
Abstract
Black-shanked douc langurs (Pygathrix nigripes) are Southeast Asian colobines about which primatologists know very little, but they are classed as endangered because of population decline due to habitat loss. Two preliminary studies have shown that this monkey is primarily folivorous, but there are few details of plant selection or seasonal changes in diet. We set out to observe douc langurs directly in the wild to quantify the diet during wet and dry seasons. We confirmed that the species relies on foliage throughout the year, but it includes significant proportions of fruit and flowers in its diet when those items are available. The douc langurs selected various parts from 152 species of plants in 2 national parks, but there did not appear to be favorite species that were heavily selected over others. In both the wet and dry seasons, the black-shanked douc langurs ate mostly leaves, but the diversity of plants consumed increased and the proportion of fruit eaten almost doubled in the wet season. There were diurnal shifts in food selection, but we found no evidence that the douc langurs were foraging strategically to maximize their protein or energy intake.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Black-shanked douc langurs (Pygathrix nigripes) are large (Napier and Napier 1985) colobines endemic to Vietnam and Cambodia (Hoang 2007). Primatologists know little of their ecology or demographics, but they are classified as endangered (IUCN 2006). Habitat fragmentation and loss and hunting are likely the major threats to the species, but there is also a live trade in primates in the region (Nadler et al. 2003).
Human population growth will probably continue to place pressure on Pygathrix nigripes. Vietnam has a human population of >85 million, which is estimated to be growing at 1.4% annually (National Environment Agency 2005). Before 1997, natural forest in Vietnam was reduced by 100,000–200,000 ha/yr (Wege et al. 1999). Since 1997, this rate of loss has slowed to ca. 10,000 ha/yr (Wege et al. 1999). Since 1995, >1.2 million people have migrated to the central highlands, which is a stronghold of the black-shanked douc langur (Hoang 2007; National Environment Agency 2005). Forest reduction and population growth have placed severe pressure on the forests and biodiversity of the area (Lippold and Vu Ngoc Thanh 1998). These circumstances suggest an urgent need to address the lack of knowledge about an obligate forest dweller such as the black-shanked douc langur so that sound conservation planning can begin.
Feeding is one of the most basic aspects of an animal's ecology. Conservation measures such as quantifying suitable habitat, choosing areas for protection or species to be planted for remediation of degraded habitat are possible only if the food plants are known. Obtaining this information is especially complicated for primates because there are often large changes in diet through the year, or over their geographical range (Barnett 1995).
There are only 2 reports on the diet of black-shanked douc langurs. A preliminary report (Hoang unpubl. data) from a study in Vietnam indicated that douc langurs ate leaves of 27 plant species, fruit of 24 species, and flowers of 10 species. A study in Cambodia (Rawson 2006) reported that the diet of douc langurs consisted of 40% leaves, 40% seeds, 11% fruits, and 9% flowers by frequency. The dentition of colobines (Lucas and Teaford 1994) and studies of other Pygathrix spp. (Lippold 1998, 1977; Pham Nhat 1993; Pham Nhat et al. 2000) suggest that black-shanked douc langurs are most likely to be folivorous, but the importance of fruit and seeds in the diet is unknown. Similarly, researchers have not yet demonstrated any seasonal or regional shifts in diet.
We report the largest and most comprehensive study of the diet of wild, free-ranging black-shanked douc langurs. Specifically, we aimed to expand the list of plant food species, examine the selection of plant parts, and quantify seasonal changes in diet.
Methods
Study Sites
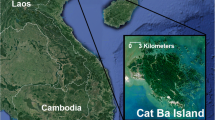
We conducted this research in Nui Chua and Phuoc Binh National Parks (Fig. 1). Both protected areas are in the Greater Amamites Ecoregion, which is one of the most diverse monsoon forests in Asia (Baltzer et al. 2001). We chose these parks because they both have known populations of black-shanked douc langurs and allowed data to be collected in a range of altitude, climatic, and forest types.

Location of study areas.
Phuoc Binh lies between 11°58′–12°10′N and 108°43′–108°49′E, covers 19,814 ha, and is in the center of black-shanked douc langurs' range. It has an annual rainfall of >2000 mm. The mean maximum and minimum temperatures are 38.8°C and 14.2°C, respectively (Sub Institute of Forest Inventory and Planning 2000); hence the park has a wet and humid climate, which allows the 3000 human inhabitants to cultivate wet rice and practice shifting cultivation in the park's buffer zone (Phuoc Binh Commune People's Committee 2005).
