatom indices also increased at W4 but showed different patterns with a decrease during the last week of the experiment for IPS and stabilization for DAIPo.
Diatom composition
The composition of diatom communities of the local and transferred diatom assemblages, which colonized the substrates through out the experiment, is presented by the relative abundances of seven diatom species (the mean relative abundances ≥8%; Fig. 4a, b). The diatom community of the Red site at W2 was dominated by Gyrosigma scalproides (GSCA) and Navicula recens (NRCS), ranging from 14.1% to 12.5%, respectively (Fig. 4a). After the transfer to the NT2 site, the composition of the transferred diatom communities was modified firstly with high proportion of G. scalproides (GSCA) and N. recens (NRCS), rapidly replaced by polysaprobic species like Nitzschia palea (NPAL; W4, 8.9%; W6, 24.6%), Nitzschia umbonata (NUMB) (W4, 16.7%; W6, 18.3%), and planktonic species Cyclotella meneghiniana (CMEN) (W4, 7%; W6, 5.4%) and Aulacoseira granulata (AUGR) (W4, 26.2%; W6, 8.4%). In contrast, the initial diatom communities of the Red site transferred to the TL site were present till the experiments ended; but with the apperance of the pollution tolerant species N. palea (NPAL) and N. umbonata (NUMB) being clearly settled with the mean relative abundances around 15.6% and 2%, respectively. Besides these two species (N. palea and N. umbonata), C. meneghiniana (CMEN) appeared as the predominant species at the TL site at the week-2 (before transfer), with the mean relative abundance ≥10% (Fig. 4b). After the transfer from the TL to the Red and from the TL to the NT2 site, the diatom composition of the communities was manifestly modified. For the diatom assemblages transferred to the Red site, the initial dominant species N. palea (NPAL) was still persistent but with lower relative abundances (ranging from 13.5 to 6.2%) until the end of the experiment. N. umbonata (NUMB) and C. meneghiniana (CMEN) were replaced by the dominance of N. recens (NRCS; 30% at W4 and 33% at W6) and Bacillaria paxillifera (BPAX; 5.4% at W4 and 19.5% W6). The assemblages of the biofilms transferred from the TL site to the NT2 were even more modified with initially differing proportions of the four main diatom species: N. palea (NPAL), N. umbonata (NUMB), C. meneghiniana (CMEN), and A. granulata (AUGR).

Relative abundances of main diatom species (mean value, n = 3) with mean relative abundances ≥8% at week 2 (W2) (before transfer), at week 4 (W4), and 6 week (W6) (after transfer) collected at the Red, NT2, and TL stations (NUMB N. umbonata, NPAL N. palea, AUGR A. granulata, GSCA G. scalproides, NCRS N. recens, CMEN C. meneghiniana, BPAX B. paxillifera.) BF before transfer; AF after transfer
Using PCA of 53 diatom species upon a total of 277 modifications in the composition of the transferred diatom communities in comparison with the local diatom communities were graphed in terms of duration time and sampling site (Fig. 5). The graph shows a projection of plane 1/2 explaining 53.9% of total variability. The diatom communities from the Red site were more rapidly discriminated when transferred to the NT2 site than to the TL site. In all other cases, the discrimination from the initial colonization occurred as soon as at week-4 (after 2 weeks of transfer), indicating a clear shift of the communities to adopt the same type of assemblages as those of their transfer sites. The diatom communities transferred to the NT2 (from Red and TL) were characterized by species such as N. umbonata (NUMB), Navicula veneta (NVEN), Sellaphora pupula (SPUP), Navicula cryptocephala (NCRY), A. granulata (AUGR), and Ulnaria ulna (UULN). The groups of diatom assemblages such as G. scalproides (GSCA), N. recens (NRCS), Seminavis strigosa (SMST), Cymbella excisa (CAEX), Achnanthidium minutissimum (ADMI), Sellaphora bacillum (SEBA), and B. paxillifera (BPAX) were representative for the communities of the Red site, transferred communities from the TL to the Red, and transferred communities from the Red to the TL site. The planktonic and polysaprobic species, e.g., Cyclotella atomus (CATO), Amphora montana (AMMO), C. meneghiniana (CMEN), Lemnicola hungarica (LHUN), N. palea (NPAL), Gomphonema parvulum (GPAR), and Eolimna minima (EOMI) characterized TL types (W2) and separated TL communities from the Red and NT types.

