
Abstract
Twenty-two tropical seaweeds from the Rhodophyta, Phaeophyta and Chlorophyta were examined for their possible use as nutritional supplements. All seaweeds contained balanced Na/K and C/N ratio and high amounts of macroelements (Na, K, Ca, and Mg) as compared to the terrestrial vegetables. Among the microelements, Fe was the highest followed by Zn, Mn, Cu and other trace elements. Fatty acid distribution showed high level of n-6 and n-3 polyunsaturated fatty acids (PUFAs), and their ratios were within the WHO prescribed limits. The higher ratios of PUFA/SFA (>0.4) are in agreement with the recommendations of nutritional guidelines. Most of the species, especially the Chlorophyta and Phaeophyta, had permissible intake values of unsaturation, atherogenic and thrombogenic indexes comparable to milk-based products. Principal component analysis demonstrated a correlation between total phenolic content, total antioxidant activity, DPPH, and O •−2 radical scavenging activity, suggesting polyphenols as the chief contributor to the antioxidant activity in seaweeds. These results indicate that these seaweeds could be a potential source of natural antioxidants, minerals and high-quality PUFAs and may be efficiently used as ingredients in functional foods.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Increasing awareness among consumers about health-promoting foods has aroused interest in food supplement research worldwide. In addition to food supplements, consumption of exotic foods with proven nutritional values has also been gaining prominence in several developed countries (Herrero et al. 2006). Many of these foods are presently promoted and marketed as functional foods with premium price. The beneficial actions of these foods are reported to be mainly due to their functional components such as minerals, antioxidants and n-3 fatty acids, which are either absent in the analogous traditional foods or present only in trace concentrations. Consequently, there has been a quest to explore and utilize foods from nonconventional sources of both terrestrial and marine origin to enhance the nutritional quality of human foods which in turn also reduces the dependability on traditional foods.
Seaweeds with their diverse bioactive compounds (Lee et al. 2008; Zubia et al. 2009) have opened up potential opportunities in pharmaceutical and agri-food processing industries. The consumption of seaweeds as part of diet has been shown to be one of the prime reasons for low incidence of breast and prostate cancer in Japan and China compared to North America and Europe (Pisani et al. 2002). Seaweeds also contain sufficient amounts of protein, polysaccharides (e.g., alginates, fucans and laminarans), and amino acids of considerable nutritional importance. Algal lipids (1–3% dry matter) contain a high proportion of essential fatty acids particularly n-3 polyunsaturated fatty acids (PUFAs). At present, marine fish and their oil are the major commercial sources of PUFAs, but their suitability for human consumption has been questioned from the biosafety perspective.
The mineral content of seaweeds is usually high enough (8–40%) to fulfill the recommended daily intakes of essential macroelements and trace elements for human nutrition (Rupérez 2002). Polyphenols in seaweeds are the foremost natural antioxidants and considered to be superior to that of essential vitamins. Moreover, these natural antioxidants have easy and unlimited access to metabolic processes in the body and produce virtually none of the side effects associated with synthetic antioxidants (Farag et al. 2003).
In India, seaweeds are exclusively utilized for the production of industrially important phycocolloids such as agar-agar, alginate, and carrageenan and have never been consumed in any form in the diet. In recent years, seaweeds from different parts of the world have been extensively studied for their antioxidant properties and mineral contents as part of nutraceutical applications. But there are no such reports on the antioxidants activities together with mineral composition and n-3 and n-6 PUFAs highlighting the unsaturation (UI), atherogenic (AI) and thrombogenic indexes (TI) of tropical seaweeds from the Indian region. Therefore, the present study was undertaken to evaluate the nutritional potentials in terms of minerals, antioxidant properties and PUFAs of the most abundantly available seaweeds from Saurashtra coast of India.
Materials and methods
A total of 22 different macroalgal species, belonging to Chlorophyta, Rhodophyta and Phaeophyta, were collected during January to April, 2009, from Saurashtra coast (20° 54′ N, 70° 22′ E), Gujarat, India. The thalli of all the algae were harvested manually during low tide and then transported to the laboratory in wet tissue towels in an ice box. They were then thoroughly cleaned with brush in autoclaved seawater to remove attached epiphytes and detritus and were then divided into three parts. One part was lyophilized and stored in airtight containers at −40°C for the estimation of antioxidant potential while the second part was dried at 60°C in an oven to constant weight for mineral analysis. The last part was shade-dried and stored at −40°C for fatty acid analysis.
The proximate composition including protein, carbohydrate and lipid content was determined following the methods of Lowry et al. (1951), Dubois et al. (1956), and Bligh and Dyer (1959). The mineral content of the samples was determined by the method of Santoso et al. (2006). Dried samples (200 mg) were weighed in a Kjeldahl flask, and 20 mL of concentrated HNO3 was added to the sample and left to stand overnight. Concentrated HClO4 (5 mL) and H2SO4 (0.5 mL) were added to the flask and then heated until no white smoke was emitted. These digested samples were dissolved in 100 mL of 2% HCl and filtered with a 0.22-μm membrane filter. Mineralogical analysis was carried out using inductively coupled plasma atomic emission spectroscopy (Perkin-Elmer, Optima 2000, USA) using inductively coupled plasma multielement reference standard solution VIII (product no. 1.09492.0100, Merck, Germany) with a concentration of 10 and 4 mg L−1 for macroelements and microelements analyzed. The average recoveries (%) for the reference standards were Na (98.93), K (97.59), Ca (99.78), Mg (104.12), Fe (97.26), Cu (98.84), Zn (99.82), Mn (102.17), Ni (98.95), Mo (101.16), As (99.89), P (91.27), and Se (97.55). The total carbon and nitrogen content was determined by combusting the dried samples using a CHN Elemental Analyzer (Perkin-Elmer Model 2400, USA), calibrated using acetanilide as a reference standard.