The complicated topography of the park is the reason that the forest has been maintained almost intact despite the fact that the area was highly impacted during the Vietnam War (Sub Institute of Forest Inventory and Planning 2002). However, recently the forest seems to be subject to increasing human disturbance, especially hunting pressure (pers. obs.). There are 6 main vegetation types in the park (Sub Institute of Forest Inventory and Planning (2002; Table I).
Nui Chua lies between 11°39′–11°48′N and 109°04′–109°14′E and covers 22,513 ha, with 7532 ha being buffer zone in which there are 14 villages or communes supporting >24,000 people (Nui Chua National Park 2005). The park is located in the driest part of Vietnam and receives an annual average rainfall of 697 mm (Sub Institute of Forest Inventory and Planning 1996). The climate is hot and dry, and only 6 of the park's many streams run in the dry season.
The original vegetation of the park was a mixture of evergreen forest, semideciduous forest, and deciduous forest. However, most of the forest was destroyed by overexploitation in the early 1990s (BirdLife International and Forest Inventory and Planning Institute 2001). The only primary forest that remains is evergreen forest at elevations >700 m asl in the north of the park. Below 700 m asl there are large areas of 4 types of secondary forest (Table II).
Feeding Records
We systematically walked across both national parks, collecting data wherever we located an individual or group of black-shanked douc langurs. Where the vegetation and landform allowed, we walked along straight-line transects to cover the areal extent of the park. In other places, e.g., thorny scrub in Nui Chua, we were restricted to existing tracks through the dense vegetation.
When we encountered a group of black-shanked douc langurs, we used scan sampling (Altmann 1974) at 10-min intervals. We defined feeding as any occasion during which a langur plucked vegetation, pulled food items toward its mouth, masticated, or swallowed (Fashing 2001). Measurement of frequencies during scan sampling gives results similar to measures of feeding time (Clutton-Brock 1977a). Using 10 × 40 binoculars, we recorded a feeding score when a langur ate a leaf, fruit, seeds, flower, or other plant item. We recorded another feeding score if the same individual ate the same or another plant part from a different tree, or a different individual ate the same plant part from the same tree as the first individual. We scanned 1–7 individuals in 2-min periods. After a 10-min pause, if the group had not moved on, we scanned them again, starting with the focal individual. If the group moved on, we followed if the terrain allowed or searched for a new group if the terrain did not.
It was difficult to distinguish between leaves and leaf buds in the field, especially for Vitex spp. and Sindora siamensis, so we recorded leaves as a single class. We recorded fruits composed of pulp and small seeds, e.g., Ficus spp., or those with large seeds surrounded by pulp from which the douc langurs usually consumed the pulp, e.g., Canarium subulatum, Artocarpus rigida, as whole fruits. We pooled flowers and floral buds because it was difficult to distinguish between them at the distances that we were obliged to use for viewing, especially for Hopea ferrea and Syzydium ripicola.
Before scan sampling to quantify diet, we used focal animal sampling (Altman 1974) to determine an activity budget. This allowed more time to observe each individual and assign it to an age and sex class. The determinations derived during that sampling were used in the dietary observations that followed immediately after focal animal sampling. We reliably determined the age and gender of adults and juveniles, but could not assign a gender to infants.
Plant Collection
We collected botanical samples for plant identification from plant species on which we had observed black-shanked douc langurs feeding. We identified the plants either by direct observation or with the aid of fallen fruit, flowers, and leaves. We collected specimens either as soon as the langurs moved on or marked and collected them later. We immediately pressed and air-dried leaf samples. We kept fruits and flowers, which are essential for plant identification, in 70% ethanol (Barnett 1995). We classified plants according to Flora of Vietnam (Pham Hoang Ho 2000) and by reference to samples in the Nui Chua National Park herbarium.
