
Abstract
Captive reintroductions often suffer high mortality, with predation as one source. Many species learn about predators; thus training captive-born animals to recognize predators may increase survivorship. We adapted variants of methods developed for birds to attempt to condition monkeys to mob a predator. Captive-reared cotton-top tamarins (Saguinus oedipus) did not differentiate between a snake and a rat, confirming previous research that naïve cotton-top tamarins do not fear snakes. Tamarins then viewed a living snake during playback of mobbing calls. In posttests, tamarins did not mob the snake. We then eliminated the pretest to avoid potential habituation and placed a stillborn tamarin with the snake in an attempt to heighten arousal. In posttests, tamarins again did not mob the snake, although they did increase their rates of calls, indicating mild arousal. Overall, none of 8 groups of cotton-top tamarins learned to mob a predator. Potential reasons for failure include conditioning multiple subjects at once and the lack of an experienced demonstrator. Observing a demonstrator was not necessary for birds to acquire mobbing, but may be necessary for tamarins.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Reintroductions of captive-born animals aim to restore populations to the wild or to augment declining numbers (Olney et al. 1994). Typically, the greatest challenge for captive reintroductions is high mortality (Soorae and Baker 2002), with predation as one source (Beck et al. 1991; Soorae and Baker 2002). Some primates learn about their predators during development. In captivity, wild-born rhesus macaques (Macaca mulatta) showed fear of snakes whereas captive-born macaques did not (Mineka et al. 1980). However, captive-born individuals learned to fear snakes simply by watching a wild-born individual behave fearfully toward a snake (Cook et al. 1985; Mineka et al. 1984). There is suggestive evidence of a role of learning in 2 other species. Captive-born, predator naïve cotton-top tamarins (Saguinus oedipus) showed no difference in reaction to a snake vs. a rat, and the intensity of the response to the snake appeared lower than that in the wild (Hayes and Snowdon 1990). Wild vervets (Chlorocebus aethiops) appeared to learn during development to respond appropriately to different alarm calls by their groupmates (Cheney and Seyfarth 1990).
Many species learn to recognize predators. Captive European blackbirds (Turdus merula) learned to fear a novel, harmless bird by seeing and hearing a mobbing response directed at the stimulus (Curio et al. 1978). New Zealand robins (Petroica australis) and carib grackles (Quiscalus lugubris) learned to fear novel animals by hearing prerecorded alarm and mobbing calls played from a speaker while viewing the animal (Griffin and Galef 2005; Maloney and McLean 1995; McLean et al. 1999). Three species of wallaby (Aepyprymnus rufescens, Setonix brachyurus, and Macropus eugenii) have been conditioned to fear a stuffed fox (Vulpes vulpes; Griffin and Evans 2003; Griffin et al. 2001; McLean et al. 2000). Predator training in captivity has produced a survival advantage in 2 species: brook trout (Salvelinus fontinalis; Mirza and Chivers 2000) and black-tailed prairie dogs (Cynomys ludovicianus; Shier and Owings 2007).
Wild marmosets and tamarins mob snakes (Bartecki and Heymann 1987; Buchanan-Smith 1990; Corrêa and Coutinho 1997; Ferrari and Lopes Ferrari 1990; Shahuano Tello et al. 2002). Few observations described the mobbing response and vocalizations in detail, so we relied on a captive study of mobbing in cotton-top tamarins to model it. Mobbing consisted of rapid approaches to the predator, retreating and reapproaching, visual fixation, piloerection, and specific vocalizations used only during mobbing (Campbell and Snowdon 2007).
We adapted the procedure used to train New Zealand robins to fear a predator (Maloney and McLean 1995; McLean et al. 1999). Both monkeys and robins use vocalizations to warn groupmates about predators, and both learn socially (Fragaszy and Perry 2003). Like robins, cotton-top tamarins and related species form long-term pair bonds, defend territories, and cooperatively rear young. Cotton-top tamarins should be good candidates to use the auditory playback method. The ultimate value of this method is the potential to condition mobbing when no experienced, wild-born individuals are present in captivity, as is the case for many breeding programs.
If conditioning is successful, we predict subjects will display more mobbing behaviors and vocalizations to the snake after conditioning than before, and more to the snake than to control objects. Other behaviors indicative of arousal, but of a lower intensity than mobbing, should show the same pattern. Relaxed behaviors should be more numerous before conditioning than after when viewing the snake, and more numerous when viewing the controls than the snake.