For estimation of antioxidant potential, lyophilized samples (10 g) were extracted twice with 50 mL of methanol at room temperature for 72 h in the dark. Extracts were combined, filtered, and evaporated to dryness under vacuum at 40°C using a rotary evaporator to give a dark green viscous mass (methanolic extract). The dried extract was dissolved in 0.2% methanol and stored at −40°C until use.
Total content of phenolic compounds was determined spectrophotometrically using Folin-Ciocalteu reagent following Lim et al. (2002). In brief, extracts at a concentration of 1 mg mL−1 were first diluted with methanol, and then an aliquot of 0.1 mL from the diluted extracts was added into the test tubes. To this, 2.9 mL of distilled water and 0.5 mL of Folin-Ciocalteu’s reagent were added and mixed thoroughly. After 10 min, 1.5 mL of 20% sodium carbonate solution was added, and the mixture was mixed thoroughly and allowed to stand at room temperature in the dark for 1 h. Absorbance was measured at 725 nm, and total content of phenolic compounds was calculated based on a standard curve of phloroglucinol.
2,2-Diphenyl-1-picrylhydrasyl (DPPH) radical scavenging activity was determined according to Kordali et al. (2005). Briefly, extract at a concentration of 1 mg mL−1 was mixed with 3.0 mL DPPH solution (25 mg L−1) prepared daily. Due to color intensity of the extracts, it was necessary to prepare a blank of 1 mL of each extract added to 3.0 mL of methanol. The reaction was completed after 2 h in the dark at room temperature, and the absorbance was read at 515 nm. The scavenging effect (%) was calculated by using the formula given by Duan et al. (2006), using butylated hydroxyanisole (BHA) and butylated hydroxytoluene (BHT) as standard at a concentration 500 μg mL−1.
Reducing power of the extracts was evaluated according to Yen and Chen (1995). Extract samples at different concentrations (0.25, 0.50 and 1 mg mL−1) were mixed with phosphate buffer (1.25 mL, 0.2 M, pH 6.6) and 1.25 mL of potassium ferricyanide (1%) and incubated at 50°C for 20 min, cooled and mixed with 1.25 mL of trichloroacetic acid (10%); 1.25 mL of this mixture was transferred to other test tubes to which distilled water (1.25 mL) and FeCl3·6H2O (0.25 mL, 0.1%) were added. The mixture was centrifuged and kept at room temperature for 10 min before reading the absorbance at 700 nm. Increased absorbance of the reaction mixture indicated increasing reducing power.
Total antioxidant activity (TAC) was determined according to Prieto et al. (1999). In brief, the methanolic extract at a concentration of 1 mg mL−1 was mixed with 3.0 mL reagent solution containing 0.6 M H2SO4, 28 mM sodium phosphate, and 4 mM ammonium molybdate and incubated at 95°C for 90 min in water bath. The absorbance was measured at 695 nm, using ascorbic acid as standard.
The superoxide anion (O •−2 ) scavenging activity of the extracts was determined using the nonenzymatic system outlined by Lim et al. (2002) while preparing all the reagents with Tris–HCl buffer (16 mM, pH 8.0). In the test tubes, 1 mL of 234 μM NADH, 1 mL of 150 μM NBT, and 0.2 mL of the seaweed methanolic extract at concentrations (1 mg mL−1) were mixed together with 0.8 mL of 37.5 μM phenazine methosulfate (PMS), and after 5 min the absorbance was measured at 560 nm. The same mixture without sample extract was used as control, and blanks for each extract were prepared without PMS. BHA and BHT were used as standards at a concentration 500 μg mL−1.
Fatty acids obtained from lipids were converted to respective methyl esters by trans-methylation using 1% NaOH in methanol and heated for 15 min at 55°C, followed by addition of 5% methanolic HCl and heating for 15 min at 55°C. Fatty acid methyl esters (FAMEs) were extracted in hexane, and the organic phase was evaporated to dryness under reduced pressure. FAMEs were analyzed by GC-2010 coupled with GCMS-QP2010 (Shimadzu, Japan). Nonadecanoic acid was used as internal standard. FAME peaks were identified by comparison with their respective retention times obtained from standard mixture (FAME Mix C4-C24, Sigma) by GC–MS postrun analysis and quantified by area normalization. For the validation of this method, a standard FAME mixture was analyzed on intra-/inter-day basis to ensure the instrument response and data accuracy with a recovery rate of nearly 90–98% for standard fatty acids. A five-point calibration was also carried out to ensure the linearity.
The AI and TI were calculated according to De Lorenzo et al. (2001), where:
All data are expressed as mean ± standard deviation (n = 3), except for minerals where a total of five replicates were analyzed. Data analysis was carried out by one-way analysis of variance, and a significant difference was considered at the level of p < 0.01. Principal component analysis (PCA) was accomplished using Unscrambler 9.8 software package (CAMO AS, Trondheim, Norway).