Results
We collected frequency records from February to August 2005. We performed scans over 22 d (144 h of contact time) of observation, excluding days when visual contact time with groups was not long enough for any continuous scans. We recorded 903 scans, during which there were 604 that contained ≥1 feeding score. There was considerable variation in the amount of data collected in each month because of differences in the time available for observation and the visibility of the subjects. We took most samples during the wet season (Table III). In total, we observed 63 individual douc langurs feeding (Table IV). We observed only 2 juveniles feeding, but relatively even numbers of other gender/age classes across the 2 national parks (Table IV). We spent less time observing feeding in Phuoc Binh (68 h) than in Nui Chua (76 h), and most of the time in Phuoc Binh was in semideciduous forests.
In both national parks, the black-shanked douc langurs primarily fed on leaves, followed by fruit, then flowers (Table V). The monkeys consumed mostly leaves in every month and tended to eat more young leaves than mature ones. They most often obtained leaves by plucking them with one hand while holding a branch with the other. Less often they broke small branches and transferred the leaves directly to their mouths. Direct observation and inspection of remains on the ground showed that the black-shanked douc langurs ate more leaf laminas than petioles.
Fruit comprised ca. 30% of feeding scores (Table V), and langurs most commonly ate whole fruit (62.31% of all fruit feeding scores), followed by seed (31.26%) and pulp (6.43%). The langurs used either hand to pick whole fruits, which they usually broke into pieces with their hands before eating. They sometimes used their canine teeth to gouge out seeds so they could eat those alone. Flowers and flower buds comprised a considerable proportion of the diet of this species, while the category of other consisted of petioles (n = 3) and bamboo shoots (n = 3; Table V).
Food Proportions Over Time and Place
Black-shanked douc langurs consumed significantly more fruit and fewer flowers in the morning and vice versa in the afternoon (Table VI; χ2 = 9.15, df = 3, p < 0.05). The sample size in the dry season was small, but even so the proportions of food items showed a significant seasonal variation (χ2 = 51.49, df = 3, p < 0.005). During the dry season, the black-shanked douc langurs ate mostly leaves, followed by flowers, then fruit and other items (Fig. 2). In the wet season, the monkeys also ate leaves in greatest proportion but the proportion of fruits was almost double that in the dry season (Fig. 2).

Proportion of food items eaten by season (combined data for Phuoc Binh and Nui Chua).
Food Composition
The black-shanked douc langurs we observed ate parts of 152 plant species. However, only 140 of those could be identified (Table VII). Leguminosae (including Caesalpinoideae, Mimosoideae, and Papilionoideae) contributed 17 species to the langur's diet. Moraceae accounted for 13 species and was consumed in all months that we recorded. Ficus was particularly important and we observed the monkeys feeding on its unripe fruit, leaves, and leaf buds. Other families contributing multiple species to the diet were Euphorbiaceae, Rutaceae, Myrtaceae, and Guttiferaea (Table VII). Vitex spp. (Verbencaeae) were an important food source in March and April when these deciduous plants had new leaves.
Black-shanked douc langurs consumed leaves from 117 species, fruit from 42 species, and the petioles from 1 species that we could not identify. For some species the monkeys ate >1 plant part (Table VIII). Black-shanked douc langurs ate unripe whole fruit from 34 species, whole ripe fruit from 1 species, whole fruit from unripe to ripe from 5 species, and the ripe pulp from 2 species. They also ate the seeds from unripe fruit of 9 species and the seeds from ripe fruit of 2 species.
Submontane evergreen forest (54 species) and sclerophyll evergreen forest supported more food plants than other vegetation types. Only 2 food plant species were from mixed coniferous and broadleaf forest.
We observed 135 food plant species in Nui Chua, which is 10.67% of the 1265 plant species reported (Doan Canh et al. 2006), and 39 (7.60%) of the 513 plant species recorded in Phuoc Binh (Sub-FIPI, 2002). However, less time was spent observing feeding in Phuoc Binh than in Nui Chua.
The black-shanked douc langurs that we observed during spent most of their time feeding in the middle and upper storeys of the forest, where they consumed parts of trees. However, they did consume shrubs and lianas (Table IX). When the monkeys fed on understorey plants they were sometimes sitting on a rock or standing bipedally on the ground.
Discussion
The proportions of food items we observed are similar to what is known for other colobines (Table X). Black-shanked douc langurs were less folivorous than red-shanked douc langurs (Pygathrix nemaeus) but more folivorous than a black-shanked douc langur population in Cambodia (Table X). The differences may be related to food availability in the different study areas that we did not investigate. However, it is difficult to draw any firm conclusions because the data for Pygathrix nemaeus are based on short-term studies and with captives included.
