
Abstract
Understanding and modelling the development of lake phytoplankton communities is a desirable goal, given the importance of these organisms to their ecosystem. PROTECH (Phytoplankton RespOnses To Environmental CHange) is one such model which attempts to do this and its applications over the last 10 years are reviewed here. These studies include: modelling very large lakes, linking catchment models to PROTECH, simulating oxygen concentrations, understanding the importance of nutrient source in moderating the influence of hydraulic retention time. Furthermore, the merits of ensemble lake modelling are considered, as are the limits of short term forecasting of blooms. Finally, climate change influences are examined with studies that include nutrient changes and an experiment that attempts to separate the influences of temperature and mixed depth.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Understanding the responses of ecosystems to drivers lies at the heart of most ecological research. Through quantification of the environment, mathematical relationships can sometimes be found which describe these responses leading to the possibility for prediction and modelling. This quantification can be derived from observational field data or experiments and it was the latter that Colin Reynolds used in the 1980s to explore the nature of phytoplankton growth rates.
Reynolds (1989), through a series of laboratory experiments under idealised conditions, found relationships between the growth rates of a range of different lake phytoplankton species and their morphology. Thus, if the surface area, volume and maximum linear dimension of a species were known, the response of its growth rate to changes in temperature and light availability could be estimated using equations derived from Reynolds’ experiments.
Following this work, Reynolds went on to begin the construction of a computer model with a colleague, Anthony Irish. At its heart lies the equations of Reynolds (1989) but further enumeration was required to allow for the effects of nutrient limitation upon growth rates and various loss processes such as zooplankton grazing, sedimentation and flushing removal. With the final addition of species-specific daily vertical movements, the biological core of the new PROTECH (Phytoplankton RespOnses To Environmental CHange) model was complete. The conception, equations and first 10 years of research of the PROTECH model was extensively reviewed in Elliott et al. (2010), but this review will consider the new PROTECH research from the last 10 years after a brief overview of the model.
The PROTECH model
PROTECH is a 1D-lake phytoplankton community model that works in daily time steps. The spatial structure is constructed using bathymetry data describing the changing surface area and volume. This creates a series of vertical layers in the model, each 0.1 m deep. The physical thermal profile can be either calculated based on daily meteorological inputs (air temperature, wind speed and air humidity) and inflow water temperature or it can be read in from observed data or another model. A further physical consideration is to model water exchange in the lake through inflow and outflow discharge. In PROTECH, it is assumed the water entering the lake mixes with the surface mixed layer and the water leaving the lake also comes from this layer unless it is a reservoir system where water is abstracted at depth.
The biological component is primarily focused on the phytoplankton community. The individual phytoplankton are characterised by their morphology, daily movement, vulnerability to grazing and nutrient requirements to reflect nitrogen-fixation and diatoms (Table 1). The morphology information is used by the equations from Reynolds (1989) to create phytoplankton specific potential growth rates for each depth layer in the model, responding to changing temperature and light availability. The availability of nutrients to support this growth is checked using specific thresholds (3, 80, 500 mg m−3 for phosphorus, nitrate and silica, respectively) and the growth rate reduced proportionally if any nutrients are limiting.
Phytoplankton biomass loss is caused by filtration-based zooplankton grazing (the phytoplankton maximum dimension must be < 50 µm, although there can be exceptions based on natural observations), sedimentation out of the water column and dilution wash-out caused by water exchange. Thus, after balancing the growth with the losses, a daily net gain in biomass can be calculated for each layer in the model. Clearly, the model is more detailed than this brief overview suggests and for further information it is recommended to consult Elliott et al. (2010). Finally, at the time of the Elliott et al. (2010) review, PROTECH had been applied in the peer-reviewed literature to ten water bodies, predominately in the UK (Table 2). However, since then the model has been used in many more new studies (Table 2), which are considered in this review.
Applying PROTECH to large lakes
Pre-2010, PROTECH had mainly been applied to relatively small lakes with the exception of Lake Erken, Sweden (24 km2; Elliott et al., 2007). However, since then it has been applied to the UK’s largest lake, Lough Neagh (383 km2; Elliott et al., 2016), Pyhäjärvi in Finland (154 km2; Pätynen et al., 2014) and Lake Simcoe, Canada (2899 km2; Crossman & Elliott, 2018; Crossman et al., 2019). Given the model’s 1D nature, these studies were a new challenge and for the first two studies, yet PROTECH performed satisfactory with only one modification regarding sediment nutrient release, which is discussed below. However, for the very large Lake Simcoe, the lake had to be divided into three separate basins where two side arms fed into the large main basin and this approach worked well with the outflows from the separate side arm simulations becoming inflows into the main basin simulation. It is amusing to note that these two “side arms” were individually a similar size to England’s largest lake, Windermere!