Results
Proximate composition and minerals
Table 1 shows the proximate composition including carbohydrate, protein and lipid content. The carbohydrate content was significantly higher (p < 0.01) in the Rhodophyta and ranged from 31.43% to 61.63% on a dry weight basis (DW) followed by Chlorophyta (33.10–57.96% DW) and Phaeophyta (21.99–33.30% DW). The protein content varied significantly between species and ranged from 6.90% to 22.11% DW. The total lipid content in all species was low (<4% DW), with the highest lipid content of 3.03 ± 0.21% DW in Caulerpa scalpeliformis. A wide variation in mineral content was observed among species and phyla (Table 2). Among the macroelements, all the rhodophycean species were rich in K and ranged from 6.04% to 21.95% DW with a maximum in Kappaphycus alvarezii. The Na content in most of the red algae was almost half or even less than the K content and ranged from 2.89% to 8.04% DW. Similarly, phaeophycean species also had a relatively higher K content (2.49–9.07% DW) compared to Na (1.19–6.24% DW). In contrast, in the green algae, the Na content was either higher or almost equal to the K content and ranged from 4.05% to 7.23% DW. It is also worth noting that the Na/K ratio varied from species to species and ranged from 0.30 to 2.24 in the studied algae groups.
A summary of trace elements (in mg 100 g−1 DW) is given in Table 2. Among the microelements present in seaweeds, Fe was the most abundant trace element and was highest in both brown and green algae and ranged from 19.88 to 44.72 and 17.44 to 44.54 mg 100 g−1, respectively, followed by red algal species with 6.80–39.62 mg 100 g−1. The Zn content was lowest in C. scalpeliformis with 1.31 ± 0.14 mg 100 g−1 while Padina tetrastromatica had the highest with 17.09 ± 1.81 mg 100 g−1. The Mn content was considerably higher in Ulva species (U. fasciata with 8.31 ± 0.16 mg 100 g−1, U. reticulata with 9.32 ± 0.23 mg 100 g−1 and U. rigida with 6.30 ± 0.19 mg 100 g−1) than its average content in the Phaeophyta with 4.75 mg/100 g mg 100 g−1 and Rhodophyta with 3.21 mg 100 g−1. The lowest and highest Cu concentrations were 0.29 ± 0.02 mg 100 g−1 in Gracilaria debilis and 1.28 ± 0.06 mg 100 g−1 in U. reticulata. The content of other trace elements such as Ni, Se, Mo and As (mg 100 g−1 DW) among the algal groups were in the range of 0.11–0.77 for Ni, 0.09–0.46 for Se, 0.05–0.48 for Mo, and 0.08–0.63 for As (Table 2).
The average C/N ratio did not differ significantly among the algal groups and was in the order of Phaeophyta (16.79) > Rhodophyta (14.48) > Chlorophyta (13.80) (Table 3). Cystoseira indica and Sargassum species (viz. Sargassum swartzii and Sargassum tenerrimum) were rich in P with values 0.50 ± 0.02%, 0.43 ± 0.04% and 0.38 ± 0.04% DW, respectively. The highest and lowest P contents were 0.59 ± 0.03% DW in Gelidiella acerosa and 0.13 ± 0.01% DW in U. fasciata, respectively (Table 3). The sulfate content of two important agarophytes G. acerosa (1.94 ± 0.15% DW) and G. dura (2.12 ± 0.28) was low but was quite high in Sargassum species (S. swartzii with 7.34 ± 1.31% DW and S. tenerrimum with 7.69 ± 0.72% DW). The average sulfate content among the phaeophycean (6.80% DW) and chlorophycean species (4.99% DW) was significantly higher when compared to rhodophycean algae (3.63% DW).
Antioxidant properties
Table 3 shows the extraction yields of the methanolic extracts on dry weight basis. The yield was maximal for the Phaeophyta and Chlorophyta ranging from 19.45% to 33.88% and 19.85–27.52%, respectively.
Total phenolics, DPPH and superoxide radical scavenging activity
Significant differences for total phenolic content (TPC) among different species were observed (Table 3). In general, brown seaweeds contained higher amounts of polyphenols ranging from 43.75 to 55.23 mg PGE g−1 extract when compared to green (32.57–56.22 mg PGE g−1 extract) and red (22.79–65.60 mg PGE g−1 extract) seaweeds. In particular, G. acerosa, Sarconema filiforme, C. indica, S. tenerrimum and C. racemosa were exceptionally rich in polyphenols. The present investigation has shown considerably high DPPH scavenging activity (%) in order of Chlorophyta ≥ Phaeophyta > Rhodophyta (Table 3), with the most effective being C. racemosa, S. tenerrimum, C. indica and C. veravalnensis followed by G. acerosa and Ulva rigida. The DPPH scavenging activity of commercial antioxidants like BHA and BHT was 89% and 96%, respectively, when used at a concentration of 500 μg mL−1. Superoxide radical scavenging activity (%) was consistently higher in the Phaeophyta and Chlorophyta (Table 3). The most effective among them were S. swartzii, C. indica, C. veravalnensis and C. scalpeliformis. Scavenging activity for commercial antioxidants like BHA and BHT were 93% and 88%, respectively, when used in concentration of 500 μg mL−1. Among the rhodophytes, S. filiforme, G. acerosa and Laurencia cruciata exhibited fairly high superoxide radical scavenging activity, while in other red algal species such as Amphiora anceps, K. alvarezii, Gelidium micropterum and G. fergusonii, it was much lower.
