
Abstract
To conserve areas and species threatened by immediate landscape change requires that we make planning decisions for large areas in the absence of adequate data. Here we study the utility of broad-scale landscape metrics as predictors of species occurrence, especially for remote areas where there is a need to make the most of limited spatial and biological data. Bonobos (Pan paniscus) are endangered great apes endemic to lowland forests of the Democratic Republic of Congo. They are threatened by bushmeat hunting that is exacerbated by habitat fragmentation through slash-and-burn agriculture and timber harvest. We developed four landscape metrics —edge density (ED), COHESION, CONTAGION, and class area (CA)— that may serve as surrogates for measuring accessibility of areas to hunting in order to predict relative bonobo-habitat suitability. We calculated the metrics for the Maringa-Lopori-Wamba (MLW) landscape and evaluated them for utility in predicting bonobo-nest occupancy based on 2009 field data. Cross-validations showed that all four metrics performed similarly. However, forest ED was arguably the best predictor, with an overall classification accuracy of 72.1% in which 85% of known nest blocks (N = 124) were classified correctly. We demonstrated that for a relatively intact landscape and a mobile forest-dwelling species that is fairly tolerant of forest openings, forest fragmentation can still be an important predictor of species occurrence. We suggest that ED can be helpful when mapping bonobo habitat in MLW and can aid landscape-planning and conservation efforts. Our approach may be applied to other edge-sensitive species, especially where high-resolution data are deficient.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Remote areas present a challenge for understanding species–habitat relationships because both biological and habitat data are scarce and difficult to obtain. Predicting species distributions generally depends upon having two primary types of data: 1) geographic locations of the species in question and 2) variables associated with those locations and the entire area of interest, such as vegetation cover type, soil type, or elevation (Austin and Meyers 1996). Statistical relationships are investigated to determine the likelihood of species occurrence given a range of environmental conditions (Austin 1998; Elith et al. 2006; Ferrier et al. 2002; Franklin 1995; Guisan and Zimmermann 2000). Species location data can be scarce in remote areas owing to inaccessibility, and this inaccessibility also complicates the collection of observations to ground-truth the classification of remotely sensed data that may exist, such as satellite imagery. Yet for endangered species, conservation planning must proceed in these data limited scenarios and such planning relies on reasonable estimates of species distributions.
Bonobos (Pan paniscus) epitomize the remote-area challenge (Nishida 1972). Bonobos occur naturally at low densities and are difficult to locate (Badrian et al. 1981; Kano 1984; Mohneke and Fruth 2008) in the rain forests of the Democratic Republic of Congo (DRC). Although decades of effort have been invested in wild bonobo studies (Furuichi and Thompson 2008; Susman 1984; White 1996), much of that work has been on habituated populations (Furuichi 1987; Furuichi and Thompson 2008; Furuichi et al. 1998; Hohmann and Fruth 2008; Kano 1992). Surveys of unhabituated bonobos tend to focus on the daily nest structures they build, because in areas where they are hunted, bonobos tend to avoid humans (Fruth et al. 2008; Hart et al. 2008; Kano 1984; Mohneke and Fruth 2008; Reinartz et al. 2006, 2008; Van Krunkelsven 2001; cf. Grossman et al. 2008). Persistent civil strife, limited road infrastructure, and food insecurity contribute to the low accessibility of the bonobo range to scientists (Dupain and Van Elsacker 2001; Eba’a Atyi and Bayol 2009; Grossman et al. 2008). The highly rural human population within the bonobo range generally sustains itself through unregulated expansion of slash-and-burn agriculture, bushmeat hunting, and forest-product use, e.g., firewood collection (Eba’a Atyi and Bayol 2009; Oates 1994; USAID 2010). These extractive activities impact wildlife and fragment Congolese rain forests, home to the only wild populations of bonobos in the world (Fruth et al. 2008; USAID 2010). Determining effects of these activities on bonobo distributions can guide future research and aid landscape planning efforts.
Ongoing conservation efforts are grappling to determine priority areas for research, monitoring, and protected-area designation for bonobos (Luetzelschwab 2007). To address the urgent call for landscape planning in the face of the aforementioned data limitations, there is a need to identify broad-scale landscape variables, derived from remotely sensed data, and test their ability to identify high-quality bonobo habitat. Although increased data collection would be ideal, we were interested in exploring the utility of broad-scale landscape metrics to bridge the gap, thereby providing a near-term solution until more species-location and high-resolution data become available. To develop a bonobo-relevant metric, we considered aspects of bonobo habitat expected to be both important to bonobos and detectible by satellite imagery. Timber harvest and slash-and-burn agriculture remove trees and forest cover that bonobos use as nesting, foraging, and shelter habitat (Badrian et al. 1981; Kano 1984; Kano and Mulavwa 1984; Oates 1994). Further, timber inventories (conducted on cut transects), road networks, logging operations, and small farms penetrate the dense forest with linear openings that facilitate hunter access to bonobos (Dupain and Van Elsacker 2001; Dupain et al. 2000; Oates 1994; Wilkie et al. 1992). It is possible then that logging and farming not only reduce bonobo habitat through tree removal, but also that these activities may actually lead to increased harvest rates of bonobos. Although not a primary species for subsistence consumption, bonobos are eaten and can be sold for considerable profit in urban markets or as part of the pet trade (Dupain and Van Elsacker 2001; Dupain et al. 2000). While satellite imagery cannot detect hunting explicitly, remote sensing can capture the forest fragmentation that exacerbates hunting and pet trade activities. In fact, habitat fragmentation and hunting are now considered principal threats to primates in general (Arroyo-Rodriguez and Mandujano 2009), and have been regarded as such for bonobos specifically for ≥25 yr (Kano 1984). Therefore a metric that captures this fragmentation may be a useful predictor of bonobo occurrence.
This study focuses on the process of creating four bonobo-specific fragmentation metrics from available Landsat Thematic Mapper (TM) products. To determine the utility of these metrics to identify potential habitat for bonobos in the face of limited available high-resolution spatial data, we evaluated their ability to predict bonobo nest occurrence in the Maringa-Lopori-Wamba (MLW) landscape and discuss their potential value for broad scale habitat-suitability modeling and management applications.