Principal component analysis (PCA) based on relative abundances of 53 diatom species (>1% and three replicates per site) at week 2 (W2) (before transfer) and week 4 (W4) and week 6 (W6) (after transfer) of the experiment on glass substrates collected at the Red, NT2, and TL stations (week is indicated in brackets)
Discussion
Qualitative and quantitative characteristics of aquatic organisms presented in communities depend on various physical, chemical, and biological factors (Vilbaste and Truu 2003; Soininen 2003, 2004). These factors determine the whole responses of aquatic organism communities in general and algal communities in particular. In the present study, the alterations of the structural characteristics of the periphytic diatom communities to adapt themselves to a new environment were illustrated by the measurements of the density of diatom cells, diversity index, specie richness, and estimated changes in diatom indices and composition. The structural adaptability of periphytic diatom communities to environmental disturbances was observed after the transfer of early stages of periphytic diatom communities developed on the artificial substrates from the relatively unpolluted site (Red) or the heavily polluted site (TL) to the moderate polluted site (NT2), and the transfer between the Red and the TL site and conversely. In the present study, the responses of the structural periphytic diatom communities determined by multiple sources of pollution in the rivers, the habitat quality, and the directions of transfer were clearly illustrated by the diatom densities. The transferred communities recovered their structure (cell density) at different rates. A significant increase of diatom densities after transfer was observed when the periphytic diatom communities were moved from the polluted site (TL) to the unpolluted site (Red) or even to the moderate polluted site (NT2) whereas a lower difference was noticed in cell density of diatom communities transferred from the reference Red site to the heavily polluted TL site. The slow development of diatom cells transferred from the Red to the TL site could be related to the high level of metals and other contaminants at the TL site (Tetsuro et al. 2009). The growth inhibition of periphytic communities exposed to toxic compound has been suggested by several authors (Tang et al. 1997; Gold et al. 2002; Morin et al. 2010). For the communities coming from the heavily pollution site and transferred to the moderate pollution site, a concomitant increase of biomass occurred after the transfer in parallel with the increase of total density, but this seems to be more related to the high nutrient enrichment conditions in the moderate pollution waters, rather than to the pollution itself. Immigration and emigration played an important role and contributed to increase the number of diatom cells on substrata, in both early and late stage of colonization in our study (Stevenson and Peterson 1991; Stevenson et al. 1991). These observations sustain the results from previous experiments when the dynamics of diatom colonization was followed in similar conditions (Duong et al. 2007). The total density appears then as a useful criterion to rapidly estimate the changes within diatom biofilm in case of returning to unpolluted conditions, but it was not so evident to diagnose the worsening of the water quality as the very dense biofilm matrix constituted in non-polluted conditions needed more than 4 weeks to show clear modifications (Figs. 2 and 3) despite the presence of contaminants, the poor conditions of oxygenation, and high levels of nutrients.
Diversity and species richness are classical indicators of changes in the community structure to disturbances (Jüttner et al. 1996; Sabate 2000), but the interpretation of their positive or negative effects differs between various authors. Sabate (2000) showed a marked decrease in diversity between reference sites in comparison with the affected sites of the Guadiamar River, S-W Spain. Stress effects of mine drainage on the diversity of primary producers in mountain stream were observed by Niyogi et al. (2002), who suggested that physical stress did not as strongly affect the diversity of primary producers in streams as chemical stress. Hill et al. (2000) asserted that periphyton richness was not related to environmental stressors, and Hoagland (1983) reported that periphytic diatom diversity did not reflect adverse environmental conditions as clearly as biomass. In our study, we found that the responses of diversity index and species richness to the water quality changes were not clear (Table 2). No significant differences in species richness and diversity index were found between the local and transferred diatom communities in all cases, except for the communities transferred from the Red to the NT2 site. This result was not surprising as the diatom communities were deeply transformed after the disturbance. The assemblages were rearranged by changing their composition from sensitive species to more tolerant species to cope with the environmentally altered conditions. In this case, numerous replacements of species without a notable and significant decrease of diversity occurred as long as polluted condition could be useful for new assemblages to settle. This adaptation can reversely happen with the replacement of resistant species by more sensitive species in aquatic stress (Kasai 1999), and, in return, communities facing stress may