Total antioxidant activity and reducing power
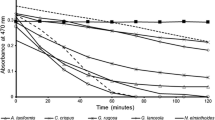
The total antioxidant activities (Table 3) were higher in the Phaeophyta with 0.91 ± 0.19 mg AAE g−1extract (S. tenerrimum) and 0.79 ± 0.22 mg AAE g−1 extract (C. indica), while among the Chlorophyta U. rigida exhibited the maximum total antioxidant activity of 0.70 ± 0.08 mg AAE g−1 extract. Of the Rhodophyta. G. acerosa and S. filiforme had the maximum antioxidant activity. In the present study, the reducing power increased with increasing concentration in all samples and exhibited the OD value of <1.0 at the studied concentrations. Hence, data have been presented for the maximum concentration (1,000 μg mL−1) of methanolic extract for each species (Fig. 1). Methanolic extract of S. swartzii, S. tenerrimum, S. asperum, C. indica, C. racemosa and C. veravalnensis had the highest reducing power with absorbance >1.0. Nevertheless, among rhodophytes, S. filiforme and G. acerosa were the most effective with an absorbance of 1.48 ± 0.06 and 0.77 ± 0.04, respectively, while the rest demonstrated an average absorbance of 0.50.

Reducing power of methanolic extract of 22 macroalgal species indexed from 1 to 22 according to Table 1
FA composition
In the analyzed seaweed varieties, palmitic acid (C16:0) was the most abundant SFA while myristic acid (C14:0), pentadecanoic acid (C15:0), and heptadecanoic acid (C17:0) were found in trace amounts. The sum of SFA ranged from 31.03% of total fatty acid (TF) in G. corticata to 78.16% TF in G. salicornia among Rhodophyta, 36.03% TF (S. asperum) to 46.63% TF (P. tetrastromatica) in the Phaeophyta, and 48.56% TF (C. scalpeliformis) to 63.67% TF (U. reticulata) in the Chlorophyta (Table 4). The average content for the sum of SFA followed the order Chlorophyta > Rhodophyta ≥ Phaeophyta. Among the investigated species, the red alga K. alvarezii and the green algae U. rigida and U. reticulata had the highest proportion of monounsaturated fatty acid (MUFA) (Table 4), and oleic acid was the predominant fatty acid (FA).
Most of the seaweed samples analyzed had a high proportion of PUFA (Table 4). The sum of PUFA ranged from 11.19% TF (G. salicornia) to 65.46% TF (G. corticata) among Rhodophyta, from 39.39% TF (P. tetrastromatica) to 49.57% TF (S. asperum) in Phaeophyta, and from 14.74% TF (U. reticulata) to 42.86% TF (C. scalpeliformis) in Chlorophyta (Table 4). n-3 and n-6 PUFAs largely contributed to the higher amount of PUFAs in almost all analyzed samples (Table 4). Linoleic acid (C18:2n-6), α-linolenic acid (C18:3n-3), γ-linolenic acid (C18:3n-6) and docosahexaenoic acid (C22:6n-3) represented the predominant n-3 and n-6 PUFAs in Chlorophyta. However, in the Rhodophyta and Phaeophyta, arachidonic acid (C20:4n-6) and eicosapentaenoic acid (C20:5n-3) were the foremost PUFAs (data not shown). The sum of n-6 PUFAs was higher in the Rhodophyceae and Phaeophyceae. The proportions of n-3 PUFAs also followed the similar order and varied between 1.83% TF and 26.29% TF. The n-6/n-3 ratio ranged from 0.5 to 3.93:1 in the Rhodophyta, 1.81–5.15:1 in the Phaeophyta, and 1.44–8.51:1 in the Chlorophyta (Table 4). Exceptionally, the n-6/n-3 ratio in few algal species namely G. corticata (12.35), G. dura (27.65), G. debilis (18.82), and G. fergusonii (18.65) exceeds the recommended values. The PUFA/SFA ratio was found to be ≥0.4 in most of the seaweeds.
Indices
The AI and TI relating to nutritional factors linked with coronary diseases are presented in Table 4. Statistically significant differences (P < 0.01) were observed among the different algal species for these indexes. The values of AI followed the order Rhodophyta (0.38–2.87) > Phaeophyta (0.53–0.81) > Chlorophyta (0.86–1.61). TI also followed a similar order with values ranging from 0.52 to 2.66 (Rhodophyta), 0.50 to 1.20 (Phaeophyta), and 1.28 to 2.90 (Chlorophyta) with the exception of G. salicornia which had a TI value of 5.75. The UI values ranged from 50.63 to 257.07 (Rhodophyta), 154.49 to 202.83 (Phaeophyta) and 70.87 to 141.87 (Chlorophyta) (Table 4).
Principal component analysis
PCA was performed for minerals, antioxidant and FAs in order to examine their distribution among the three phyla. Full cross-validation was used in the validation model in accordance to Wang et al. (2009).
The PCA plot of the mineral data matrix (Fig. 2) depicts the distribution of the major macroelements (Na, K, Ca and Mg) and microelements (Fe, Zn, Mn and Cu). The first two principal components (PCs) explained 70% of variability present in the data set accounting PC1 for 50% (on x-axis) and PC2 20% (on y-axis) of the total variance. The y-axis separated all the Chlorophyta from the Rhodophyta with the exception of C. veravalnensis and C. scalpeliformis, indicating that the Chlorophyta had higher Fe, Cu, Mn, Ca and Mg contents. On the contrary, the Rhodophyta contained more macroelements such as K and Na but had lower micronutrients when compared with the other two phyla. The Phaeophyta being segregated near the origin are intermediate, with the exception of P. tetrastromatica that was an outlier and showed higher Zn.