Experiment 1
Methods
We tested 4 adult male-female pairs of cotton-top tamarins (Saguinus oedipus). All were born in captivity and spent their entire lives indoors. The tamarins had never seen any natural predators, and their only experiences with any nonhuman animal species were with the mealworms and crickets they occasionally received during feeding. All of the subjects were born after Hayes and Snowdon (1990), in which tamarins saw a snake. Pairs lived in 1.6 m × 0.93 m × 2.4 m (L × W × H) cages. We equipped all cages with natural branches and ropes as the main climbing structures, and we offered food and water at ca. 1 m above the floor to approximate the natural arboreal habitat of tamarins. The main noon feed consisted of commercial foods (Zu-Preem Marmoset Diet and Purina Marmoset Chow), fresh fruits and vegetables, and a starch. We provided supplemental protein-rich foods at lights-on and in the late afternoon. Water was available ad libitum. All subjects were on a 12-h light-dark cycle (Ginther et al. 2001).
The box used for stimulus presentation was of clear Plexiglas on 5 sides and opaque Plexiglas on the end (61 cm × 45.5 cm × 32 cm, L W x H). An opaque plastic cover sat in grooves over the clear box, which allowed us to slide the cover open and closed. The opaque cover added 0.5–1.5 cm to each side, front, and top. Using a hook an observer could pull back the opaque cover, exposing the box and the stimulus inside to the subjects, and then push the cover back on to end exposure. We used a Sennheiser directional microphone (Model ME 66), RCA speaker (Model 40-5000), two Marantz tape recorders (Model PMD 201), University Sound amplifier (Model 1800-40), observation blind, and 2 Dell laptop computers for recording data. The test box was placed inside the subjects’ cage on a cart, which elevated the bottom of the box to ca. 1 m. The microphone, tape recorder, and amplifier were outside of the cage. The speaker was hidden at the side of the cage behind a sheet to prevent the tamarins from seeing the speaker with minimal sound degradation. The blind faced the front of the cage and had room for 2 observers, both with computers.
The stimuli were a live Colombian rainbow boa constrictor (Epicrates cenchria maurus), a live white laboratory rat (Rattus norvegicus), and a multicolored plastic toy (ca. 25 cm × 10 cm × 10 cm, L × W × H). The boa was a fully-grown adult male about 1.6 m long. This subspecies ranges from Costa Rica to northern Colombia and Venezuela, whereas the species in general extends south through much of South America. Cotton-top tamarins are endemic to northwest Colombia, so both species likely overlap in range. No reports exist of rainbow boas preying on callitrichids, but they could pose a threat to tamarins, especially infants and juveniles. Saddle-back tamarins (Saguinis fuscicollis nigrifrons) mobbed 2 tree boas (Corallus enydris), a species similar in size to rainbow boas (Bartecki and Heymann 1987); a common boa (Boa constrictor) of ca. 1.5 m attacked a moustached tamarin (S. mystax; Shahuano Tello et al. 2002); and buffy tufted-ear marmosets (Callithrix aurita), mobbed a 1.0–1.2 m long viper (Bothrops jararaca; Corrêa and Coutinho 1997). Therefore, snakes in the size range of this boa are a threat to wild callitrichids.
There were 4 phases: habituation, pretest, conditioning, and posttest. We conducted 3 habituation sessions exactly like experimental sessions except that there was no stimulus in the box. We opened and closed the box during these sessions to habituate subjects to the movement. We considered the tamarins habituated when they stopped displaying arousal or anxiety to the moving lid. All subjects habituated rapidly and showed no signs of needing additional sessions. The pretest phase established baseline rates of response to each of the 3 stimuli. During the conditioning phase, we played prerecorded cotton-top tamarin mobbing calls while exposing subjects to the snake on 3 separate days. During the posttests, we exposed subjects to each stimulus twice.
We conducted only 1 session per day, with 1–2 d between sessions. All subjects encountered the toy in the first pretest, the snake second, and the rat third. To control for the novelty of seeing an object inside the box, we presented the toy first. We presented the snake second so that there would be ≥1 session between the pretest exposure to the snake and the conditioning sessions. After 3 snake conditioning sessions, the first posttest was with the snake, to maximize any effect of learning. We next counterbalanced the order of the rat and toy for posttest sessions 2 and 3; we then repeated the same sequence for the second set of posttests.