There are many potential metrics one could use to estimate habitat fragmentation. Selection of a fragmentation metric is challenging because quantifications of fragmentation and habitat loss have been shown to be confounded (Fahrig 2003; Neel et al. 2004) in that fragmentation itself is caused by dispersed habitat loss, resulting in a correlation between the two measures. Neel et al. (2004) examined numerous metrics across gradients of aggregation and percent habitat (area). Aggregation is a measure of the degree to which pixels of a focal class (say, forest) are spatially clustered. Conceptually, aggregation is similar to connectivity and is essentially the inverse of fragmentation. Neel et al. (2004) showed that certain landscape metrics purported to measure aggregation sometimes correlate more highly with percent habitat (P) than with aggregation. This nonintuitive behavior adds to the difficulty in selecting appropriate metrics.
We evaluated forest edge density (ED, the linear edge between forest and nonforest in a given area), COHESION, CONTAGION, and class area (CA) of forest, as predictors of bonobo occurrence (Table I). We expected ED (McGarigal and Marks 1995) to be useful as a broad landscape metric that simultaneously captures the importance of intact forest (low ED) and the concomitant negative impacts of forest loss and forest fragmentation (high ED). A strength of the conceptually intuitive ED metric is that it has a strong negative correlation (Kendall's τ =−0.79) with aggregation (Neel et al. 2004), which translates to a positive correlation with fragmentation. However, there is no perfect metric. The weakness of ED is that it exhibits a parabolic response in relation to percent-habitat, P, hypothetically represented in Fig. 1. This means that as area of target habitat nears 50% of the landscape, the potential for high ED will be highest (Li et al. 1993), e.g., complete disaggregation of forest pixels is possible, as in a checkerboard. Although this parabolic behavior may at first seem problematic, because there is the potential for similar ED values at different levels of forest disturbance, it is still true that complexity of patch edge influences ED such that for the same P, convoluted edges result in higher ED than simple edges (Fahrig 2003; Hargis et al. 1998). Therefore, ED can highlight differences in edge within a narrow range of P values. For landscapes with a wide range of P values, one could multiply ED by CA of forest such that the interaction term would capture the entire 0–100% range. Either of these approaches is likely to capture useful information on landscape pattern for species responding to edge effects (Chalfoun et al. 2002; Donovan et al. 1997). In our highly forested study area, we expected P values predominantly in the upper tail of the ED-P curve (Fig. 1) and for bonobo occurrence to decline with increasing ED; therefore, for simplicity we used ED alone.

A hypothetical representation of the parabolic nature of edge density (ED) over the percent habitat (P) gradient. ED values tend to be lower for both low and high values of P.
We chose COHESION because it proved to be a useful predictor of dispersal for another highly mobile forest-associated species, the northern spotted owl (Strix occidentalis caurina: Schumaker 1996; UMass 2000). However, COHESION is nonintuitive because, although it was originally conceived as a measure of aggregation or connectedness, it was later found to correlate more strongly with quantity of habitat (τ = 0.88) rather than aggregation (τ = 0.12) (Neel et al. 2004).
We chose CONTAGION (Li and Reynolds 1993) because it was originally created to capture both the degree to which habitat patch types are mixed and the spatial distribution of patch types at the landscape level (McGarigal and Marks 1995). However, the ability of CONTAGION to represent spatial distribution of habitat patches is disputed (Hargis et al. 1998). Generally, lower CONTAGION indicates a more mixed pattern of different patch types and higher CONTAGION indicates more aggregation of like patch types in a landscape. We expected a positive correlation between bonobo occupancy and both COHESION and CONTAGION.
We chose CA to compare fragmentation metrics to simple habitat loss and to evaluate where our study landscape, MLW, resides on the P gradient.
Methods
Study Area
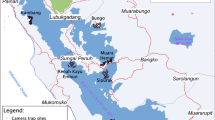
The study area, MLW, is designated as a conservation landscape by the Central African Regional Program for the Environment (CARPE), a branch of the U.S. Agency for International Development (USAID) (Hickey and Sidle 2006). MLW (Fig. 2) was selected as a CARPE landscape specifically for the conservation of bonobos in conjunction with alleviation of poverty (Hickey and Sidle 2006). MLW is ca. 74,000 km2 and characterized by large areas of intact lowland rain forest, human settlement, slash-and-burn agriculture, and numerous large timber concessions (totaling >6000 km2) in different states of harvest rotation or harvest planning (Dupain et al. 2009).

MLW landscape, Democratic Republic of Congo, selected focal areas, and location of line transects for bonobo nest surveys conducted in 2009.
Focal Area Selection
To determine the range of fragmentation values in MLW (of four landscape metrics with values for each pixel based on a 20-km2 window around each pixel) in relation to land use, we selected three focal areas (Fig. 2). Each focal area was ca. 430 km2 in size and represented one of the three most typical land use categories: minimum use, logging use, and human settlements, respectively. We selected the minimum-use area based on continuous high canopy cover, absence of roads, and distance from detectable human activity. We assumed this minimum-use area represents optimal bonobo habitat and likely provides a bookend-reference point for greatest habitat quantity/quality based on least fragmentation achievable in MLW. We selected the logging-use area for its regular grid-like pattern of logging-access roads in an area with no detectable human settlements. We assumed the logging-use area represents an area in which bonobo-habitat value is reduced by the level of fragmentation. Such fragmentation may allow increased hunter access and, subsequently, either increased harvest of bonobos or avoidance of the area by bonobos. We selected the human-settlement area for the known high human population, large interruptions in forest canopy due to the presence of villages and agriculture, and the absence of nearby logging. This human-settlement area represents the opposite end of the spectrum from the minimum-use area and provides a reference for maximal levels of fragmentation within MLW as of 2000.
Development of Base Forest-Cover Layers
We used seven scenes of Landsat TM imagery c. 2000 as classified for the USAID-CARPE decadal forest change mapping project with a pixel size of 57 × 57 m (Hansen et al. 2008). The original classification portrayed each pixel as the likelihood (1–99%) of having ≥60% canopy cover (henceforth termed “forest”). Hansen et al. (2008) validated the classification using MODIS (MODerate Resolution Imaging Spectroradiometer) data, with a coarser resolution of 231 m, which is atypical of most validation processes. Ideally, finer grained imagery or field reconnaissance would inform the validation process; but again, this remote area was deficient in data, including high-resolution data. Although more recent imagery (c. 2010) has since been classified in a similar manner, it was not available at the time of these analyses. We expect the 9-yr difference between satellite images and field data is negligible for these analyses because the amount of forest loss between 2000 and 2010 was <0.44% of MLW (OSFAC 2010).