PCA bi-plot of minerals distribution (NA, K, Ca, and Mg as macroelements; Fe, Zn, Cu, and Mn as microelements) of 22 macroalgal species indexed from 1 to 22 according to Table 1
The PCA bi-plot (Fig. 3) and correlation loading plot (Fig. 4) of the antioxidant data matrix illustrated the antioxidant potential of different seaweeds, highlighting the relationships among total phenolic content and other antioxidant activity assays. A close loading of TPC, TAC, DPPH and O •−2 (SOD) radical scavenging activity on the right along PC1 axis depicts a high correlation between them with an explanation of 70% and 18% by the first two principal components PC1 and PC2, respectively. Algal species belonging to the Rhodophyta formed a separate group and are located on the left along PC1 axis with the exception of G. acerosa and S. filiforme, indicating their lower antioxidant potential when compared to the green and brown algae. A high and significant correlation was demonstrated between TPC and TAC (Pearson correlation coefficient r = 0.83), further supported by the correlation loading plot as both were closely located to each other. However, the close loading of DPPH, SOD, RP distant from TPC on PC1 axis indicates a fair correlation with TPC as also demonstrated by the Pearson correlation coefficient of TPC and DPPH (r = 0.70) and TPC and SOD (r = 0.52) (Fig. 4).

PCA bi-plot for total phenolic content (TPC) and other antioxidative assay such as DPPH, superoxide radical (SOD) scavenging, reducing power (RP), and total antioxidant activity (TAC) of 22 macroalgal species indexed from 1 to 22 according to Table 1

Correlation loading plot for total phenolic content (TPC) and other antioxidative assay such as DPPH, superoxide radical (SOD) scavenging, reducing power (RP), and total antioxidant activity (TAC) of 22 macroalgal species indexed from 1 to 22 according to Table 1
The FA data matrix subjected to PCA analysis satisfactorily explained the variability present with PC1 accounting for 68% and PC2 for 22% of total variation. This analysis discriminates the three phyla with the Chlorophyta rich in MUFAs, Phaeophyta rich in n-3 PUFAs and Rhodophyta, especially Gracilaria sp., rich in n-6 PUFAs (Fig. 5a). Further, the opposite positions of UI and AI, TI on the loading plot (Fig. 5b) signifies their inverse correlation (Pearson correlation coefficient for UI and AI, r = −0.87; UI and TI, r = −0.74).

PCA score plot (a) and loading plot (b) of FAs groups with distribution of UI, AI, and TI indexes of 22 macroalgal species indexed from 1 to 22 according to Table 1
Discussion
Proximate composition and minerals
Measurable differences in nutritional composition were evident among the algal species investigated in the present study. The percentage of soluble carbohydrate was significantly higher than the reported content of 16–40% in the seaweeds collected from Hawaii (McDermid and Stuercke 2003) but were marginally lower than the values of 39–70% in Vietnamese seaweeds (Hong et al. 2007). The protein content varied significantly in this study and was comparable with that of protein-rich foods from terrestrial plants such as soya bean. The protein values were comparatively higher than the values (5.8–0.41% DW) reported by Hong et al. (2007) and Matanjun et al. (2008). The values obtained for lipids, in particular for K. alvarezii and Gracilarian sp., were similar to earlier report for Rhodophytes which ranged from 1.1 to 2.4% DW (McDermid and Stuercke 2003; Matanjun et al. 2009). However, the higher lipid content of Caulerpa species in the present study was significantly lower than the values (3.8–4.4% DW) for C. racemosa from Australia reported by Renaud and Luong-Van (2006).
The Na/K ratio with values ranging from 0.30 to 2.24 could be of interest from a nutritional point of view as the diet with balanced Na/K ratio is important for people who take diuretics to control hypertension and suffer from excessive secretion of potassium. The species in the present study can therefore help to provide balanced Na/K ratio diets in contrast to olives with Na/K ratio of 43.63 and sausages with 4.89 (Ortego-Calvo et al. 1993). The sum of macroelements (Na + K + Ca + Mg in % DW) in this study falls within the rage already reported for seaweeds (Rupérez 2002; Hong et al. 2007) and was higher when compared to land vegetables (USDA 2001) ranging from 11.48 to 29.91. The corresponding values for carrot (3.27), sweet corn (1.33), green peas (1.45), tomato (3.42), potato (6.01), and spinach (9.67) are conspicuously lower.
The uptake capacity for the trace metals in algae generally occurs in two ways. The first is the surface reaction and is independent of factors influencing metabolism such as temperature, light, pH, or age of the plant. This seems to be the main uptake mechanism for Zn. The second is a slower active uptake in which metal ions like Cu, Mn, Se, and Ni are transported across the cell membrane into the cytoplasm. Moreover, their uptake process is directly dependent on metabolic processes and also varies with changes in temperature, light, or age of the plant (Sanchez-Rodriguez et al. 2001). In humans, iron deficiency occurs mostly when the demand for iron is high, e.g., in growth, high menstrual loss, and pregnancy, and this can lead to anemia. The reported Mn content in seaweeds is in the range of 0.9–28.5 mg 100 g−1 (Tuzen et al. 2009) and 0.3–5.5 mg 100 g−1 in commercially available edible marine algae (van Netten et al. 2000; Rupérez 2002). Similarly, the Cu contents obtained in this study are in agreement with earlier reports (Rupérez 2002; Topcuoglu et al. 2003; Tuzen et al. 2009). The average content of microelements (Fe + Zn + Mn + Cu in mg 100 g−1 DW) followed the order Phaeophyta (46.25) > Chlorophyta (40.02) > Rhodophyta (27.93) which were markedly higher to any of the land vegetables except spinach (50.9 mg 100−1 g).