Campbell and Snowdon (2007) recorded and described mobbing calls. The calls that served as stimuli in the current study came from 1 family in our colony that did not participate in the experiment. During conditioning, we played back the mobbing calls at a volume that matched the intensity of the vocalizations as measured by a sound pressure meter during an actual mobbing response (90–100 dB).
After we set up the equipment, 2 observers recorded preexposure data for 10 min. Then we removed the cover and the subjects could view the stimulus for 3 min. After exposure, we replaced the cover and took 10 min of postexposure data.
Each observer sampled 1 member of a pair as a focal individual via The Observer 4.1 (Noldus Information Technology). Observers recorded the time spent looking at the stimulus or covered box during the preexposure and postexposure and recorded behavior that indicated mobbing, including piloerection and rapid lunges at the stimulus. They also recorded relaxed behavior, including frequency of eating, drinking, and mating; and time spent allogrooming, huddling, and in contact with mate. Tamarins did not contact mates when anxious, but only when they were in a calm state. To measure interobserver reliability during pilot tests, both observers focal sampled the same individual.
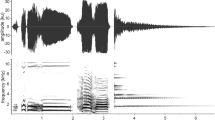
We tape recorded vocalizations, which were filtered (1-kHz high pass, 14-kHz low pass), digitized into .WAV files, and visualized via sonogram using Avisoft SASLab Pro (sampling frequency = 44 kHz, FFT-length = 512, Hamming window, bandwidth = 81 Hz, resolution = 62 Hz; temporal: 1/bandwidth = 12.3 ms, resolution = 4 ms). We identified calls by comparing them to catalogs of cotton-top tamarin vocalizations (Campbell and Snowdon 2007; Cleveland and Snowdon 1982). We tallied the vocalizations associated with fear, mobbing, or antipredator behavior in general, which included barks; growls; high-chirp trills; H-chirp trills; chevron chatters; chirp bursts; and A, E, and H chirps. We sorted calls into groups based on the criteria of Campbell and Snowdon (2007). We sorted barks and growls into the “mob” class as the highest intensity vocalizations; we sorted high-chirp trills, H-chirp trills, chevron chatters, and chirp bursts into the pulsed class as intermediate intensity; and A, E, and H chirps comprised the chirp class as the lowest intensity calls. Thus we have 3 classes of calls for analysis: mob, pulsed, and chirp, in descending intensity.
For each group we compared pretest with posttest behavior to determine if conditioning was successful. We looked at 2 levels of analysis: First, did conditioning induce any tamarins to display mobbing, the most adaptive response to a large snake? Second, were there changes in any behavioral measure, indicating some behavioral change short of a full mobbing response? Each pair served as its own control, and we evaluated each pair as to whether it learned to mob the snake or not.
The pair is the appropriate unit of analysis because we cannot distinguish between individual recognition of the snake as a predator vs. emotional contagion to the mate’s response. This would require testing subjects alone, to which the tamarins were not habituated. The criteria for a learned mobbing response were the highest intensity mobbing behaviors in the species: piloerection, lunging, and mob calls (Campbell and Snowdon 2007). The statistical assessment of responses across pairs used measures of attention, relaxed behaviors, and pulsed and chirp calls to indicate how anxious the monkeys were in the presence of the various stimuli. The measures do not indicate mobbing, but instead the degree of arousal. The sample size for both assessments is 4.
Results
Interobserver reliability scores were always in the 85–99% range (both Index of Concordance and Cohen’s kappa).
Subjects gave no piloerections, lunges, or mob calls to the snake or any other stimulus during the pretest. The overall rate of calling is in Fig. 1. The tamarins spent similar amounts of time looking at the snake (51.3% ± 9.1% of exposure) and the rat (48.2% ± 5.3%), but they spent significantly more time looking at both the snake and rat than at the toy (31.2% ± 3.9%, 2-tailed paired t-tests, t 3 = 5.57, p = 0.011, d = 2.30 snake vs. toy; t 3 = 6.01, p = 0.009, d = 2.79 rat vs. toy). We never observed comfort behavior during exposure to any stimulus. The tamarins displayed equal frequencies (2.8 ± 2.5 pretest vs. 6.8 ± 5.7 posttest, 2-tailed paired t-test, t 3 = 2.38, p = 0.10, d = 1.19) and durations (1.4 ± 1.3 s pretest vs. 22.1 ± 24.5 s posttest, 2-tailed paired t-test, t 3 = 1.74, p = 0.18, d = 0.87) of comfort behavior before and after viewing the snake. Friedman tests comparing frequencies and durations of comfort behavior during postexposure across all 3 stimuli are nonsignificant.