modify their structure and function on their own (Niederlehner and Cairns 1992). Communities react to water quality fluctuation by changing their community composition indicating the occurrence of a recovery process. According to Keller and Yan (1998), three sources of species and individuals can be expected to become forces in the restructuring of aquatic communities. They include the colonization by “external” species from new site, the colonization by “internal” species from the resident site, and the expansion of species that have persisted in the reduced abundance in the stressed community. Thus, the diatom communities transferred from the Red to the NT2 site, from the TL to the Red site, and from the TL to the NT2 site changed quickly their compositions to adapt to new conditions (Figs. 4a, b and 5). The communities characterized by rich nutrients, high conductivity, and low dissolved oxygen with the dominant species such as N. umbonata (NUMB), C. meneghiniana (CMEN), and N. palea (NPAL) were replaced by less pollution tolerant species such as N. recens (NRCS), B. paxillifera (BPAX), and sensitive species A. minutissimum (ADMI) when diatom assemblages were moved from the TL site to the Red site. On the other hand, the complete replacement of pollution-tolerant species in the transferred communities to the NT2 site (Red to NT2 and TL to NT2) within 2 weeks after the translocation showed the capacity of rapid recovery of periphytic diatoms after environmental changes. Our results were in agreement with those of Gold et al. (2002), who observed the structural community adaptation to the new environmental conditions within 2 weeks after the exposure to metal pollution in the French hydrosystem. However, data on the time needed for periphytic assemblages to reveal a change in environmental conditions are variable. Previous studies demonstrated that periphytic algal assemblages react rapidly to the degradation of the water quality, but less rapidly to the improvement of the water quality (Iserentant and Blancke 1986; Ivorra et al. 1999; Tolcach and Gomez 2002; Lacoursière et al. 2011). In the work of Tolcach and Gomez (2002), the microbenthic assemblages were still different after 2 weeks of being transferred from polluted to unpolluted sites. Iserentant and Blancke (1986) reported that no significant changes were observed in the dominant diatom species in transferred community after 45 days. Morin et al. (2010) also showed that the characteristics of the translocated communities were not as acquired as the characteristics of the reference communities within 1 month and a longer translocation time would be expected. Lavoie et al. (2008) underlined that response time of diatom assemblages to simulate water quality improvement, and degradation varied between 1 to 5 weeks as a function of trophic status. Their study showed that the change in diatom assemblage structure and IDEC were observed in the oligotrophic, mesotrophic, and eutrophic rivers within 1, 2, and 5 weeks, respectively. Another study showed that diatom communities responded to water quality improvement only after 60 days (Rimet et al. 2005). It has been shown that the colonization of new substrata and the establishment of mature biofilm can take 2 to 4 weeks (Hoagland et al. 1982; Tien et al. 2009). Thus, in the present study, the colonization of substratum took place over a period of 2 weeks (before the transfer) and could establish a mature diatom assemblage. The biofilm matrix densely covering substrata and its adhesion to the substratum seemed to limit the settlement of indigenous diatom species of the polluted site (TL) to the transferred samples. Unlike what was observed for the transferred communities from the Red to the NT2, from the TL to the Red, and from the TL to the Red sites, the transferred samples from the Red site to the TL site did modify their structure but not completely (Fig. 5). At the end of the experiment within the transferred communities of the TL site, the initial species of the Red site such as N. recens (NRCS), B. paxillifera (BPAX), and G. scalproides (GSCA) still represented in minor proportion and some native diatom species of the TL site such as N. palea (NPAL) and C. meneghiniana (CMEN) were already settled and gradually increased at W6 (after 4 weeks transfer). The diatom communities transferred to the TL site showed a slower response to short-term disturbance. After 4 weeks of translocation, the composition of the transferred diatom assemblages was still different to the local diatom communities. This slower recovery may be explained by the architecture of the diatom assemblages within the biofilm. The complexity of the assemblages, the life form of the dominant diatom species, and the sensitivity or resistance of diatom species have been considered as the important factors influencing the time for assemblage restructuring (Rimet et al. 2009; Lacoursière et al. 2011). It has been shown that nutrient enrichments were not the only parameters affecting diatom assemblage communities. The increased metal concentration and other contaminants presented at the TL site played a possible and important role in the responses of community structure (Soldo and Behra 2000). The nature and source of wastewater effluents were diversified but with no precise data. In this case, a longer translocation time would be necessary for the transferred diatom communities to the TL site to acclimate to the TL water quality conditions.