Researchers have generally considered colobines highly selective feeders living in inhospitable environments (Waterman and Kool 1994), which is supported by our findings. We now know that Pygathrix nigripes does not have favorite food plants because we have demonstrated that it will eat various parts of ≥152 plant species, but only 20 species accounted for >2% of the total feeding scores recorded during scan sampling. Further investigation may well reveal a wider food selection because in this study the plant species consumed account for only 10.57% and 7.60% of the total plants inventory of Nui Chua and Phuoc Binh National Parks, respectively.
There was a switch in food selection during the day. While the proportions of leaves consumed before and after midday were similar, black-shanked douc langurs consumed more fruits and fewer flowers in the morning and vice versa after midday. This is consistent with the suggestion that leaf-eating monkeys consume mainly fruits in the morning because they provide readily accessible energy that is used to offset the animal's energy deficit immediately after sleep (Clutton-Brock 1977b; Oates 1987). However, it does not support the suggestion that animals should not eat many leaves in the morning to avoid carrying a full stomach while foraging (Chapman and Chapman 1991).
The small sample size in our study precludes any seasonal comparison of diet; however, our findings suggest the possibility of a seasonal shift in diet, which should be investigated further. In the months that we sampled (3 in the dry season and 4 in the wet) we observed that the black-shanked douc langurs ate a greater proportion of leaves and reduced their dietary diversity in the dry season. They also spent more time feeding and less time resting (unpubl. data). In the wet season the langurs ate a more diverse diet and increased the proportion of fruits in the diet. In Nui Chua National Park some local residents informed us that the black-shanked douc langurs did not often feed on the ground, but when they did it was mostly in the dry season. If there are seasonal changes they are most likely a response to changes in food availability. We did not conduct any surveys to determine the relationship between food eaten and its availability, but most trees in the 2 national parks flower from the mid dry season to early wet season and fruit in the wet season. These changes in food availability correlate with our observations on consumption. The small dry season sample size means that the results must be treated with caution, but we observed that in the wet season, when plant reproductive parts are more abundant, fruit and seeds combined accounted for 34% of the langur's total feeding score. The suggestion of a seasonal shift in diet has important implications for the conservation of the species and should be investigated further because it may be that different habitat is required in the wet season to the dry season.
The food plants consumed by the black-shanked douc langurs do contain a number of secondary compounds that are used in traditional Vietnamese medicine (Hoang 2007). We did not measure the compounds, and their selection by black-shanked douc langurs awaits investigation.
Conclusion
The black-shanked douc langurs that we observed had a predominantly folivorous diet, but the proportion of fruit and flowers increased in some sampling sessions. The most likely explanation is that in the dry season alternatives to leaves are unavailable. However, a firm conclusion cannot be reached until further study has examined the relationship between diet and phenological cycles. The list of known food plants has now been expanded to 152 species, and doubtless more will be added with further investigation. The black-shanked douc langurs that we observed seemed well able to satisfy their energy requirements in both wet and dry seasons. Hence even in harsh environments like Nui Chua National Park, the conservation of the species appears feasible if sufficient native forest can be maintained.
References
Altmann, J. (1974). Observational study of behaviour: Sampling methods. Behaviour, 49, 227–267. doi:10.1163/156853974X00534.
Baltzer, M. C., Nguyen, T. D., & Shore, R. G. (Eds.).(2001). Towards a vision for biodiversity conservation in the forests of the lower mekong ecoregion complex. World Wide Fund for Nature, Hanoi Vietnam.
Barnett, A. (1995). Expedition field techniques primates. London: Royal Geographical Society.
Bennett, E. L., & Sebastian, T. (1988). Social organisation and ecology of proboscis monkeys (Nasalis larvatus) in mixed coastal forest in Sarawak. International Journal of Primatology, 9, 233–256. doi:10.1007/BF02737402.
BirdLife International and Forest Inventory and Planning Institute (2001). Sourcebook of Existing and Proposed Protected Areas in Vietnam. Hanoi: Vietnam.