To calculate landscape metrics, we first had to establish binary habitat types from the original classification. Using ESRI’s ArcMap version 9.3 we reclassified the probability of being forested into a binary raster (FOR), where 0 = unforested (0–30% likelihood of being forest) and 1 = forested (31–99% likelihood of being forest). Likelihood of forest cover was allowed to range widely (31–99%) for the definition of forested habitat because bonobos are tolerant of low canopy cover and openings (Thompson 1997; Uehara 1990) in the absence of human activity, i.e., hunting, and have been documented in some forest-savannah mosaics (Inogwabini et al. 2008). In addition, we compared several different thresholds to Google® Earth imagery in the few locations where high-resolution data were available, and the 30% threshold appeared to best capture forest/nonforest habitats. Because we had access to reasonably good spatial data for roads and rivers (CARPE-UMD 1997; Lehner et al. 2006) that were not always classified as nonforest in many areas of the Landsat-derived data, we created two additional forest/nonforest layers on which to base landscape metrics to test the value of this additional information. Owing to documented reduced bonobo numbers around roads (Dupain and Van Elsacker 2001; Dupain et al. 2000; Horn 1980), we buffered roads 100 m on either side and added the buffered roads to the unforested class, to create a second base layer (referred to as RD). Finally, because of increased access and hunting activity near rivers, we buffered both roads and rivers by 100 m and added those buffered areas to the unforested class, for a third base layer (referred to as RR, for roads and rivers).
Calculating Landscape Metrics
Because we were interested in comparing the ability of four landscape metrics to predict bonobo presence across all of MLW, including sites we never surveyed, we needed to build spatially explicit raster layers of each metric across the entire landscape. We conducted a moving window analysis in FRAGSTATS Version 3.3 (McGarigal et al. 2002) to calculate ED, COHESION, CONTAGION, and CA on each of the above three base forest-cover layers (FOR, RD, and RR). We assumed that home-range size is the scale at which bonobos respond to fragmentation, and therefore applied a radius of 2.524 km to the moving windows to approximate the mean area (20 km2) of a bonobo group home range (Hashimoto et al. 1998). We then assigned the value of each metric within a given window to the centroid of that window. By stepping these windows across the entire landscape, this procedure results in a raster with a home-range scaled landscape metric assigned to every pixel. We also ran moving-window analyses on the three focal areas to investigate the nature of the metrics along a relative continuum of impacted areas. We employed a four-cell rule for neighborhood size.
We reported ED as a positive number, with larger values indicating greater fragmentation. An ED value of 1 m/ha converted to 1 km of edge/10 km2 and equated to 2 km of edge across a given 20-km2 window. COHESION of forest was a positive number <100, with higher values indicating more connected (less fragmented) forest. Similarly, CONTAGION was a positive number ≤100, with larger numbers indicating more aggregation of like patch types. CA of forest was simply the forested area in ha within each moving window and ranged from 0 to 2000 ha. To investigate if ED is a potentially useful metric for this landscape, we converted CA to percent-forested habitat, P. This allowed us to assess whether MLW represents a relatively narrow portion of the P gradient (Hargis et al. 1999; Neel et al. 2004).
Field Verification
We randomly stratified survey sites a priori to represent a range of fragmentation levels, including logged and unlogged areas. Because our premise is that bonobos may avoid highly fragmented areas owing to the potential for increased hunting pressure in those areas, we also stratified by protected status, distance-from-fire (a proxy for villages), and distance-from-river (a proxy for human access sites). In this way, we ensured that fragmentation was explored across a gradient of potential hunting pressure. In 2009, we conducted line-transect surveys for bonobo nests (Fig. 2) using double-independent observer techniques (Williams et al. 2002). We generated start and end points of all transects randomly in ArcMap (ESRI, version 9.3) for each strata described. We completed ca. 73 km of line-transect surveys, all of which we surveyed twice, once each by two separate observation teams. We recorded the geographic coordinates of the transect point located perpendicularly to both singly and doubly observed nests, as well as other sign (other sign not analyzed here). We complied with protocols approved by the University of Georgia’s Institutional Animal Care and Use Committee (AUP no. A2009-10042) and adhered to the legal requirements of the DRC.
Designation of Nest Blocks and Random Blocks
Bonobo nests tend to occur in groups and would be expected to be clustered on the landscape (Mulavwa et al. 2010). To reduce this spatial autocorrelation, we decreased the resolution of our individual nest locations to 57 × 57 m blocks (the same resolution as our Landsat TM imagery classification; Hansen et al. 2008). We termed a block with one or more nests in it a nest block. For a comparison set of random (non-nest) blocks that we surveyed with effort equal to nest blocks, we created random points on the transects that fulfilled the criteria of being ≥100 m both from nest blocks and from each other. The 134 random points were distributed proportionally to each transect based on its length compared to the total length of all transects surveyed. It is worth noting that these random blocks do not equate to known absences, because bonobo nests could have occurred there in the past. Because bonobo nests decay between about 75 and 99 days (Mohneke and Fruth 2008), bonobos could have used these random blocks any time >3 mo before our surveys.
Logistic Regression Modeling of Bonobo Nest Occurrence
Using logistic regression analysis (Neter et al. 1989) in SAS (v. 9.1), we examined the individual relationships between nest-block occurrence and ED, COHESION, CONTAGION, and CA. We input 124 nest blocks (1), 134 random blocks (0), and their corresponding landscape-metric value into logistic regression models (one model for each landscape metric, separately). To test for multicollinearity, we calculated Pearson’s correlations (r) between all pairs of variables to assess if multiple landscape metrics could be included in a multivariate model. We ranked the metrics based on their leave-one-out predictive error rates to select one for mapping (Kearns and Ron 1999; Kearns et al. 1997). In logistic regression the coefficients are expressed in log odds; therefore, to calculate an odds ratio, the parameter estimate for the coefficients must be back-transformed with the exponential function, e x, where x is the logistic parameter estimate. This procedure allows inference of the relationship between the predictor and response variables (Hosmer and Lemeshow 1989). Therefore, we calculated odds ratios for the selected metric to infer the direction and magnitude of the relationship with nest-block occurrence. Odds ratios >1 indicate positive relationships, such that with each unit increase in the variable, the probability of occurrence is e x times greater. Odds ratios <1 indicate a negative relationship and are interpreted more easily by taking the inverse and stating “nest blocks are 1/e x times” less likely to occur with each unit increase in the parameter.