In comparison with the results obtained in 2003, the diatom assemblages in the present study were similar. However, the proportion of dominant diatom species between the 2 years varied. In particular, the phytoplanktonic Aulacoseira granulate (AUGR) was less abundant in the translocation experiment. As shown in another study, this species is mainly observed in large and turbid as well as in moderately eutrophic waters (O’Farrell et al. 2001). Most of the dominant diatom species in this study have already been described as pollution-moderate/tolerant species (Lange-Bertalot 1979). The results can be explained by the low water level of the rivers during the dry season of 2005 (Duong et al. 2007). The diatom indices (IPS and DAIPo) have already been applied successfully to assess river water quality in Vietnam (Duong et al. 2006; 2007). The shift in diatom composition of the transferred assemblages resulted in change of IPS and DAIPo values (Table 2). In this study, both of them gave a similar trend of improvement in water quality when the diatom communities were transferred from the polluted site to the unpolluted and moderate polluted sites, and a trend of water quality worsening when the communities were transferred from the unpolluted site to the heavily polluted and the moderate polluted sites (Table 2). The recovery of diatom indices was clearly noticed as soon as at the 2 weeks after the transfer to the reference site. A slow decrease in diatom indices occurred when the diatom communities were moved from the Red to the TL site, even if a proportion of diatoms species characterizing the Red site still remained in its biofilm. These results are in agreement with the previous study (Lavoie et al. 2008) where the integration period of diatom indices was defined from 2 to 5 weeks as a function of trophic status. However, Rimet et al. (2005) observed that the time needed for an assemblage to adapt themselves to environmental changes varied between 29 and 60 days and depended on the diatom index considered. In the present study, the changes of diatom indices showed a sensitivity of indices to the alteration of water quality. This result is in agreement with the observation of Rimet et al. (2005) in which the diatom index sensitivity to water quality was changed by using the transferred biofilms from several polluted rivers to an unpolluted stream. According to them, several indices such as IPS (Index of pollution sensitivity), ROT (Saprobic index of Rott), SHE (Schiefele and Schreiner index), EPI (Eutrophication Pollution Index), and CEE (European index) well reflected the improvement in water quality. DAIPo should be added to this list of valuable indices. Thus, diatom indices appear as relevant global criteria to assess the water quality in rivers.
Conclusion
The responses of periphytic diatom communities to the water quality changes were investigated by transferring the early diatom communities colonized on glass substrata from a non-polluted river to less polluted or polluted rivers, and conversely. The responses and adaptability of the periphytic diatom communities to the environmental changes varied and not only depended on the specific environmental conditions, but also on the architecture of the assemblages. The transfer experiments have led to a replacement of species to adapt the periphytic biofilm to the new environmental conditions after 2 to 4 weeks exposure. These changes in taxonomic composition were recoded by the shifts in IPS and DAIPo indices, which proved to be sensitive to the water quality changes. The recovery or replacement of the diatom community biofilm highlights their potential to integrate environmental variation over long periods and their possible role to check both the restoration and pollution status of aquatic systems.