The content of Ni in seaweeds has been estimated in the range of 0.05–7.0 mg 100 g−1 (van Netten et al. 2000; Topcuoglu et al. 2003; Tuzen et al. 2009), while Se varied between 0.01 and 0.6 mg 100 g−1 (van Netten et al. 2000, Tuzen et al. 2009). Selenium is recognized as an essential micronutrient in animals and humans, playing important biological roles as antioxidant, anticarcinogen and regulator of thyroid hormone metabolism. Low concentrations of selenium can cause anomalies in organisms while high concentrations are toxic. For inorganic As, the values were similar to those found elsewhere (Nakajima et al. 2006; Tuzen et al. 2009). Moreover, the bioavailability of inorganic As may also be affected by the high fiber content in algae. The levels of trace elements detected are below the toxic limits recommended by EU Health and Consumer Directorate-General (2008) and thus can be supplemented in human foods.
The carbon/nitrogen (C/N) ratio is used as an index for assessing the nutritional quality of food. In the present study, the average C/N ratio did not differ significantly among the algal groups and was highest in the Phaeophyta. Sulfate is a typical component of marine algal polysaccharides derived from fucans and galactans in brown and red seaweeds, respectively. Such sulfated mucilages are not found in land plants. The higher average sulfate content of the Phaeophyta and Chlorophyta is consistent with earlier reports of Rupérez (2002) with 1.3–5.9% sulfate in edible red and brown seaweeds.
Antioxidant properties
In the present study, extraction was carried out in methanol for 72 h in dark since this has been shown to give the highest antioxidant activity in many seaweed species (Matanjun et al. 2008). Moreover, Lapornik et al. (2005) reported that the yield of polyphenols in alcohol extracts strongly increases with longer duration of extraction. It is assumed that the antioxidant properties of phenolics are related to the number of phenol rings that makes them more effective hydrogen donors and free radical quenchers. The phenolics can also act as metal chelators which, in turn, prevent the catalytic function of metal in the process of initiating radicals (Devi et al. 2008). The higher phenolic contents of brown algae specifically C. indica and S. tenerrimum in the present study support similar findings of Matanjun et al. (2008). A series of polyphenolic compounds such as catechins, flavonols, phlorotannins and fucoxanthin have been identified from methanolic extracts of red and brown algae (Matanjun et al. 2008; Ganesan et al. 2008; Kuda and Ikemori 2009). Further, in Cystoseira spp., tetraprenyltoluquinol derivatives (tocopherol-like compounds) have also been identified for their marked antioxidant potential.
The higher DPPH radical scavenging activity of C. racemosa, S. tenerrimum, C. indica, C. veravalnensis and G. acerosa extracts indicates the hydrogen-donating ability of these extracts. Devi et al. (2008), Wang et al. (2009), and Zubia et al. (2009) also reported higher DPPH scavenging capacity in brown and green seaweeds compared to red seaweeds. In the present study, superoxide radical scavenging activity was consistently higher in the Phaeophyta and Chlorophyta. It is considered that the superoxide radical scavenging activity of seaweeds is caused not only by phenolic compounds but also by other water-soluble compounds, such as peptides, fucoidan and Maillard reaction products (Kuda et al. 2006). The radical scavenging activities of ulvans and ulvan derivatives also have been reported (Qi et al. 2006). The total antioxidant activity increased with increasing concentration of the methanolic extract and was higher in the brown and green seaweeds, as also found by Ganesan et al. (2008).
FA composition and indices
In the studied seaweeds, the green algae U. rigida and U. reticulata had the highest proportion of oleic acid which is considered as a characteristic of the Chlorophyta. The PUFA content (mainly arachidonic and eicosapentaenoic acids) in the Rhodophytes was similar to previous reports for red algae from the Bohai Sea (Li et al. 2002). Interestingly, the total sum of SFA, MUFA and PUFA in the present study followed the order as SFA > PUFA > MUFA. In contrast, the warm water macroalgal species have been reported to have higher SFAs and MUFA with relatively lower amount of PUFA than cold-water species (Khotimchenko 2003). Also, the sum of n-6 PUFAs being higher in Rhodophycean and Phaeophycean was in accordance with previous studies where n-3 and n-6 PUFAs were recorded as the dominant fraction of total FA in Rhodophyta and Phaeophyta (Dawezynski et al. 2007; Khotimchenko et al. 2002). These two classes of PUFA have opposing physiological functions, but their balance is important for normal growth and development and is beneficial for the prevention of cardiovascular and other chronic diseases such as diabetes, hypertension and autoimmune diseases. The n-6/n-3 ratio in this study ranged from 0.5 to 8.51 which is within the WHO prescribed standard of <10.