Fragmentation Thresholds
While landscape variables are often useful for habitat modeling in their continuous form, it is often necessary to choose a threshold for visualization and planning. To that end, we produced fragmentation maps based on three threshold choices. The Continuous map (no threshold) simply depicts the continuous gradient of fragmentation values across MLW. Next, because conservation planners may be interested in a distinct demarcation between acceptable and unacceptable amounts of canopy alteration in bonobo habitat, we applied two fragmentation thresholds resulting in binary maps. To assign defensible thresholds, we evaluated fragmentation values using Jenks’ natural breaks (Jenks 1967) to identify a natural break in the landscape metric data (Habitat Threshold) and we determined the maximal fragmentation value where bonobo nest blocks were found (Nest Threshold). The Jenks procedure defined categories by maximizing interclass variance and minimizing intraclass variance for ED across MLW. The Habitat Threshold allowed the data on forest pattern to define the threshold, whereas the Nest Threshold allowed the data about bonobo nest occurrence to define the threshold.
Results
Landscape Metrics
All four landscape metrics (Fig. 3) correlate highly with each other (Pearson's |r| > 0.95). The strongest correlations are between CONTAGION and CA (r = 0.989), followed by CONTAGION and COHESION (r = 0.987) and by CONTAGION and ED (r = −0.982) (N = 258 for all comparisons). Evaluating all metrics across all three base layers, we found ED consistently ranked as the first or second for correlation with bonobo nest occurrence (r = −0.5 for ED-RD), whereas the other metrics frequently ranked 3rd or 4th. Further, leave-one-out cross validations also ranked ED the highest. Therefore, we discuss only ED in more detail. ED-RD values ranged from 0 to 104 m/ha across MLW (Fig. 3). The mean ED for MLW was 9.83 ± SE 0.003 m/ha. ED was higher (42.6 ± SE 0.06 m/ha) in the human-settlement area compared to all other focal areas (Fig. 4). The minimum-use area had virtually no ED (3.06 ± SE 0.015 m/ha), whereas the logging-use area had slightly higher ED vs. the entire MLW. The mean ED for each focal area was significantly different than that of MLW (p < 0.01 for all comparisons).

Maps of the four landscape metrics across MLW.

Mean ED (±SD) for the Maringa-Lopori-Wamba (MLW) landscape and selected focal areas. Focal area labels correspond as follows: Min=Minimum Use, Log=Logging Use, and Hum=Human Settlement; and (**) indicates significant difference from MLW at the p < 0.01 level.
The Jenks (1967) natural breaks procedure demonstrates that there is a natural break around 6.56 m/ha (equivalent to 13.12 km of edge within a 5.048-km diameter home range) that defines well the difference between minimum-use and logging-use areas (Fig. 5). The vast majority of pixels in the minimum-use area had an ED <6.56 m/ha, whereas the majority of the logging area had EDs between 6.56 and 18.71 m/ha. In addition, >60% of MLW had an ED <6 m/ha and >80% had an ED <19 m/ha. Further, we converted CA to percent habitat, P, and found that >90% of MLW was >80% forested, demonstrating that MLW represents a sufficiently narrow portion of the P gradient to warrant the use of ED as a measure of fragmentation. As we predicted, most areas were in the upper tail of the ED-P curve (Fig. 1); therefore ED can be expected to correlate negatively with percent-habitat and positively with forest disturbance in MLW (Neel et al. 2004).

Frequency of edge density values (per pixel based on a 20-km2 window around each pixel) for MLW and selected focal areas. Categories on the x-axis represent Jenks’ natural breaks (Jenks 1967) for ED across MLW.
Field Verification
During 2009, we completed ca. 73 km of line-transect surveys and recorded the geographic coordinates of 338 bonobo nests. We treated multiple nests occurring within a single 57-m pixel of our base GIS data layers as a single observation resulting in 124 total nest blocks.
Logistic Regression Modeling of Bonobo Nest Occurrence
Owing to the high correlations found among the landscape metrics, they appear to contain nearly the same information and therefore should not be included together in the same predictive model (Neter et al. 1989). Hence we were interested in ranking their classification accuracy to select a single best predictor. Leave-one-out cross validation performed on single-variable logistic regression models confirmed that ED, COHESION, CONTAGION, and CA predicted bonobo-nest occurrence similarly in all instances. Their predictive error rates, which reflect the sum of the false-positive (commission) and false-negative (omission) rates, ranged from 27.91% to 30.62%. We ranked all the metrics based first on predictive error (lower is better) and second on true positive rates (higher is better) (Table II). The top three predictors all had a predictive error rate of 27.91% and included ED calculated on FOR, ED calculated on RD, and CONTAGION calculated on FOR. Although we acknowledge the merit of CONTAGION as a fragmentation metric, as well as the strong similarity in predictive capability of all the calculated metrics, we selected ED for our maps because of both its high ranking here and its ease in interpretation over the other metrics. Note that the model containing CA-RD as a sole predictor had an exceptionally high true-positive rate of 95%; however, this model had a substantially higher commission error than the top three models (52% vs. 40%). Therefore, although the model looks strong initially, its ability to discriminate is marginal in comparison to the top models. Although we ran 12 separate logistic models, one for each landscape metric and base forest-cover layer, we summarize the results of the four models run on the intermediate representation of forest cover, the RD base layer (Table III). As mentioned earlier, all metrics performed similarly as predictors of bonobo nest occurrence, and ED ranked consistently the highest. As an example, a confusion matrix (Fig. 6) describes the strengths and weaknesses of the logistic ED-RD model in classifying bonobo occurrence.

Confusion matrix showing classification accuracy of the ED-RD model. Box (a) reflects that 105/124 = 84.7% of known nest blocks were correctly classified. Box (c) highlights that the greatest source of error in this model is in misclassifying 53/134 = 39.6% of blocks as nest-blocks, when in fact, no nest was found (commission error).
For proper inference we transformed the log odds parameter estimate (−0.255) in the ED-RD logistic model (e –0.255 = 0.775), and because 0.775 is <1 the odds ratio indicates a negative relationship between ED and nest-block occurrence. Therefore, nest blocks were (1/0.775) = 1.3 times less likely to occur for each 1-km increase of edge per 10 km2. Or more simply, about one-third fewer nests were expected to occur for each unit increase in ED.
Fragmentation Thresholds
We produced continuous ED and binary-thresholded ED maps for the three focal areas and for all of MLW (Fig. 7). The Habitat Threshold map uses a threshold ED of 6.56 m/ha, the natural break in ED values across the landscape and between minimum-use and logging-use areas. The Nest Threshold map uses a threshold ED of 12.256 m/ha, the highest ED value for a nest block from our field verification surveys.

Focal areas highlight differences between continuous and thresholded ED maps, and landscape-wide maps depict corresponding focal areas in light boxes from left to right: logging-use, minimum-use, and human-settlement areas. Thresholds accentuate areas where ED may be too high for bonobo nesting.