Furthermore, these studies highlighted a weakness in PROTECH regarding the model’s lack of functions to elucidate nutrient release from sediments. In such large surface area water bodies, sediment inputs can be an important source of nutrients, particularly phosphorus, and from the three lakes discussed above a forced input of phosphorus had to be added to the model using observed in-water nutrient data as a guide. Some other lake models do include such functions (e.g. PCLake (Janse, 1997)) and it is something that will hopefully be added to PROTECH in the future. However, one of the problems is that such a function would require knowledge of nutrient sediment concentrations, something that is rarely measured in standard programmes of lake monitoring. This lack of data has certainly been the case in many previous PROTECH studies, hence the forced introduction of nutrients to the water column using in-lake measurements of nutrients.
Linking the catchment to the lake through modelling
Lakes are intrinsically part of their catchment and are affected by the types of land that make up that area. Given this connection, it is understandable that the management of catchments is a prevalent method for reducing nutrient inputs to lakes and thus improve their trophic status. It also follows that using models that can adequately describe this connection are very useful in exploring how changes in land use might impact upon the lake phytoplankton.
An example of such a cascade of different models is Norton et al.’s (2012) study of Loweswater, UK, where a catchment model’s (GWLF; Schneidermann et al., 2002) output was used to drive PROTECH. Loweswater was suffering from poor water quality and cyanobacteria blooms due to high nutrient inputs from the surrounding farm land. The investigation tested the relationship between these nutrient inputs and the phytoplankton produced by the lake through the application of a number of different land use scenarios. These covered different ratios of livestock (cattle and sheep) as well as more extreme scenarios such as no livestock with all grassland or all woodland.
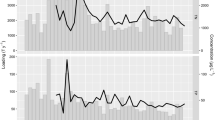
The wide range of phosphorus loads created by the GWLF model and these nutrient outputs were used to drive PROTECH. The resultant outputs from the two models allowed the relationship between nutrient load and the modelled phytoplankton community to be described (Fig. 1). This was interesting because it showed that PROTECH produced two different responses to the changing phosphorus loads. For example, the annual mean chlorophyll produced by PROTECH grew with increasing loads producing an asymptotic-like curve whereas the increase in the cyanobacteria part of the community was linear (Fig. 1). Thus, the study suggested there was an escalating trend cyanobacteria dominance within the community with increasing input nutrient load, despite a declining rate in production of overall biomass.

Adapted from Norton et al. (2012) removing the individual scenario data points to show only the relationships
The modelled relationship between changing Loweswater catchment soluble reactive phosphorus (SRP) load and annual mean concentrations of total (solid line) and cyanobacteria (dashed line) chlorophyll a
Modelling oxygen concentrations in PROTECH
Oxygen was not a variable originally considered in PROTECH, so in a study that wished to simulate the potential impact of climate change on the Vendace [Coregonus albula (Linnaeus, 1758)] fish species, the Lake OXygen model (LOX; Bell et al., 2006) was added into PROTECH (Elliott & Bell, 2011). LOX works by dividing the vertical component of the lake into two layers (epilimnion and hypolimnion) and its equations were used to create a new sub-routine in PROTECH. By using PROTECH’s temperature and chlorophyll data, LOX can estimate the dissolved oxygen concentrations in the two layers and through a simple depth function create an oxygen profile from the surface to the bottom of the lake.
If the approach of other lake models is considered, LOX’s complexity is comparable to PCLake’s oxygen calculations (Janse, 1997), but is simplistic compared to DYRESM-CAEDM (Hamilton & Schladow, 1997), which uses process-based calculation throughout all of its layers, Nevertheless, LOX proved effective both for the Vendace study in Bassenthwaite Lake, UK (R2 > 0.7 against fortnightly observed O2; Elliott & Bell, 2011) and for the Lake Simcoe, Canada, study (R2 > 0.8 against monthly mean observed O2; Crossman & Elliott, 2018) where the impacts of catchment land use changes on the lake’s oxygen concentrations were simulated.