Mean rate of calling + SD during pretest exposure to the stimulus in Experiment 1. The difference in the rate of chirps is not significant (paired t-tests).
During conditioning, subjects consistently piloerected on hearing the mob calls on the audiotape, but we observed no lunges at the snake. We could not count vocalizations during conditioning sessions owing to the overlap between calls from the subjects and calls from the playback tape. However, we observed no mob calls from any tamarin during conditioning sessions. The general rate of vocalizing was high, but all of the vocalizations belonged to the pulsed and chirp calls. The mean percentage of time looking at the stimulus during conditioning was 31.3% (SD = 14.4%). We observed no comfort behavior during conditioning, but frequencies (t 3 < 1.0, NS) and durations (t 3 < 1.0, NS) of comfort behaviors were similar in pre- and postexposure.
In posttests, no individual piloerected or lunged at the snake, and we recorded no calls from the mob class from any group. We expected tamarins to produce more pulsed and chirp calls to the snake in the posttest than the snake in the pretest or the rat in the posttest, but this did not occur. Looking times between the snake and rat during posttest exposures do not differ significantly (t 3 < 1.0, NS). Comfort behavior was similar before and after viewing the snake for frequencies (t 3 < 1.0, NS) and durations (t 3 < 1.0, NS). A Friedman test among all 3 stimuli during postexposure showed a trend for counts of comfort behaviors to be ranked in order of rat (2.9) > toy (1.9) > snake (1.3, χ2 2 = 5.73, p = 0.0569). Two-tailed paired t-tests comparing frequencies (6.8 ± 5.7 before conditioning vs. 2.8 ± 1.8 after conditioning, t 3 = 1.26, p = 0.30, d = 0.70) and durations (22.1 ± 24.5 s before conditioning vs. 2.6 ± 3.5 s after conditioning, t 3 = 1.42, p = 0.25, d = 0.91) of comfort behavior during postexposure to the snake before and after conditioning are nonsignificant.
Discussion
The pretest response to the snake indicated that the captive-born, predator-naive tamarins did not recognize the snake as a predator. The response of wild callitrichids to snakes is to mob, and there was no behavior consistent with a mobbing response. Further, the response to the snake did not differ from the response to the rat. These results confirm those of Hayes and Snowdon (1990). Therefore, any mobbing behavior observed in the posttests would be due to conditioning.
However, there was no conditioned mobbing of the snake. There were no piloerections, lunges, or mob calls to the snake in the posttest. This method failed to condition a mobbing response. However, the tamarins spent relatively little time looking at the snake during conditioning. Thus, their attention may not have been sufficiently attracted to the snake during conditioning. The pretest exposure to the snake may have immunized the subjects against successful learning. Because the subjects showed no signs of mobbing during the pretest, we removed the pretests in Experiment 2 and attempted to attract attention to the snake during conditioning.
Experiment 2
Methods
We tested 3 male-female pairs of cotton-top tamarins and 1 family of 6 individuals. Husbandry details were the same as for Experiment 1 except that the family was housed in a cage measuring 3.0 × 2.3 × 1.8 m. The apparatus and stimuli were identical to those in Experiment 1.
We eliminated the pretest phase to prevent possible latent inhibition against successful learning during the initial, consequence-free exposure to the snake, so subjects went directly from habituation to conditioning. During the conditioning phase, we placed a dead, frozen infant tamarin that had died of natural causes in the box along with the snake to attract the subjects’ attention. Seeing an infant lying alone, i.e., not carried, was arousing for many tamarins. We thought that the presence of an infant with the snake might simulate a situation the monkeys would perceive as dangerous. We manipulated other variables during pilot testing, but eliminating the pretest and adding the stillborn tamarin showed the most promise. All other aspects of the design and procedure were identical.
Data recording and analysis was the same, except that we could not gather data on looking time and comfort behavior for 1 pair and the family because of logistical constraints. Owing to the small sample size (2), we removed these 2 measures from the analysis. We recorded vocalizations, piloerections, and lunges for all 4 groups. Owing to additional constraints, the family group had only 1 posttest session with the snake and rat, and no sessions with the toy. We analyzed data as for Experiment 1.