Discussion
Our comparison of prediction accuracy of landscape metrics derived from remotely sensed data demonstrated that fragmentation, no matter how we measured it, is a useful predictor of bonobo nest presence; therefore we encourage the use of a single, well-chosen fragmentation metric for use in multivariate bonobo distribution or habitat-suitability models. The four bonobo-relevant landscape metrics, each built on forest base-layers to represent the potential tolerance of bonobos to open canopy and edge effects, correlate highly with each other and performed similarly in predicting bonobo nest presence. We favor the use of ED because it ranked highest in leave-one-out cross-validations (Table II) and perhaps, more importantly, because it is the most intuitive representation of fragmentation (Table I). Our field surveys demonstrated that bonobo nest block occurrence was indeed higher where ED (fragmentation) was lower. We found ED, as we calculated it, to be a useful predictor of nest occurrence and potential bonobo nesting habitat. Although ED-RD is merely a single-variable model, it boasted 72.1% overall prediction accuracy and correctly classified 85% of nest blocks.
There is precedent for including ED in evaluations of sustainable management in multiowner landscapes (Gustafson et al. 2007) and as an indicator of conditions for edge-sensitive species (USFS 2004). With that in mind, future land managers may desire a binary ED value either for assigning areas worthy of protection or for assessing acceptable levels of canopy alteration in multiple-use landscapes, e.g., extractive zones or community-use zones (Hickey and Sidle 2006). Ultimately, selecting a threshold for management choices such as allowable canopy alteration is an arbitrary decision, yet to be defensible requires scientific rationale. Therefore, we explored the range of fragmentation (ED) values in relation to land use in MLW. We identified a natural break in MLW-wide ED values that corresponded to the difference between the minimum-use and logging-use areas. Our field data supported this break as biologically meaningful to bonobos because >92% of nest blocks had an ED <6.56 m/ha, the natural break, and no nests were observed in the logging area. Based on the natural break in ED values for the landscape and the ED values in nest blocks, we offer two thresholds that are supported by the data from MLW. The Habitat Threshold employed the natural break in ED values (6.56 m/ha) to define the threshold, whereas the Nest Threshold allowed the maximal ED (12.256 m/ha) found in a bonobo nest block to define the threshold.
Because conservation spending can depend heavily on visualization of habitat and species ranges (Halpern et al. 2006), we advocate careful selection of thresholds both for communicating results and for conservation planning. To display potentially acceptable and unacceptable amounts of canopy alteration in bonobo habitat, we produced maps of the Continuous ED metric and two alternate ED thresholds for consideration. The Continuous map allows visualization of how fragmentation changes across the landscape. This depiction can be satisfying in that it is easy to perceive the gradient of fragmentation intensity, differentiating areas that are highly fragmented from those that may be of conservation value. The continuous metric is also preferred for potential inclusion in bonobo-habitat suitability models, along with other covariates, e.g., landcover, elevation. Eventually, multivariate models will be needed that take several other such explanatory variables into account. All maps portray the range of fragmentation conditions present in MLW c. 2000. Each depicts very low levels of suitably continuous forest for bonobo nesting in the human-settlement area, which is supported by field reconnaissance near the village of Djolu (Hickey and Sidle 2006) and previous research (Kano 1984). Further, all maps portray suitably contiguous forest in the minimum-use area, which we interpret as a reasonable conclusion for a species that is rather plastic in its use of cover types (Thompson 1997; Uehara 1990) in the absence of hunting pressure.
The Habitat Threshold is useful for highlighting areas of intact forest that may be most important for conserving bonobos in MLW. The Habitat Threshold may at first appear a cautious estimate of bonobo tolerance to fragmentation; however, our nest surveys suggest that the Habitat Threshold likely is a plausible binary representation of bonobo-habitat suitability in MLW. For instance, we found no nests in the logging-impact area, which the Habitat Threshold essentially depicts as entirely fragmented, and <8% of nest blocks in areas with ED above the Habitat Threshold. The Habitat Threshold demonstrates the pervasive nature of human presence even in a remote area plagued by an unreliable transportation system (Hickey and Sidle 2006; USAID 2010). For an area boasting one of the last strongholds of bonobos in the world, the Habitat Threshold suggests that <62% of MLW is sufficiently unfragmented for bonobos. The Nest Threshold is a liberal threshold from a conservation perspective, resulting in a binary map showing more area with suitably low levels of forest fragmentation. Although decisions based on liberal thresholds may be criticized because they are prone to commission error (inclusion of unsuitable areas), they represent the best choice for describing all potential habitat given all observations. Further study of the relationship between ED and bonobo nest occurrence both in MLW and other areas is recommended to assess whether the relationship and relevant thresholds change temporally or regionally.
At a broad scale, ED is an effective landscape metric for estimating bonobo nest occurrence and therefore potential bonobo habitat. We suggest ED can be used to increase the efficiency of future bonobo surveys, by increasing survey effort in areas of lower ED and decreasing effort in areas of higher ED. Future surveys can continue to inform the relationship between ED and nest occurrence and can allow extrapolation to unsurveyed areas based on those ED values. We advise against zero effort in areas of higher ED because estimates of overall bonobo abundance or density rely on characterizing the areas of both low and high bonobo densities. Extrapolating high-density estimates across all areas would result in grave overestimates of bonobo abundance in a given region. ED appears well suited for predicting bonobo occurrence in the MLW, an area with documented hunting impact on the bonobo population (Dupain and Van Elsacker 2001), and may extrapolate well to other areas of similar hunting pressure. However, the predictive value of ED may be weaker in areas where hunting pressure is relatively low because, in the absence of hunting, bonobos are relatively tolerant of open canopies and have been documented in some forest-savannah (fragmented) mosaics (Inogwabini et al. 2008; Thompson 1997; Uehara 1990). Conversely, there could be specific locales within the bonobo range in which the forest is relatively intact, yet bonobos do not occur. This could be due to occasional targeted hunting of remote areas, or other habitat factors that remain unknown. However, these areas are rare enough in our data that they did not mask the relationship between nest presence and ED. To find these anomalous areas, and potentially discover other relevant controlling factors, it will be important to institute continued monitoring and communicate with biologists and local people alike. Further investigations may help elucidate this relationship.