The effect of changes in hydraulic retention time
Building upon previous PROTECH studies (Elliott et al., 2009; Elliott, 2010), further work explored the importance of changing hydraulic retention time upon phytoplankton. Many aspects of weather are predicted to change in the future and in a study focused on the eutrophic Loch Leven, UK, PROTECH was used to assess the relative importance of increasing water temperature and changing retention time (Elliott & Defew, 2012). The study showed clearly that changes in inflow, both increase and decreases, were of greater importance than temperature increase as large as 4°C. In general, increased inflows which caused a reduction in retention time were detrimental to the slower growing species in the model leading to the heightened presence of smaller, faster growing phytoplankton. However, this flow effect had a seasonal aspect to it because at times of year with naturally low flows (e.g. summer), an increase actually benefited the phytoplankton by providing more nutrients at a time when they were limiting growth. This was because the source of nutrients to the lake was assumed to be diffuse leading to an increase in flow delivering a greater nutrient load to the lake.
This importance of nutrient source was explored further in a PROTECH study of Bassenthwaite Lake, UK (Jones et al., 2011). This model experiment recognised the relationship between nutrient load to a lake and the type of nutrient source. Specifically, this means a load derived from a point nutrient source is independent of the flow whereas a diffuse source load changes in proportion with inflow. To test the importance of source, 32-year flow scenarios were run based on a number of climate change scenarios. These runs provided a large range of retention times over which annual and seasonal means could be calculated.
The results showed that the sensitivity of nutrient source to changes in flow was seasonally dependent: winter and autumn were the least responsive with spring and summer being the most sensitive. Furthermore, for the sensitive seasons, the nature of the relationship was greatly dependent upon nutrient source. Thus, for the flow-independent scenarios short retention times produced less phytoplankton biomass than when retention time was long (Fig. 2). Conversely, with flow dependence, high inflows increased biomass and low flows saw little decline in chlorophyll (Fig. 2). The mechanism behind these differences was the balance between the dilution loss of biomass caused by high flows and the potential to bring more nutrients into the lake and thus stimulate more phytoplankton growth. Therefore, whilst the former is a universal effect with high flows, the latter can only happen under the flow-dependent (diffuse) conditions.

Illustration of the modelled relationship between summer mean chlorophyll and retention time when the nutrient source for the lake is either flow independent (solid line) or flow dependent (dotted line)
Ensemble modelling
Uncertainty in model parameters and construction is a constant issue. An approach used in other disciplines (e.g. weather forecasting) is to apply different versions of the same model (the most common approach) or different models (much rarer) to the same scenarios and then amalgamate the results to provide a predicted mean and uncertainty envelope. In ecology, though, it is a method that seems to be rarely used therefore, in Trolle et al. (2014) an ensemble of different lake models was applied to Lake Engelsholm, Denmark.
Three lake models formed the ensemble: DYRESM-CAEDYM (Hamilton & Schladow, 1997), PCLake (Janse, 1997) and PROTECH. It should be noted that this approach of using independently created models for the ensemble is rare and clearly has a greater capability of capturing more levels of uncertainty than would be gained from simply using re-parametrisations of one model. For calibration and validation, a 3-year simulation was conducted and model performance at the daily and monthly level was assessed using observation data to determine the coefficient of determination (r2) and relative absolute error. These statistics showed clearly that the mean values created from the three individual models’ outputs were a better fit to the observations than any individual model managed to produce. Coupled with that, the combination of the models also produced an uncertainty range using the minimum and maximum values from the models. This showed that the greatest area of uncertainty was during the summer where phytoplankton biomass was at its highest. Using this baseline, a number of climate change scenarios were then run for the lake and showed that small increases in water temperature increased both the total phytoplankton and cyanobacteria biomass.
Forecasting phytoplankton abundance
Being able to forecast phytoplankton abundance in a way similar to weather forecasts is a challenging goal but it was attempted recently using PROTECH. This ambitious programme of research had two phases, the first being to test and understand the behaviours of PROTECH when driven by high frequency in-lake observations (Page et al., 2018). This was done using sub-daily buoy data collected from three lakes in the English Lake District (Blelham Tran, Esthwaite Water and Windermere). The results revealed that the most difficult aspect of forecasting was not the future weather uncertainty but rather the uncertainty surrounding the daily nutrient load to the lake.