References
APHA (1995) American Public Health Association. American Water Works Association (AWWA), Water Environment Federation (WEF). Standard methods for the examination of water and wastewater, 19th edn. American Public Health Association, Washington
Barranguet C, Charantoni E, Plans M, Admiraal W (2000) Short-term response of monospecific and natural algal biolms to copper exposure. Eur J Phycol 35:39–406
Cattaneo A, Couillard Y, Wunsam S, Courcelles M (2004) Diatom taxonomic and morphological changes as indicators of metal pollution and recovery in Lac Dufault (Québec, Canada). J Paleolimnol 32:163–175
Cemagref (1982) Etude des méthodes biologiques d’appréciation quantitative de la qualité des eaux. Rapport Q. E. Lyon-A. F. Bassin Rhône-Méditeranée-Corse-Cemagref, Lyon, France, p 218
Centis B, Tolotti M, Salmaso N (2010) Structure of the diatom community of the River Adige (North-Eastern Italy) along a hydrological gradient. Hydrobiologia 639:37–42
Descy JP, Coste M (1991) A test of methods for assessing water quality based on diatoms. Verh Internat Limnol 24:2112–2116
Dixit SS, Smol JP, Kinston JC, Charles DF (1992) Diatoms: powerful indicators of environmental change. Environ Sci Tech 26:23–33
Duong TT, Coste M, Feurtet-Mazel A, Dang DK, Gold C, Park YS, Boudou A (2006) Impact of urban pollution from the Hanoi area on benthic diatom communities collected from the Red, Nhue and To Lich rivers (Vietnam). Hydrobiologia 563:201–216
Duong TT, Feurtet-Mazel A, Coste M, Dang DK, Boudou A (2007) Dynamics of diatom colonization process in some rivers influenced by urban pollution (Hanoi, Vietnam). Ecol Indic 7:839–851
EN 13946 2003 (2003) Water quality. Guidance standard for the routine sampling and pre-treatment of benthic diatom from rivers, p. 18
EN 14407 2004 (2004) Water quality. Guidance standard for the identification, enumeration and interpretation of benthic diatom samples from running waters, p. 16
Genter RB (1996) Ecotoxicology of inorganic chemical stress to algae. In: Stevenson RJ, Bothwell ML, Lowe RL (eds) Algal ecology of freshwater benthic ecosystem, aquatic ecology series. Academic, Boston, pp 403–468
Gibson CE, Fitzsimons AG (1991) Light break in the dark period depresses the growth rate of a freshwater planktonic diatom. Diatom Res 6:15–20
Gold C, Feurtet-Mazel A, Coste M, Boudou A (2002) Field transfer of periphytic diatom communities to assess short-term structural effects of metals (Cd, Zn) in rivers. Water Res 36:3654–3664
Gold C, Feurtet-Mazel A, Coste M, Boudou A (2003) Impacts of metals (Cd, Zn) on the development of periphytic diatom communities within outdoor artificial streams along a pollution gradient. Arch Environ Con Tox 44:189–197
Hill BH, Willingham WT, Parrish LP, McFarland BH (2000) Periphyton community responses to elevated metal concentrations in a Rocky Mountain stream. Hydrobiologia 428:161–169
Hoagland KD (1983) Short-term standing crop and diversity of periphytic diatom in a eutrophic reservoir. J Phycol 19:30–38
Hoagland KD, Roemer SC, Rosowski JR (1982) Colonization and community structure of two periphyton assemblages, with emphasis on the diatoms (Bacillariophyceae). Am J Bot 69:188–213
Iserentant R, Blancke D (1986) A transplantation experiment in running water to measure the response rate of diatoms to changes in water quality. In: Ricard M (ed), Proceedings of the 8th International Diatom Symposium, Paris, 1984. Koeltz Scientific Books, Koenigstein. In Proceedings of the 8th Diatom Symposium, Paris, pp. 347–354
Ivorra N, Hettelaar J, Tubbing GMJ, Kraak MHS, Sabater S, Admiraal W (1999) Translocation of microbenthic algal assemblages used for in situ analysis of metal pollution in rivers. Arch Environ Contam Toxicol 37:19–28
Jüttner I, Rothfritz H, Ormerod JO (1996) Diatoms as indicators of river quality in the Nepalese Middle Hills with consideration of the effects of habitat-specific sampling. Freshwater Biol 36:475–486
Kasai F (1999) Shift in herbicide tolerance in paddy field periphyton following herbicide application. Chemosphere 38:919–931
Keller W, Yan ND (1998) Biological recovery from lake acidification: zooplankton communities as a model of patterns and processes. Restor Ecol 6:364–375
Kono Y, Doan TD (1995) Effect of water control on rice cultivation in the Red river delta, Vietnam: a case study in the Nhue river irrigation system. JSEAS 32:425–445
Krammer K, Lange-Betarlot H (1986–1991) Bacillariophyceae. 1.Teil: Naviculaceae. 876 p; 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae, 596 p; 3. Teil: Centrales, Fragilariaceae, Eunotiaceae, 576 p; 4. Teil: Achnanthaceae. Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema. 437 p. In: H, Ettl Gerloff, J Heynig, H Mollenhauer, D. (Eds.), Süßwasserflora von Mitteleuropa. Gustav Fischer Verlag, Stuttgart, 2485 pp.