Results
All subjects in Experiment 2 piloerected when vocal playback began. During conditioning, we recorded no mob calls and observed no lunges toward the snake. The rate of vocalizing during conditioning was high, but all vocalizations came from the lower intensity pulsed and chirp classes.
In the posttests, no individual piloerected or lunged at the snake. We recorded no calls from the mob class from any group, and only 1 group gave >1 pulsed call to the snake. Subjects gave significantly more chirp calls to the snake than the rat during the posttest (Fig. 2; log10 transformed: 1.5 ± 0.5 snake vs. 0.4 ± 0.5 rat, 2-tailed paired t-test, t 3 = 4.33, p = 0.023, d = 2.19).

Back-transformed rate of calling for a pair of tamarins during posttest exposure in Experiment 2. Values are averaged for groups with 2 posttests to each stimulus (except P, which only had one post-test). We corrected group P for its larger size (N = 6) by dividing its rate by 3, which gives the rate of calling for 2 tamarins. We used a log10 transformation to normalize a skewed distribution.
In combined results from both experiments, none of 8 groups displayed mobbing behaviors or vocalizations toward the snake after conditioning. With pilot subjects included, none of 15 groups or pairs showed mobbing toward the snake after conditioning.
Discussion
There was no conditioned mobbing of the snake. We never observed piloerections, lunges, or mob calls in posttests. Thus, the second method also failed to condition a mobbing response. All of the groups gave more low-intensity chirp calls to the snake than the rat in the posttest, suggesting a possible increase in vigilance. This is unlikely the result of pseudoconditioning, i.e., the subjects having more exposures to the snake than the rat or toy, because the groups in Experiment 1 and pilot testing also had more exposure to the snake than to the rat or toy, but none of the groups showed an increase in chirp calls toward the snake.
General Discussion
The tamarins in our studies failed to show conditioned mobbing behavior. In all, we tested 15 pairs or groups of tamarins with modifications of methods that have successfully induced mobbing in other species. We did not observe a single mobbing behavior in posttests (mobbing vocalizations, lunges, piloerections), making it unlikely that further variations on the method with more groups of tamarins would produce different results. Future studies should explore qualitatively different methods of conditioning instead of continuing with the auditory playback method.
Several factors may have prevented the subjects from learning to mob the snake. The most important cues missing from conditioning were visual cues. Auditory playback alone was sufficient to condition mobbing in New Zealand robins (Maloney and McLean 1995; McLean et al. 1999), but it was not sufficient for tamarins in this experiment. Cotton-top tamarins, and perhaps other primates, may learn during mobbing both by hearing vocalizations and by seeing groupmates direct aggressive behaviors toward a novel stimulus. The successful conditioning of snake-fear in rhesus macaques involved both watching and hearing a knowledgeable macaque behave fearfully toward the snake (Cook et al. 1985; Mineka et al. 1984).
A second problem may have been attempting to condition 2 subjects together. As attention to the snake was generally low and mobbing behaviors were never directed at the snake, this may have reinforced to the mate that the snake was not the object to mob. Individual conditioning might produce different results. Conversely, the most responsive group we tested was the family, and adults may be more responsive with offspring present. In contrast to the prior conclusion, having more individuals present, especially dependent young, may fuel a greater response. Rhesus macaques were conditioned with one naïve individual watching a demonstrator (Cook et al. 1985; Mineka et al. 1984). Birds have learned to mob a novel stimulus with a naïve individual present (Curio et al. 1978) or a naïve pair present with no demonstrator (Maloney and McLean 1995; McLean et al. 1999). Conditioning multiple subjects together was not a problem for New Zealand robins, but it may be problematic for primates if they indeed are more sensitive to visual displays of mobbing.
Another possibility for the lack of conditioning is that the playback fidelity may have been insufficient to stimulate all of the appropriate behaviors, and calls of strange tamarins may have confused the subjects. Because all of the tamarins piloerected and became very aroused at the onset of playback of mobbing calls, problems with fidelity appear to be ruled out. Pseudoreplication (having only 1 example of a predator and 1 stimulus set) should not be a problem, as every previous study of predator learning had these same issues in their designs.