We believe we are the first to demonstrate quantitatively that fragmentation can be an important predictor of species occurrence for primates while their habitat remains relatively intact. Across taxa, the preponderance of fragmentation studies focus on landscapes in the lower tail of the ED-P curve, showing that the dispersed loss of habitat is important in highly disturbed landscapes in which habitat occurs in isolated patches surrounded by a matrix of nonhabitat (Andrén 1994; Arroyo-Rodriguez et al. 2008; Fischer and Lindenmayer 2007). Our study takes a different approach, investigating the potential consequences of habitat fragmentation before the matrix transitions from habitat to nonhabitat (sometimes called perforation). We suspect that the mechanism by which fragmentation affects bonobo distributions is through increased bonobo avoidance of areas owing to increased hunter access and increased hunting mortality near linear openings. Our study supports findings that hunting activity increases near openings and results in lower nest occurrence (Reinartz et al. 2008). Those linear openings in the forest habitat are effectively detected by remote sensing and measured by ED. We surmise that ED is most useful for landscapes with a preponderance of percent-habitat values in just one tail of the ED-P curve (Fig. 1). In our landscape, the majority of areas were well over 80% forested, in the upper tail, where there exists a negative relationship between ED and percent habitat. While our analysis reinforces previous assertions of similarities among various measures of fragmentation and area (Hargis et al. 1998, 1999) and supports the suggestion that fragmentation may impact bonobo habitat suitability (Kano 1984), it also quantitatively describes applicability of landscape-level fragmentation metrics for great ape habitat and assesses the impacts of fragmentation on habitat suitability.
Given the lack of studies of landscape-scale fragmentation metrics relevant to primates (Arroyo-Rodriguez and Mandujano 2009), we believe we have offered an approach that can be applied to other taxa. Fragmentation metrics can be developed for a given species by classifying habitat specifically with that species’ needs in mind and by selecting a window size at the scale that the species likely responds to fragmentation (perhaps the scale of the species’ home range). These metrics can be ranked using field data to evaluate their utility in predicting species occurrence and made spatially explicit in maps. For land managers and conservation planners, we have outlined some defensible ways to identify thresholds of allowable canopy alteration for a given species based on an evaluation of fragmentation values across the landscape and in different land use categories, in combination with levels of fragmentation where the species shelters. When delineating such thresholds we suggest employing species occurrence records that likely indicate areas of quality habitat rather than areas used in a transient manner. For example, we used nests where bonobos seek shelter for the night. The appropriate type of sign will depend on individual species’ habits and needs. In addition, species-covariate relationships may change over time, especially as land use and climates shift; therefore repeated studies examining such relationships are warranted.
References
Andrén, H. (1994). Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos, 71, 355–366.
Arroyo-Rodriguez, V., & Mandujano, S. (2009). Conceptualization and measurement of habitat fragmentation from the primates’ perspective. International Journal of Primatology, 30, 497–514.
Arroyo-Rodriguez, V., Mandujano, S., & Benítez-Malvido, J. (2008). Landscape attributes affecting patch occupancy by howler monkeys (Alouatta palliata mexicana) at Los Tuxtlas, Mexico. American Journal of Primatology, 70, 69–77.
Austin, M. P. (1998). An ecological perspective on biodiversity investigations: examples from Australian eucalypt forests. Annals of the Missouri Botanical Garden, 85, 2–17.
Austin, M. P., & Meyers, J. A. (1996). Current approaches to modelling the environmental niche of eucalypts: Implication for management of forest biodiversity. Forest Ecology and Management, 85, 95–106.
Badrian, N., Badrian, A., & Susman, R. L. (1981). Preliminary observations on the feeding behavior of Pan paniscus in the Lomako Forest of central Zaire. Primates, 22, 173–181.
CARPE-UMD. (1997). Democratic Republic of Congo Roads. Vector digital data. CARPE baseline data. CARPE, University of Maryland. Available at: ftp://congo.iluci.org/CARPE_data_explorer/Products/drc_road.zip
Chalfoun, A. D., Thompson, F. R., III, & Ratnaswamy, M. J. (2002). Nest predators and fragmentation: a review and meta-analysis. Conservation Biology, 16, 306–318.
Donovan, T. M., Jones, P. W., Annand, E. M., & Thompson, F. R., III. (1997). Variation in local-scale edge effect: mechanisms and landscape context. Ecology, 78, 2064–2075.
Dupain, J., & Van Elsacker, L. (2001). The status of the bonobo in the Democratic Republic of Congo. In B. M. F. Galdikas, N. Erickson Briggs, L. K. Sheeran, G. L. Shapiro, & J. Goodall (Eds.), All apes great and small, Vol. 1: African apes (pp. 57–74). New York: Kluwer Academic/Plenum Press.
Dupain, J., Van Krunkelsven, E., Van Elsacker, L., & Verheyen, R. F. (2000). Current status of the bonobo (Pan paniscus) in the proposed Lomako Reserve (Democratic Republic of Congo). Biological Conservation, 94, 265–272.
Dupain, J., Nackoney, J., Kibambe, J., Bokelo, D., & Williams, D. (2009). Maringa-Lopori-Wamba landscape. In C. de Wasseige, D. Devers, P. de Marcken, R. Eba’a Atyi, R. Nasi, & Ph. Mayaux (Eds.), The forests of the Congo Basin—State of the forest 2008, Luxembourg: Publications Office of the European Union. doi: 10.2788/32259. Available at: http://carpe.umd.edu/resources/Documents/SOF_23_Maringa.pdf
Eba’a Atyi, R., & Bayol, N. (2009). The forests of the Democratic Republic of Congo in 2008. In C. de Wasseige, D. Devers, P. de Marcken, R. Eba’a Atyi, R. Nasi, & Ph. Mayaux (Eds.), The forests of the Congo Basin—State of the forest 2008. Luxembourg: Publications Office of the European Union. doi: 10.2788/32259.
Elith, J., Graham, C. H., Anderson, R. P., Dudík, R. P., Ferrier, S., Guisan, A., et al. (2006). Novel methods improve prediction of species’ distributions from occurrence data. Ecography, 29, 129–151.
Fahrig, L. (2003). Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution, and Systematics, 34, 487–515.
Ferrier, S., Watson, G., Pearce, J., & Drielsma, M. (2002). Extended statistical approaches to modeling spatial pattern in biodiversity in northeast New South Wales. I. Species-level modelling. Biodiversity and Conservation., 11, 2275–2307.
Fischer, J., & Lindenmayer, D. B. (2007). Landscape modification and habitat fragmentation: a synthesis. Global Ecology & Biogeography, 16, 265–280.
Franklin, J. (1995). Predictive vegetative mapping: geographic modelling of biospatial patterns in relation to environmental gradients. Progress in Physical Geography, 19, 474–499.