Kurosawa K, Do NNH, Nguyen HT, Ho TLT, Nguyen TC, Kazuhiko E (2004) Monitoring of inorganic nitrogen levels in the surface and ground water of the Red River Delta, Northern Vietnam. Commun Soil Sci Plant 35:1645–1662
Lacoursière S, Lavoie I, Rodríguez MA, Campeau S (2011) Modeling the response time of diatom assemblages to simulated water quality improvement and degradation in running waters. Can J Fish Aquat Sci 68:487–497
Lange-Bertalot H (1979) Pollution tolerance of diatoms as a criterion for water quality estimation. Nova. Hedwigia 64: 285–304
Lavoie I, Campeau S, Grenier M, Dillon PJ (2006) A diatom-based index for the biological assessment of eastern Canadian rivers: an application of correspondence analysis (CA). Can J Fish Aquat Sci 63:1793–1811
Lavoie I, Campeau S, Darchambeau F, Cabana G, Dillon PJ (2008) Are diatoms good integrators of temporal variability in stream water quality? Freshwater Biol 53:827–841
Le TPQ (2005) Biogeochemical functioning of the Red River (North Vietnam): budgets and modeling. Thèse Université Paris VI (France) and Vietnam Academy of Science and Technology (VAST), p. 187
Lecointe C, Coste M, Prygiel J (1993) “OMNIDIA” software for taxonomy, calculation of diatom indices and inventories management. Hydrobiologia 269/270:509–513
Lowe RL, Pan Y (1996) Benthic algal communities as biological monitors. In: Stevenson RJ, Bothwell ML, Lowe RL (eds) Algal ecology of freshwater benthic ecosystem, aquatic ecology series. Academic, Boston, pp 705–739
McCormick PV, Stevenson RJ (1998) Periphyton as a tool for ecological assessment and management in the Florida everglades. J Phycol 34:726–733
McCune B, Mefford MJ (1999) PC-ORD. Multivariate analysis of ecological data. Version 4.0. MjM Software Design, Gleneden Beach, Oregon USA, p. 237
Morin S, Vivas-Nogues M, Duong TT, Boudou A, Coste M, Delmas F (2007) Dynamics of benthic diatom colonization in a cadmium/zinc-polluted river (Riou-Mort, France). Fundam Appl Limnol 168:179–187
Morin S, Bottin M, Mazzella N, Macary F, Delmas F, Winterton P, Coste M (2009) Linking diatom community structure to pesticide input as evaluated through a spatial contamination potential (Phytopixal): a case study in the Neste river system (South-West France). Aquat Toxicol 94:28–39
Morin S, Pesce S, Tlili A, Coste M, Montuelle B (2010) Recovery potential of periphytic communities in a river impacted by a vineyard watershed. Ecol Indic 10:419–426
Nguyen QT (2001) Impact of wastewater on water quality in irrigation system and treatment measures to reduce pollution: a case study in Nhue irrigation system. In: Working Paper 30 Wastewater reuse in agriculture in Vietnam: water management, environment and human health aspects. Proceedings of a workshop held in Hanoi (Vietnam), IWMI, p 18–19.