Because the snake was inside a clear plastic box, the situation may not have been perceived as dangerous enough to be stimulating mobbing calls. It seems unlikely that the subjects would know that the snake could not get out of the box because their own plastic nest boxes have an opening. Finally, snakes, being ambush predators, are not highly active. The snake had plenty of room to move around and rear up (tightly coiled, the snake would occupy less than a quarter of the surface area of the box), and his activity varied from exploring the box to being mostly still. A more active predator may have elicited more attention from the tamarins, and hence improved learning, but this would not be the whole story. Tamarins need to learn to recognize snakes in the wild, and this is especially important for immobile snakes, as that is their hunting strategy. An immobile snake is a dangerous snake. Acquiring fear of snakes was not a problem for rhesus macaques (Cook et al. 1985; Mineka et al. 1984).
Our results confirm other results that captive-born, naïve primates do not innately recognize snakes as predators (Hayes and Snowdon 1990; Mineka et al. 1980), although our results conflict with conclusions drawn by other researchers (Barros et al. 2002; Castro 1990). We have demonstrated the difficulty of conditioning a mobbing response in captive-reared cotton-top tamarins. Simply pairing mobbing calls and a novel, alive, and moving predator was not sufficient to condition a mobbing response. The combination of a lack of innate predator recognition and the difficulty in conditioning an appropriate mobbing response (in the absence of a knowledgeable demonstrator) casts doubt on the ability of captive-reared callitrichids to cope with terrestrial predators in the wild.
The difference between our results and those for rhesus macaques reinforces the likelihood that cues from a demonstrator may be necessary for primates to acquire predator recognition. The most important factors to include in future studies seem to be either a demonstrator or conditioning subjects when alone if using a social learning paradigm. Methods that utilize individual training and do not try to model social learning may also be successful (Griffin et al. 2001; McLean et al. 2000).
References
Barros, M., Boere, V., Mello, E. L. J., & Tomaz, C. (2002). Reactions to potential predators in captive-born marmosets (Callithrix penicillata). International Journal of Primatology, 23, 443–454. doi:10.1023/A:1013899931878.
Bartecki, U., & Heymann, E. W. (1987). Field observations of snake-mobbing in a group of saddle-back tamarins, Saguinus fuscicollis nigrifrons. Folia Primatologica, 48, 199–202. doi:10.1159/000156296.
Beck, B. B., Kleiman, D. G., Dietz, J. M., Castro, M. I., Carvalho, C., Martins, A., et al. (1991). Losses and reproduction in reintroduced golden lion tamarins, Leontopithecus rosalia. Dodo. Journal of the Jersey Wildlife Preservation Trust, 27, 50–61.
Buchanan-Smith, H. M. (1990). Polyspecific association of two tamarin species, Saguinus labiatus and Saguinus fuscicollis, in Bolivia. American Journal of Primatology, 22, 205–214. doi:10.1002/ajp.1350220306.
Campbell, M. W., & Snowdon, C. T. (2007). Vocal response of Saguinus oedipus during mobbing. International Journal of Primatology, 28, 257–270. doi:10.1007/s10764-007-9123-y.
Castro, M. I. (1990). A Comparative Study of the Anti-predator Behavior in the Three Species of Lion Tamarins (Leontopithecus) in Captivity. Master's thesis, University of Maryland, College Park.
Cheney, D. L., & Seyfarth, R. M. (1990). How monkeys see the world. Chicago: University of Chicago Press.
Cleveland, J., & Snowdon, C. T. (1982). The complex vocal repertoire of the adult cotton-top tamarin (Saguinus oedipus oedipus). Zeitschrift fur Tierpsychologie, 58, 231–270.
Cook, M., Mineka, S., Wolkenstein, B., & Laitsch, K. (1985). Observational conditioning of snake fear in unrelated rhesus monkeys. Journal of Abnormal Psychology, 94, 591–610. doi:10.1037/0021-843X.94.4.591.
Corrêa, H. K. M., & Coutinho, P. E. G. (1997). Fatal attack of a pit viper, Bothrops jararaca, on an infant buffy-tufted ear marmoset (Callithrix aurita). Primates, 38, 215–217. doi:10.1007/BF02382010.
Curio, E., Ernst, U., & Vieth, W. (1978). Cultural transmission of enemy recognition: one function of mobbing. Science, 202, 899–901. doi:10.1126/science.202.4370.899.