Fruth, B., Benishay, J. M., Bila-Isia, I., Coxe, S., Dupain, J., Furuichi, et al. (2008). Pan paniscus. In IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. Available at: www.iucnredlist.org. Accessed July 25, 2011.
Furuichi, T. (1987). Sexual swelling, receptivity, and grouping of wild pygmy chimpanzee females at Wamba, Zaire. Primates, 28, 309–318.
Furuichi, T., & Thompson, J. (2008). The bonobos: Behavior, ecology, and conservation. Developments in Primatology: Progress and Prospects. New York: Springer.
Furuichi, T., Idani, G., Ihobe, H., Kuroda, S., Kitamura, K., Mori, A., Enomoto, T., Okayasu, N., Hashimoto, C., & Kano, T. (1998). Population dynamics of wild bonobos (Pan paniscus) at Wamba. International Journal of Primatology, 19, 1029–1043.
Grossman, F., Hart, J., Vosper, A., & Ilambu, O. (2008). Range occupation and population estimates of bonobos in the Salonga National Park: Application to large-scale surveys of bonobos in the Democratic Republic of Congo. In T. Furuichi & J. Thompson (Eds.), The bonobos: Behavior, ecology, and conservation (pp. 189–216). New York: Springer.
Guisan, A., & Zimmermann, N. E. (2000). Predictive habitat distribution models in ecology. Ecological Modelling, 135, 147–186.
Gustafson, E. J., Lytle, D. E., Swaty, R., & Loehle, C. (2007). Simulating the cumulative effects of multiple forest management strategies on landscape measures of forest sustainability. Landscape Ecology, 22, 141–156.
Halpern, B. S., Pyke, C. R., Fox, H. E., Haney, J. C., Schlaepfer, M. A., & Zaradic, P. (2006). Gaps and mismatches between global conservation priorities and spending. Conservation Biology, 20, 56–65.
Hansen, M. C., Roy, D., Lindquist, E., Adusei, B., Justice, C. O., & Altstatt, A. (2008). A method for integrating MODIS and Landsat data for systematic monitoring of forest cover and change in the Congo Basin. Remote Sensing of Environment, 112, 2495–2513.
Hargis, C., Bissonette, J., & David, J. L. (1998). The behavior of landscape metrics commonly used in the study of habitat fragmentation. Landscape Ecology, 13, 167–186.
Hargis, C., Bissonette, J., & Turner, D. L. (1999). The influence of forest fragmentation and landscape pattern on American martens. Journal of Applied Ecology, 36, 157–172.
Hart, J. A., Grossman, F., Vosper, A., & Ilanga, J. (2008). Human hunting and its impact on bonobos in Salonga National Park, Democratic Republic of Congo. In T. Furuichi & J. Thompson (Eds.), The bonobos: Behavior, ecology, and conservation (pp. 245–271). New York: Springer.
Hashimoto, C., Tashioro, Y., Daiji, K., Enomoto, T., Ingmanson, E. J., Idani, G., & Furuichi, T. (1998). Habitat use and ranging of wild bonobos (Pan paniscus) at Wamba. International Journal of Primatology, 19, 1045–1060.
Hickey, J. R., & Sidle, J. G. (2006). USDA Forest Service Office of International Programs trip report: Mission to support landscape planning in the Maringa-Lopori-Wamba landscape, Democratic Republic of Congo. Available at: (http://carpe.umd.edu/resources/Documents/MLWTripReportFinal.pdf)
Hohmann, G., & Fruth, B. (2008). New records on prey capture and meat eating by bonobos at Lui Kotale, Salonga National Park, Democratic Republic of Congo. Folia Primatologica, 79, 103–110.
Horn, A. D. (1980). Some observations on the ecology of the bonobo chimpanzee (Pan paniscus, Schwarz 1929) near Lake Tumba, Zaire. Folia Primatologica, 34, 145–169.
Hosmer, D., & Lemeshow, S. (1989). Applied logistic regression. New York: Wiley.
Inogwabini, B., Bewa, M., Longwango, M., Abokome, M., & Vuvu, M. (2008). The bonobos of the Lake Tumba—Lake Maindombe hinterland: Threats and opportunities for population conservation. In T. Furuichi & J. Thompson (Eds.), The bonobos: Behavior, ecology, and conservation (pp. 273–290). New York: Springer.
Jenks, G. F. (1967). The data model concept in statistical mapping. International Yearbook of Cartography, 7, 186–190.
Kano, T. (1984). Distribution of pygmy chimpanzees (Pan paniscus) in the Central Zaire Basin. Folia Primatologica, 43, 36–52.
Kano, T. (1992). The last ape: Behavior and ecology of pygmy chimpanzees. Stanford: Stanford University Press.
Kano, T., & Mulavwa, M. (1984). Feeding ecology of the pygmy chimpanzees (Pan paniscus) of Wamba. In R. L. Susman (Ed.), The pygmy chimpanzee: Evolutionary biology and behavior (pp. 233–274). New York: Plenum.
Kearns, M., & Ron, D. (1999). Algorithmic stability and sanity-check bounds for leave-one-out cross-validation. Neural Computation, 11, 1427–1453.
Kearns, M. J., Mansour, Y., Ng, A., & Ron, D. (1997). An experimental and theoretical comparison of model selection methods. Machine Learning, 27, 7–50.
Lehner, B., Verdin, K., & Jarvis, A. (2006). HydroSHEDS Technical Documentation. Washington, DC: World Wildlife Fund US. Available at: http://hydrosheds.cr.usgs.gov.
Li, H., & Reynolds, J. F. (1993). A new contagion index to quantify spatial patterns in landscapes. Landscape Ecology, 8, 155–162.
Li, H., Franklin, J. F., Swanson, F. J., & Spies, T. A. (1993). Developing alternative forest cutting patterns: a simulation approach. Landscape Ecology, 8, 63–75.
Luetzelschwab, J. (2007). Technical assistance to the African Wildlife Foundation on Planning and Zoning in the Maringa-Lopori-Wamba landscape, Congo. Final Trip Report. U.S. Forest Service. Available at: http://carpe.umd.edu/resources/Documents/USFS_MLWZoningReport.pdf
McGarigal, K., & Marks, B. J. (1995). FRAGSTATS: Spatial pattern analysis program for quantifying landscape structure. General Technical Report PNW-GTR-351. Portland, OR: U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station.