Building upon this initial testing, a second phase of modelling used historic weather forecasts that used an ensemble of 50 simulations of 10-days-ahead weather to drive the model coupled with stochastic perturbations of model parameters (Page et al., 2018). This allowed PROTECH, day by day, to forecast a range of possible futures and create an uncertainty envelope. Windermere and Esthwaite Water were used for these tests and the results were compared to the benchmark prediction of persistence i.e. the chlorophyll concentration will not change over the 10-day forecast period and is reset to equal new observation data when they become available every 2 weeks. Given this criteria, PROTECH was only better than the persistence forecast at a forecast range of less than 6 days. In terms of the cyanobacteria forecast, the model was not successful of predicting its biomass, showing the limitations of the approach at predicting specific species although it was more successful at predicting functional types i.e. low-light or low-nutrient specialists.
Climate change and nutrient load impacts
Building on previous PROTECH investigations (Elliott et al., 2006; Elliott & May, 2008), two additional studies were conducted examining the combined impact changing temperature and nutrient load. These studies explored lakes that were different to the previous studies, specially a larger lake (Windermere, UK) and a deep lake with a small surface area (Rostherne Mere, UK).
Windermere is England’s largest lake (64 m deep, 14.7 km2 surface area) and has been a focal point for tourism in the English Lake District for over 100 years (McGowan et al., 2012). Correspondingly, the lake has been under ever-growing anthropological pressure both in terms of its usage and ecosystem health. How those pressures would affect Windermere in the future was the subject of a PROTECH investigation where water temperature and nutrient load were altered in a factorial modelling experiment (Elliott, 2012).
Assessing the impact of these changes, it was clear that the simulated spring diatom bloom was more influenced by the changes in temperature than nutrients, showing earlier bloom peaks with increasing temperature. However, in the summer period increasing nutrient load and water temperature synergised to enhance cyanobacteria growth. This led to more days of exceedance of the World Health Organisation’s cyanobacteria threshold of 10 mg m−3, but nutrients were the crucial factor with the reduced nutrient scenarios greatly limiting the temperature impacts.
Rostherne Mere is a very different lake to Windermere with a depth of 31 m but a surface area of only 0.49 km2 (Radbourne et al., 2019). This basin shape leads to it having a long, stable period of stratification lasting 9–10 months to a depth of 10 m. Historically, the lake has suffered from eutrophication driven by external nutrient sources, which led to the diversion upstream of sewage effluent in 1991. However, due to the strong stratification in the lake and corresponding sediment nutrient release, recovery has been very slow (Moss et al., 2005). Therefore, to explore the potential future trajectories of recovery for Rostherne Mere, the PROTECH model was applied (Radbourne et al., 2019).
The study used future climate scenarios from the UKCP09 projections (Murphy et al., 2009) and coupled them with a range of external and internal nutrient scenarios. The results reinforced the importance of the internal nutrient problem, quantifying that substantial reductions in this source for decades would be needed to see improvements in the lake. However, the scenarios towards the end of twenty-first century actually showed a reduction in phytoplankton biomass because, with an increase in stratification length, the reliance by phytoplankton upon external nutrients increased and given that this source had been reduced, late summer biomass declined. Nevertheless, the winter overturn of the water column still brought the considerable amounts hypolinimon nutrients to the surface, meaning that long-term recovery was still hindered.
Disentangling mixed depth and temperature effects
The thermal structure of a lake varies annually in temperate regions with the formation and dissipation of stratification occurring in many lakes. Physically, temperature and mixed layer depth vary together as both variables can influence each other. However, changes in temperature and mixed depth effect phytoplankton communities in different ways with the former influencing cell metabolic processes and the latter effecting light and nutrient availability. Given this correlation between temperature and mixing, it is very difficult to attribute cause and relative importance to any observed changes in the lake phytoplankton. Therefore, a PROTECH experiment on Blelham Tarn, UK, was conducted to separate these relative effects by forcing the modelled lake structure (Gray et al., 2019). Essentially, this meant artificially manipulating the mixed depth in the model independently of changing the temperature and vice versa. Whilst such a thing would be highly unlikely in the real world, it did allow great insight into the relative importance of changes in mixing and temperature. For example, whilst stratification length was prevented from changing in the model scenarios, increasing temperature alone caused an earlier spring bloom by accelerating growth rates in the phytoplankton. Given such advances in bloom timing have been observed in lakes (e.g. Thackeray et al., 2010), this model experiment shows that although changes in stratification can be a cause of such advancement, temperature alone is capable of producing the same effect.