Niederlehner BR, Cairns J (1992) Community response to cumulative toxic impact: effects of acclimation on zinc tolerance of Auwuchs. Can J Fish Aquat Sci 49:2155–2163
Niyogi DK, Lewis WMJ, McKnight DM (2002) Effects of stress from mine drainage on diversity, biomass, and function of primary producers in mountain streams. Ecosystem 5:554–567
O’Farrell I, Tell G, Podlejski A (2001) Morphological variability of Aulacoseira granulata (Ehr.) Simonsen (Bacillariophyceae) in the Lower Paraná River (Argentina). Limnology 2:65–71
Pan YD, Stevenson RJ, Vaithiyanathan P, Slate J, Richardson CJ (2000) Changes in algal assemblage along observed and experimental phosphorus gradients in a subtropical wetland, USA. Freshwater Biol 44:339–353
Potapova M, Charles FD (2005) Choice of substrate in algae-based water-quality assessment. J N Am Bentholl Soc 24:415–427
Rimet F, Cauchie HM, Hoffmann L, Ector L (2005) Response of diatom to simulated water quality improvements in a river. J Appl Phycol 17:119–128
Rimet F, Ector L, Cauchie HM, Hoffmann L (2009) Changes in diatom-dominated biofilms during simulated improvements in water quality: implications for diatom-based monitoring in rivers. Eur J Phycol 44:567–577
Rott E, Duthie HC, Pipp E (1998) Monitoring organic pollution and eutrophication in the Grand River, Ontario, by means of diatoms. Can J Fish Aquat Sci 55:1443–1453
Sabate S (2000) Diatom communities as indicators of environmental stress in the Guadiamar River, S-W Spain, following a major mine tailing spill. J Appl Phycol 12:113–124
Shannon CE, Weaver W (1963) The mathematical theory of communication. University of Illinois, Urbana, p 125
Soininen J (2003) Heterogeneity of benthic diatom communities in different spatial scales and current velocities in a turbid river. Archiv für Hydrobiologie 156:551–564
Soininen J (2004) Determinants of benthic diatom community structure in boreal streams: the role of environmental and spatial factors at different scales. Int Rev Hydrobiol 89:139–150
Soldo D, Behra R (2000) Long-term effects of copper on the structure of freshwater periphyton communities and their tolerance to copper, zinc, nickel and silver. Aquat Toxicol 47:181–189
StatSoft Inc (2004) STATISTICA (data analysis software system), version 7. www.statsoft.com
Stevenson RJ, Peterson CG (1991) Emigration and immigration can be important determinants of benthic diatom assemblages in streams. Freshwater Biol 26:279–294
Stevenson RJ, Peterson CG, Kirschtel DB, King CC, Tuchman NC (1991) Density-dependent growth, ecological strategies and effects of nutrients and shading on benthic diatom succession in streams. J Phycol 27:59–69
Tang JX, Hoagland KD, Siegfried BD (1997) Differential toxicity of atrazine to selected freshwater algae. Bull Contam Toxicol 59:631–637
Tetsuro K, Takuma F, Huynh TH, Shuzo T (2009) Assessment of heavy metal pollution in river of Hanoi, Vietnam using multivariate analysis. Environ Contam Tox 83:575–582
Tien Ch J, Wub WH, Chuang TL, Chen CS (2009) Development of river biofilms on artificial substrates and their potential for biomonitoring water quality. Chemosphere 76:1288–1295
Tolcach ER, Gomez N (2002) The effect of translocation of microbenthic communities in a polluted lowland stream. Verh Int Verein Limnol 28:254–258
Trinh AD (2003) Etude de la qualité des eaux d’un hydrosystème fluvial urbain autour de Hanoi (Vietnam) suivi expérimental et modélisation. Thèse Université Grenoble 1, France and Vietnam Academy of Science and Technology (VAST), 265 pp.
Vilbaste S, Truu J (2003) Distribution of benthic diatoms in relation to environmental variables in lowland streams. Hydrobiologia 493:81–93
Watanabe T, Asai K, Houki A (1986) Numerical estimation to organic pollution of flowing water by using epilithic diatom assemblage. Diatom assemblage Index (DAIpo). Sci Total Environ 55:209–218
Whitton BA, Rott E (1996) Use of Algae for monitoring rivers II. Institut für Botanik, Universität Innsbruck, Innsbruck, Austria, p. 196
Acknowledgments
This study was carried out at first in the scope of the PhD thesis of T.T. Duong, financially supported by CNRS (Centre National de la Recherche Scientifique) and the ESPOIR programme, a French-Vietnamese co-operation project. Thanks to Prof. Georges Vachaud, Research Director at the CNRS for the coordination of the ESPOIR program (CNRS–VAST). This work was then performed in the framework of the NAFOSTED (Nasfosted Vietnam’s National Foundation for Science and Technology Development) (105.09.89.09) project. We thank Mr. Joost Hegger for improvements to the native style of the manuscript. The authors thank many individuals for their help in collecting samples in the field. We gratefully acknowledge the anonymous reviewers whose comments and suggestions significantly improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Duong, T.T., Coste, M., Feurtet-Mazel, A. et al. Responses and structural recovery of periphytic diatom communities after short-term disturbance in some rivers (Hanoi, Vietnam). J Appl Phycol 24, 1053–1065 (2012). https://doi.org/10.1007/s10811-011-9733-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-011-9733-9