McGarigal, K., Cushman, S. A., Neel, M. C., & Ene, E. (2002). FRAGSTATS: Spatial pattern analysis program for categorical maps. Computer software program produced by the authors at the University of Massachusetts, Amherst. Available at: www.umass.edu/landeco/research/fragstats/fragstats.html
Mohneke, M., & Fruth, B. (2008). Bonobo (Pan paniscus) density estimation in the SW-Salonga National Park, Democratic Republic of Congo: Common methodology revisited. In T. Furuichi & J. Thompson (Eds.), The bonobos: Behavior, ecology, and conservation (pp. 151–166). New York: Springer.
Mulavwa, M. N., Yangozene, K., Yamba-Yamba, M., Motema-Salo, B., Mwanza, N. N., & Furuichi, T. (2010). Nest groups of wild bonobos at Wamba: selection of vegetation and tree species and relationships between nest group size and party size. American Journal of Primatology, 72, 575–586.
Neel, M. C., McGarigal, K., & Cushman, S. (2004). Behavior of class-level landscape metrics across gradients of class aggregation and area. Landscape Ecology, 19, 435–455.
Neter, J., Wasserman, W., & Kutner, M. H. (1989). Applied linear statistical models (2nd ed.). Homewood: Irwin.
Nishida, T. (1972). Preliminary information of the pygmy chimpanzees (Pan paniscus) of the Congo Basin. Primates, 13, 415–425.
Oates, J. F. (1994). Africa's primates in 1992: Conservation issues and options. American Journal of Primatology, 34, 61–71.
OSFAC (Observatoire Satellital des forêts d’Afrique central). (2010). Forêts d'Afrique centrale évaluées par télédétection (FACET): Forest cover and forest cover loss in the Democratic Republic of Congo from 2000 to 2010. South Dakota State University, Brookings, and University of Maryland, College Park.
Reinartz, G. E., Isia, I. B., Ngamankosi, M., & Wema, L. W. (2006). Effects of forest type and human presence on bonobo (Pan paniscus) density in the Salonga National Park. International Journal of Primatology, 27, 603–634.
Reinartz, G. E., Guislain, P., Mboyo Bolinga, T. D., Isomana, E., Inogwabini, B., Bokomo, N., Ngamankosi, M., & Wema Wema, L. (2008). Ecological factors influencing bonobo density and distribution in the Salonga National Park: Applications for Population Assessment. In T. Furuichi & J. Thompson (Eds.), The bonobos: Behavior, ecology, and conservation (pp. 167–188). New York: Springer.
Schumaker, N. H. (1996). Using landscape indices to predict habitat connectivity. Ecology, 77, 1210–1225.
Susman, R. L. (1984). The pygmy chimpanzee: Its evolutionary biology and behavior. New York: Plenum.
Thompson, J. (1997). The history, taxonomy, and ecology of the bonobo (Pan paniscus, Schwarz, 1929) with a first description of a wild populations living in a forest/savanna mosaic habitat. Doctoral dissertation, University of Oxford, Oxford.
Uehara, S. (1990). Utilization patterns of a marsh grassland within the tropical rain forest by the bonobos (Pan paniscus) of Yalosidi, Republic of Zaire. Primates, 31, 311–322.
UMass. (2000). UMass Landscape Ecology Lab, University of Massachusetts Amherst, MA. Available at: http://www.umass.edu/landeco/research/fragstats/documents/Metrics/Connectivity%20Metrics/Metrics/C121%20–%20COHESION.htm
USAID. (2010). Democratic Republic of Congo: Biodiversity and tropical forestry assessment (118/119) Final Report. Prosperity, livelihoods and conserving ecosystems indefinite quantity contract (PLACE IQC) Contract no. EPP-I-03-06-00021-00. Available at: http://pdf.usaid.gov/pdf_docs/PNADS946.pdf
USFS. (2004). Final environmental impact statement for Chippewa and superior national forests. Chapter 3.3.2: Spatial patterns management indicator habitats. Available at: http://www.fs.fed.us/r9/chippewa/plan/final/feis/Final_EIS/Final_EIS_Chapter_3/Final_EIS_3_3_2_Wildlife_Spatial_Patterns.pdf
Van Krunkelsven, E. (2001). Density estimation of bonobos (Pan paniscus) in Salonga National Park, Congo. Biological Conservation, 99, 387–391.
White, F. (1996). Pan paniscus 1973 to 1996: twenty-three years of field research. Evolutionary Anthropology: Issues, News, and Reviews, 5, 11–17.
Wilkie, D. S., Sidle, J. G., & Boundzanga, G. C. (1992). Mechanized logging, market hunting and a bank loan in Congo. Conservation Biology, 6, 570–580.
Williams, B. K., Nichols, J. D., & Conroy, M. J. (2002). Analysis and management of animal populations. San Diego: Academic.
Acknowledgments
We thank Pasteur C. W. Balongelwa, G. T. Muda, Principal Conservator Nayifilua, and F. Botamba for coordinating our work with their respective agencies; the Congolese Institute for the Conservation of Nature (ICCN); the Congolese Ministry of Scientific Research; the Lomako-Yokokala Faunal Reserve; and the African Wildlife Foundation (AWF). R. Mujinga, the Chief Administrator of the Territory of Bongandanga, facilitated community acceptance of our presence and generously provided secure housing. We warmly thank G. Reinartz, S. McLaughlin, P. Guislain, and N. Etienne of the Zoological Society of Milwaukee. We greatly appreciate the hospitality of T. Hart and TL2. We thank T. Eppley, S. Iloko-Nsonge, S. Boongo, and J. Lokuli-Lokuli, who assisted with field work. Funding for field research was provided by the U.S. Fish and Wildlife Service – Great Ape Conservation Fund (98210-8-G654) to J. Carroll, N. Nibbelink, and J. Hickey; a research fellowship from the Wildlife Conservation Society (WCS) to J. Hickey; and a small grant from Global Forest Science to N. Nibbelink, J. Carroll, and J. Hickey. Ideawild provided essential field gear. J. Hickey holds an American Fellowship from the American Association of University Women (AAUW). AWF and WCS provided logistical support in DRC, notably J. Dupain, T. Senga, and R. K. Tshombe. We thank K. Barrett, D. Elkins, L. Worsham, and three anonymous reviewers for insightful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Hickey, J.R., Carroll, J.P. & Nibbelink, N.P. Applying Landscape Metrics to Characterize Potential Habitat of Bonobos (Pan paniscus) in the Maringa-Lopori-Wamba Landscape, Democratic Republic of Congo. Int J Primatol 33, 381–400 (2012). https://doi.org/10.1007/s10764-012-9581-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-012-9581-8