Another emergent model result concerned cyanobacteria blooms in the summer and autumn periods. Here, two different types of cyanobacteria dominated with different mixed depths as temperature increased. Thus, the Dolichospermum-type in PROTECH thrived with shallow mixing whilst the Plantothrix-type dominated with deep mixing. This suggested that a shallowing of mixed depth in lakes where mixing is not too deep (e.g. < 5 m) would see greater blooms of buoyant cyanobactera. Conversely, lakes that currently experience blooms of low-light tolerant phytoplankton, like Plantothrix, might see a decline in their dominance if the deeper mixing (e.g. > 10 m) seen in those lakes shallowed in the future.
The legacy of Colin Reynolds: PROTECH
This review forms part of a special issue celebrating the legacy of Colin Reynolds who sadly passed away in December 2018. That legacy includes PROTECH, a model which Colin was so fundamental in conceiving and creating. When I was lucky enough to fall under Colin’s mentorship and begin to use PROTECH, we used to joke that the model was a digitization of his brain, taking his great knowledge of phytoplankton and turning it into the “0 s and 1 s” binary of the computing world!
Of course, since those early days, PROTECH has continued to be applied to lakes all around the world, flourishing and developing to face new challenges and creating a growing body of research, the latest of which has been reviewed here. I have no doubt that in the future PROTECH will continue to contribute to our understanding of lakes and their phytoplankton, and that through such work, an important part of Colin’s legacy will live on.
Finally, I would like to record here my heartfelt thanks to Colin for all the support he gave me over the years: he was a great mentor, colleague and, perhaps most importantly, friend. You are missed by me, every day.
References
Bell, V. A., D. G. George, R. J. Moore & J. Parker, 2006. The use of a 1-D mixing lake model and high resolution data to assess the impact of extremes of wind and temperature on lake dynamics. Ecological Modelling 190: 41–54.
Bernhardt, J., J. A. Elliott & I. D. Jones, 2008. Modelling the effects on phytoplankton communities of changing mixed depth and background extinction coefficient on three contrasting lakes in the English Lake District. Freshwater Biology 53: 2573–2586.
Crossman, J. & J. A. Elliott, 2018. Bridging the gap between terrestrial, riverine and limnological research: application of a model chain to a mesotrophic lake in North America. Science of The Total Environment 622–23: 1363–1378.
Crossman, J., M. N. Futter, J. A. Elliott, P. G. Whitehead, L. Jin & P. J. Dillon, 2019. Otimizing land management strategies for maximum improvements in lake dissolved oxygen concentrations. Science of the Total Environment 652: 382–397.
Elliott, J. A., 2010. The seasonal sensitivity of Cyanobacteria and other phytoplankton to changes in flushing rate and water temperature. Global Change Biology 16: 864–876.
Elliott, J. A., 2012. Predicting the impact of changing nutrient load and temperature on the phytoplankton of England’s largest lake, Windermere. Freshwater Biology 57: 400–413.
Elliott, J. A. & V. A. Bell, 2011. Predicting the potential long-term influence of climate change on vendace (Coregonus albula) habitat in Bassenthwaite Lake, U.K. Freshwater Biology 56: 395–405.
Elliott, J. A. & L. Defew, 2012. Modelling the response of phytoplankton in a shallow lake (Loch Leven, UK) to changes in lake retention time and water temperature. Hydrobiologia 681: 105–116.
Elliott, J. A. & L. May, 2008. The sensitivity of phytoplankton in Loch Leven (UK) to changes in nutrient load and water temperature. Freshwater Biology 53: 32–41.
Elliott, J. A. & S. J. Thackeray, 2004. The simulation of phytoplankton in shallow and deep lakes using PROTECH. Ecological Modelling 178: 357–369.
Elliott, J. A., A. E. Irish, C. S. Reynolds & P. Tett, 2000. Modelling freshwater phytoplankton communities: an exercise in validation. Ecological Modelling 128: 19–26.