Bleisch, W. V., Cheng, A.-S., Ren, X.-D., & Xie, J.-H. (1993). Preliminary results from a field study of wild Guizhou snub-nosed monkeys (Rhinopithecis brelichi). Folia Primatologica, 60, 72–82. doi:10.1159/000156677.
Boonratana, R., & Canh, L. X. (1998). Preliminary observations of the ecology and behavior of the Tonkin snub-nosed monkey (Rhinopithecus [Presbytiscus] avunculus) in Northern Vietnam. In N. G. Jablonski (Ed.), The Natural History of the Doucs and Snub-nosed Monkeys (pp. 207–215). Singapore: World Scientific Publishing.
Chapman, C. A., & Chapman, L. J. (1991). The foraging itinerary of spider monkeys: When to eat leaves. Folia Primatologica, 56, 162–166. doi:10.1159/000156542.
Clutton-Brock, T. H. (1977a). Methodology and Measurement. In T. H. Clutton-Brock (Ed.), Primate Ecology: Studies of Feeding and Ranging Behaviour in Lemurs, Monkeys and Apes (pp. 585–589). New York: Academic Press.
Clutton-Brock, T. H. (1977b). Some aspects of intraspecific variation in feeding and ranging behaviour in primates. In T. H. Clutton-Brock (Ed.), Primate Ecology: Studies of Feeding and Ranging Behaviour in Lemurs, Monkeys and Apes (pp. 539–556). New York: Academic Press.
Ding, W., & Zhao, Q. K. (2004). Rhinopithecus bieti at Tacheng, Yunnan: Diet and daytime activities. International Journal of Primatology, 25, 583–598. doi:10.1023/B:IJOP.0000023576.60883.e5.
Doan Canh, Do Bich Loc, Truong Quang Tam, Pham Bach Viet, Nguyen Tran Vy, Hoang Duc Dat, Ngo Van Tri, Nguyen Quoc Thang, Ta Huy Thinh, and Nguyen Hong Hai. (2006). Survey on bio-resources and semi-arid ecosystem of Nui Chua National Park, Ninh Thuan Province. Institute of Tropical Biology, Hochiminh City (in Vietnamese).
Fashing, P. J. (2001). Feeding ecology of Guerezas in the Kakamega Forest, Kenya: The importance of Moraceae fruit in their diet. International Journal of Primatology, 22, 579–609. doi:10.1023/A:1010737601922.
Hoang, M. D. (2007). Ecology and Conservation Status of the Black-shanked Douc (Pygathrix nigripes) in Nui Chua and Phuoc Binh National Parks, Ninh Thuan Province, Vietnam. Ph.D. Thesis, School of Natural & Rural Systems Management, The University of Queensland, Australia.
Hoang Minh Duc.(unpublished). Preliminary results on the present status and the diet of black-shanked douc langur (Pygathrix nigripes) in Nui Chua Nature Reserve, Ninh Thuan province, Vietnam. Report to Primate Conservation Inc., and Institute of Tropical Biology, Hochiminh City, 2003.
IUCN World Conservation Union.(2006). 2007 Red list of threatened species. Available at http://www.iucnredlist.org/ (Accessed December 5, 2007).
Kirkpatrick, R. C. (1999). Colobine diet and social organization. In P. Dolhinow, & A. Fuentes (Eds.), The Nonhuman Primates (pp. 93–105). Mountain View, CA: Mayfield Publishing.
Lippold, L. K. (1977). The douc langur: A time for conservation. In H. S. H. Prince Rainier, & G. H. Bourne (Eds.), Primate Conservation (pp. 513–538). New York: Academic Press.
Lippold, L. K. (1998). Natural history of douc langurs. In N. G. Jablonski (Ed.), The Natural History of the Doucs and Snub-nosed Monkeys (pp. 191–206). Singapore: World Scientific.
Lippold, L. K., & Vu Ngoc Thanh (1998). Primate conservation in Vietnam. In N. G. Jablonski (ed.), The Natural History of the Doucs and Snub-nosed Monkeys (pp. 293–300). Singapore: World Scientific.
Lucas, P. W., & Teaford, M. F. (1994). Functional morphology of colobine teeth. In J. F. Oates, & A. G. Davies (Eds.), Colobine Monkeys: Their Ecology, Behaviour and Evolution (pp. 205–228). Cambridge: Cambridge University Press.