The fatty acid composition of the dietary fats and, particularly, of some individual fatty acids are of great importance in human nutrition and health. Low intake of saturated fat and an increased PUFA-to-SFA ratio are associated with a lower risk of human coronary heart disease. Thus, the PUFA/SFA ratio is one of the main parameters used to assess the nutritional quality of the lipid fraction of foods. In present study, this ratio was found to be ≥0.4 in most of the seaweeds and is within nutritional guidelines that recommend a PUFA/SFA ratio above 0.4 (Wood et al. 2004). The observed AI and TI value of <3 in most of the seaweeds in the present study was mainly due to the n-3 PUFA content and lower n-6/n-3 PUFA ratios. These results for the TI are important because thrombosis is a central event in atherosclerosis. TI values obtained in this study are similar to the values for products such as lamb (1.58), bovine meat (1.08), lean pork (1.37) and milk-based products (2.1). Therefore, although the addition of seaweeds to meat products may be useful for technological reasons such as gel-forming and water-retaining capacity, the addition of algal oils could be a more satisfactory strategy for the development of healthier lipid formulations. Recently, Lopez-Lopez et al. (2009) demonstrated that addition of edible seaweeds such as Sea Spaghetti, Wakame, and Nori as ingredients to meat product improves its thrombogenic and atherogenic potential and thus help in the quest for developing healthier lipid formulations. Further, the higher UI values in the seaweed lipids indicated a higher degree of unsaturation.
Principal component analysis that provides an interpretable overview of the main information enclosed in a multidimensional data set clearly showed a higher metal uptake capacity for Fe, Cu, Mn, Ca and Mg in the Chlorophyta and for K and Na in the Rhodophyta. Thus, rhodophycean algae could be of use for preparing the K salts from marine origin. The close loading of TPC and TAC coupled with their significant positive correlation particularly in the Phaeophyceae suggests that polyphenols are the principal contributors to the antioxidant activity. A significant positive correlation has also been observed between TPC and antioxidant activity of different seaweed extracts by many researchers (e.g., Kuda and Ikemori 2009; Wang et al. 2009). PCA further discriminates the Chlorophyta, which is rich in MUFAs, from the Phaeophyta and Rhodophyta, which are rich in n-3PUFAs and n-6 PUFAs, respectively. Thus, supplementation of human diets with seaweeds rich in natural antioxidants and other nutritional elements including PUFAs could provide a health benefit for consumers.
References
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Biophysiol 37(8):911–915
Dawezynski C, Schubert R, Jahreis G (2007) Amino acids, fatty acids and dietary fiber in edible seaweed products. Food Chem 103:891–899
De Lorenzo A, Petroni ML, De Luca PP, Andreoli A, Morini P, Iacopino L, Innocente I, Perriello G (2001) Use of quality control indices in moderately hypocaloric Mediterranean diet for treatment of obesity. Diabet Nutr Metab 14:181–188
Devi KP, Suganthy N, Kesika P, Pandian SK (2008) Bioprotective properties of seaweeds: in vitro evaluation of antioxidant activity and antimicrobial activity against food borne bacteria in relation to polyphenolic content. BMC Complement Altern Med 8:38–49
Duan XJ, Zang WW, Li XM, Wang BG (2006) Evaluation of antioxidant property of extract and fractions obtained from a red alga, Polysiphonia urceolata. Food Chem 95:37–43
Dubois M, Giles KA, Hamilton KS, Rebers PA, Smith F (1956) Colorimetric method for the determination of sugar and related substances. Anal Chem 18:350–356
EU Health and Consumer Directorate-General (2008) Council directive on nutrition labeling for food stuffs 2008/100/EC. Official Journal of the European Communities. No. L285/9. http://ec.europa.eu/food/food/labellingnutrition/nutritionlabel/indexen.htm
Farag RS, El-Baroty GS, Basuny AM (2003) The influence of phenolic extracts obtained from the olive plant (cvs. Picual and Kronakii), on the stability of sunflower oil. Int J Food Sci Technol 38:81–87
Ganesan P, Chandini SK, Bhaskar N (2008) Antioxidant properties of methanol extract and its solvent fractions obtained from selected Indian red seaweeds. Bioresour Technol 99(8):2717–2723
Herrero M, Cifuentes A, Ibanez E (2006) Sub- and super-critical fluid extraction of functional ingredients from different natural sources: plants, food-by-products, algae and microalgae—a review. Food Chem 98:136–148
Hong DD, Hein HM, Son PN (2007) Seaweeds from Vietnam used for functional food, medicine and bio-fertilizer. J Appl Phycol 19:817–826
Khotimchenko SV (2003) Fatty acids of species in the genus Codium. Bot Mar 46:456–460
Khotimchenko SV, Vaskovsky VE, Titlyanova TV (2002) Fatty acids from the Pacific coast of North California. Bot Mar 45:17–22
Kordali S, Cakir A, Mavi A, Kilic H, Yildirim A (2005) Screening of chemical composition and antifungal and antioxidant activities of the essential oils from three Turkish Artemisia species. J Agric Food Chem 53:1408–1416
Kuda T, Ikemori T (2009) Minerals, polysaccharides and antioxidant properties of aqueous solutions obtained from macroalgal beach-coast in the Noto Peninsula, Ishikawa, Japan. Food Chem 112:575–581