Ferrari, S. F., & Lopes Ferrari, M. A. (1990). Predator avoidance behaviour in the buffy-headed marmoset, Callithrix flaviceps. Primates, 31, 323–338. doi:10.1007/BF02381104.
Fragaszy, D. M., & Perry, S. (2003). The biology of traditions. Cambridge, UK: Cambridge University Press.
Ginther, A. J., Ziegler, T. E., & Snowdon, C. T. (2001). Reproductive biology of captive male cottontop tamarin monkeys as a function of social environment. Animal Behaviour, 61, 65–78. doi:10.1006/anbe.2000.1587.
Griffin, A. S., & Evans, C. S. (2003). Social learning of antipredator behaviour in a marsupial. Animal Behaviour, 66, 485–492. doi:10.1006/anbe.2003.2207.
Griffin, A. S., & Galef, B. G. (2005). Social learning about predators: does timing matter? Animal Behaviour, 69, 669–678. doi:10.1016/j.anbehav.2004.05.020.
Griffin, A. S., Evans, C. S., & Blumstein, D. T. (2001). Learning specificity in acquired predator recognition. Animal Behaviour, 62, 577–589. doi:10.1006/anbe.2001.1781.
Hayes, S. L., & Snowdon, C. T. (1990). Predator recognition in cotton-top tamarins (Saguinus oedipus). American Journal of Primatology, 20, 283–291. doi:10.1002/ajp.1350200404.
Maloney, R. F., & McLean, I. G. (1995). Historical and experimental learned predator recognition in free-living New Zealand robins. Animal Behaviour, 50, 1193–1201. doi:10.1016/0003-3472(95)80036-0.
McLean, I. G., Hölzer, C., & Studholme, B. J. S. (1999). Teaching predator-recognition to a naïve bird: Implications for management. Biological Conservation, 87, 123–130. doi:10.1016/S0006-3207(98)00024-X.
McLean, I. G., Schmitt, N. T., Jarman, P. J., Duncan, C., & Wynne, C. D. L. (2000). Learning for life: training marsupials to recognise introduced predators. Behaviour, 137, 1361–1376. doi:10.1163/156853900501971.
Mineka, S., Keir, R., & Price, V. (1980). Fear of snakes in wild- and laboratory-reared rhesus monkeys (Macaca mulatta). Animal Learning & Behavior, 8, 653–663.
Mineka, S., Davidson, M., Cook, M., & Keir, R. (1984). Observational conditioning of snake fear in rhesus monkeys. Journal of Abnormal Psychology, 93, 355–372. doi:10.1037/0021-843X.93.4.355.
Mirza, R. S., & Chivers, D. P. (2000). Predator-recognition training enhances survival of brook trout: evidence from laboratory and field-enclosure studies. Canadian Journal of Zoology, 78, 2198–2208. doi:10.1139/cjz-78-12-2198.
Olney, P. J. S., Mace, G. M., & Feistner, A. T. C. (1994). Creative conservation. London: Chapman and Hall.
Shahuano Tello, N., Huck, M., & Heymann, E. W. (2002). Boa constrictor attack and successful group defence in moustached tamarins, Saguinus mystax. Folia Primatologica, 73, 146–148. doi:10.1159/000064795.
Shier, D. M., & Owings, D. H. (2007). Effects of social learning on predator training and postrelease survival in juvenile black-tailed prairie dogs, Cynomys ludovicianus. Animal Behaviour, 73, 567–577. doi:10.1016/j.anbehav.2006.09.009.
Soorae, P. S., & Baker, L. R. (2002). Re-introduction NEWS: Special primate issue, newsletter of the IUCN/SSC re-introduction specialist group. 21:60. Abu Dhabi, UAE.
Acknowledgments
We thank Julie A. Borowski, Stella M. Joyce, and Kate F. Washabaugh for assistance with data collection, and Andrew Mulder for construction of the experimental box and the blind. Liza R. Moscovice provided comments on an early version of this manuscript. The comments of 2 anonymous reviewers significantly improved this manuscript. The American Psychological Association, the Animal Behavior Society, the Department of Psychology at the University of Wisconsin-Madison, and the National Institute of Mental Health grant MH029775 provided funding for this research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Campbell, M.W., Snowdon, C.T. Can Auditory Playback Condition Predator Mobbing in Captive-reared Saguinus oedipus?. Int J Primatol 30, 93–102 (2009). https://doi.org/10.1007/s10764-008-9331-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-008-9331-0