Nadler, T., & Momberg, F.Le Xuan Canh, and Lormee, N. (2003). Vietnam Primate Conservation Status Review 2002: Part 2 Leaf Monkeys. Hanoi, Vietnam. Frankfurt Zoological Society-Cuc Phuong National Park Conservation Program, Fauna & Flora International Program.
Napier, J. R., & Napier, P. H. (1985). The Natural History of the Primates. Cambridge, UK: Cambridge University Press.
National Environment Agency (2005). Environment Status of Vietnam: Biodiversity. Ha Noi: National Environment Agency.
Nhat, P. (1993). First results on the diet of the Red shanked Douc Langur (Pygathrix nemaeus). Australasian Primatology, 8, 5–6.
Nhat, P., Huy, D. Q., & Nguyen, P. H. (2000). Report on Research Result on Distribution, Ecology and Monitoring Survey of the Red-shanked Douc Langurs (Pygathrix nemaeus nemaeus) in Phong Nha-Ke Bang Forest Area. WWF- Indochina.Programme, Hanoi.
Nui Chua National Park. (2005). 2004 Annual report on forest status of Nui Chua National Park. Nui Chua National Park, Phan Rang.
Oates, J. F. (1987). Food distribution and foraging behavior. In B. B. Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham, & T. T. Struhsaker (Eds.), Primate Societies (pp. 197–209). Chicago: The University of Chicago Press.
Otto, C. (2005). Food Intake, Nutrient Intake, and Food Selection in Captive and Semi-free Douc Langur. Munster: Schuling Verlag.
Pham Hoang Ho. (2000). The Flora of Vietnam (Cay co Vietnam in Vietnamese), Volumes 1, 2 and 3. Youth Publisher, Hochiminh City (in Vietnamese).
Phuoc Binh Commune Peoples Committee.(2005). Annual report of Phuoc Binh Commune People Committee, Phuoc Binh (in Vietnamese).
Rawson, B. (2006). Activity budget in black-shanked douc langur (Pygathrix nigripes), International Journal of Primatology, 27(Supp. 1), Abstract 307.
Sub-FIPI. (1996). Feasibility study of Nui Chua Nature Reserve. Sub Institute of Forest Inventory and Planning II, Hochiminh City (in Vietnamese).
Sub-FIPI. (2000). Investment plan for Phuoc Binh Nature Reserve, Sub Institute of Forest Inventory and Planning II, Hochiminh City (in Vietnamese).
Sub-FIPI. (2002). Investment Project for Phuoc Binh Nature Reserve and Buffer zone: 2003–2007. SubInstitute of Forest Inventory and Planning II. Unpublished report to Ninh Thuan Provincial People Committee, Phan Rang (in Vietnamese).
Thai Van Trung (1978). Vegetation of Vietnamese forests. Science and Technology Publishing House: Hanoi. (in Vietnamese).
Vu, N. L., Hoang, M. D., Luu, H. T., Le, B. T., Diep, D. P., & Nguyen, P. N. (2001). Biodiversity and social studies of Nui Chua Nature Reserve in Ninh Thuan Province, Vietnam. Scientific report to Japan NAGAO. National Environment Foundation. Mo Chi Minh City.
Waterman, P. G., & Kool, K. M. (1994). Colobine food selection and plant chemistry. In A. G. Davies, & J. F. Oates (Eds.),Colobine Monkeys: Their Ecology, Behaviour and Evolution (pp. 251–284). Cambridge, UK: Cambridge University Press.
Wege, D., & Long, A. J., Mai Ky Vinh, Vu Van Dung, and Eames, J. C. (1999). Expanding the Protected Area Network in Vietnam for the 21st Century: an Analysis of the Current System with Recommendations for Equitable Expansion', Forestry Inventory and Planning Institute and BirdLife International Vietnam Program, Hanoi.
Acknowledgments
Conservation International, Primate Conservation Inc., SeaWorld & Busch Gardens, and The University of Queensland, Australia provided funding for this study. Luu Hong Truong and Ly Ngoc Sam from The Institute of Tropical Biology assisted in identification of plants.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Duc, H.M., Baxter, G.S. & Page, M.J. Diet of Pygathrix nigripes in Southern Vietnam. Int J Primatol 30, 15–28 (2009). https://doi.org/10.1007/s10764-008-9325-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-008-9325-y