Kuda T, Hishi T, Maekawa S (2006) Antioxidant properties of dried product of ‘haba-nori’, an edible brown alga, Petalonia binghamiae (J. Agaradh) Vinogradova. Food Chem 98:545–550
Lapornik B, Prošek M, Wondra AG (2005) Comparison of extracts prepared from plant by-products using different solvents and extraction time. J Food Eng 71:214–222
Lee SB, Lee JY, Song DG, Pan CH, Nho CW, Kim MC (2008) Cancer chemopreventive effects of Korean seaweed extracts. Food Sci Biotechnol 17(3):613–622
Li X, Fan X, Han L, Lou H (2002) Fatty acids of some algae from the Bohai Sea. Phytochemistry 59:157–161
Lim SN, Cheung PCK, Ooi VEC, Ang PO (2002) Evaluation of antioxidative activity of extracts from brown seaweed, Sargassum siliquastrum. J Agric Food Chem 50:3862–3866
Lopez-Lopez I, Bastida S, Ruiz-Capillas C, Bravo L, Larrea MT, Sanchez-Muniz F, Cofrades S, Jimenez-Colmenero F (2009) Composition and antioxidant capacity of low-salt meat emulsion model system containing edible seaweeds. Meat Sci 83:492–498
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193:265–275
Matanjun P, Mohamed S, Mustapha NM, Muhammad K, Ming CH (2008) Antioxidant activities and phenolics content of eight species of seaweeds from north Borneo. J Appl Phycol 20:367–373
Matanjun P, Mohamed S, Mustapha NM, Muhammad K (2009) Nutrient content of tropical edible seaweeds, Eucheuma cottonii, Caulerpa lentillifera and Sargassum polycystum. J Appl Phycol 21:75–80
McDermid KJ, Stuercke B (2003) Nutritional composition of edible Hawaiian seaweeds. J Appl Phycol 15:513–524
Nakajima Y, Endo Y, Ionue Y, Yamanaka K, Kato K (2006) Ingestion of Hijiki seaweed and risk of arsenic poisoning. Appl Organomet Chem 20:557–564
Ortego-Calvo JJ, Mazuelos C, Hermosin B, Saiz-Jimenez C (1993) Chemical composition of Spirulina and eukaryotic algae food products marketed in Spain. J Appl Phycol 5:425–435
Pisani P, Bray F, Parkin DM (2002) Estimates of the world-wide prevalence of cancer for 25 sites in the adult population. Int J Cancer 97:71–81
Prieto P, Pineda M, Aguilar M (1999) Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: specific application to the determination of vitamin E. Anal Biochem 269:337–341
Puja K, Manoj K, Vishal G, Reddy CRK, Jha B (2010) Tropical marine macroalgae as potential sources of nutritionally important PUFAs. Food Chem 120:749–757
Qi H, Zhang Q, Zhao T, Hu R, Zhang K, Li Z (2006) In vitro antioxidant activity of acetylated and benzoylated derivatives of polysaccharide extracted from Ulva pertusa (Chlorophyta). Bioorg Med Chem Lett 16:2441–2445
Renaud SM, Luong-Van JT (2006) Seasonal variation in the chemical composition of tropical Australian marine macroalgae. J Appl Phycol 18:381–387
Rupérez P (2002) Mineral content of edible marine seaweeds. Food Chem 79:23–26
Sanchez-Rodriguez I, Huerta-Dia MA, Choumiline E, Holguin-Quinones O, Zertuche-Gonzalez JA (2001) Elemental concentrations in different species of seaweeds from Loreto Bay, Baja California Sur, Mexico: implication for the geochemical control of metals in algal tissue. Environ Pollut 114:145–160
Santoso J, Gunji S, Yoshie-Stark Y, Suzuki T (2006) Mineral contents of Indonesian seaweeds and mineral solubility affected by basic cooking. Food Sci Technol 12:59–66
Topcuoglu S, Guven KC, Balkis N, Kirbasoglu C (2003) Heavy metal monitoring of marine algae from the Turkish Coast of the Black Sea, 1998–2000. Chemosphere 52:1683–1688
Tuzen M, Verep B, Ogretmen AO, Soylak M (2009) Trace element content in marine algae species from the Black Sea, Turkey. Environ Monit Assess 151:363–368
USDA (2001) Agricultural Research Service. Nutrient Database for Standard Reference, Release 14
van Netten C, Hoption Cann SA, Morley DR, van Netten JP (2000) Elemental and radioactive analysis of commercially available seaweed. Sci Total Environ 255:169–175
Wang T, Jonsdottir R, Olafsdottir G (2009) Total phenolic compounds, radical scavenging and metal chelation of extracts from Icelandic seaweeds. Food Chem 116:240–248
Wood JD, Richardson RI, Nute GR, Fisher AV, Campo MM, Kasapidou E (2004) Effects of fatty acids on meat quality: a review. Meat Sci 66:21–32
Yen GC, Chen HY (1995) Antioxidant activity of various tea extracts in relation to their antimutagenicity. J Agric Food Chem 43:27–32
Zubia M, Fabre MS, Kerjean V, Lann KL, Stiger-Poureau V, Fauchon M, Deslandes E (2009) Antioxidant and antitumoural activities of some Phaeophyta from Brittany coasts. Food Chem 116:693–701
Acknowledgements
The financial support received from CSIR (NWP-018) is gratefully acknowledged. The first author (MK) and second author (PK) gratefully acknowledge the CSIR, New Delhi, for awarding the Senior and Junior Research Fellowships. The third (NT), fourth (MKS), and fifth (VG) authors also expresses their gratitude to CSIR (RSP-016) and Department of Science and Technology, New Delhi, for financial support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kumar, M., Kumari, P., Trivedi, N. et al. Minerals, PUFAs and antioxidant properties of some tropical seaweeds from Saurashtra coast of India. J Appl Phycol 23, 797–810 (2011). https://doi.org/10.1007/s10811-010-9578-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10811-010-9578-7