Elliott, J. A., C. Escot, A. Basanta-Alves & L. Cruz-Pizarro, 2005. Simulations of phytoplankton dynamics in El Gergal reservoir, southern Spain (PROTECH). In K. Rouen & G. George (eds) New Tools for the Monitoring, Modelling & Management of Fresh Waters. Freshwater Forum 23: 78–92.
Elliott, J. A., I. D. Jones & S. J. Thackeray, 2006. Testing the sensitivity of phytoplankton communities to changes in water temperature and nutrient load, in a temperate lake. Hydrobiologia 559: 401–411.
Elliott, J. A., I. Persson, S. J. Thackeray & T. Blenckner, 2007. Phytoplankton modelling of Lake Erken, Sweden by linking the models PROBE and PROTECH. Ecological Modelling 202: 421–426.
Elliott, J. A., I. D. Jones & T. Page, 2009. The importance of nutrient source in determining the influence of retention time on phytoplankton: an explorative modelling study of a naturally well-flushed lake. Hydrobiologia 627: 129–142.
Elliott, J. A., A. E. Irish & C. S. Reynolds, 2010. Modelling phytoplankton dynamics in fresh waters: affirmation of the PROTECH approach to simulation. Freshwater Reviews 3: 75–96.
Elliott, J. A., Y. McElarney & M. Allen, 2016. The past and future of phytoplankton in the UK’s largest lake, Lough Neagh. Ecological Indicators 68: 142–149.
Gray, E., J. A. Elliott, E. B. Mackay, A. M. Folkard, P. O. Keenan & I. D. Jones, 2019. Modelling lake cyanobacterial blooms: disentangling the climate-driven impacts of changing mixed depth and water temperature. Freshwater Biology 64: 2141–2155.
Hamilton, D. P. & S. G. Schladow, 1997. Prediction of water quality in lakes and reservoirs. Part 1: model description. Ecological Modelling 96: 91–110.
Hutchins, M. G., C. Abesser, C. Prudhomme, J. A. Elliott, J. P. Bloomfield, M. M. Mansour & O. E. Hitt, 2018. Combined impacts of future land-use and climate stressors on water resources and quality in groundwater and surface waterbodies of the upper Thames river basin, UK. Science of the Total Environment 631–632: 962–986.
Janse, J. H., 1997. A model of nutrient dynamics in shallow lakes in relation to multiple stable states. Hydrobiologia 342–343: 1–8.
Jones, I. D., T. Page, J. A. Elliott, S. J. Thackeray & A. L. Heathwaite, 2011. Increases in lake phytoplankton biomass caused by future climate-driven changes to seasonal river flow. Global Change Biology 17: 1809–1820.
Lewis, D. M., J. A. Elliott, M. F. Lambert & C. S. Reynolds, 2002. The simulation of an Australian reservoir using a phytoplankton community model (PROTECH). Ecological Modelling 150: 107–116.
McGowan, S., P. Barker, E. Y. Haworth, P. R. Leavitt, S. C. Maberly & J. Pates, 2012. Humans and climate as drivers of algal community change in Windermere since 1850. Freshwater Biology 57: 260–277.
Moss, B., T. Barker, D. Stephen, A. E. Williams, D. J. Balayla, M. Beklioglu & L. Carvalho, 2005. Consequences of reduced nutrient loading on a lake system in a lowland catchment: deviations from the norm? Freshwater Biology 50: 1687–1705.
Murphy, J. M., Sexton, D. M. H., Jenkins, G. J., Booth, B. B. B., Brown, C.C., Clark, R. T., … Wood, R. A. 2009. UK Climate Projections Science Report: Climate Change Projections. Exeter, UK: Meteorological Office Hadley Centre.
Norton, L., J. A. Elliott, S. C. Maberly & L. May, 2012. Using models to bridge the gap between land use and algal blooms: an example from the Loweswater catchment, UK. Environmental Modelling & Software 36: 64–75.
Page, T., P. J. Smith, K. J. Beven, I. D. Jones, J. A. Elliott, S. C. Maberly, E. B. Mackay, M. De Ville & H. Feuchtmayr, 2018. Adaptive forecasting of phytoplankton communities. Water Research 134: 74–85.
Pätynen, A., J. A. Elliott, P. Kiuru, J. Sarvala, A. Ventelä & R. I. Jones, 2014. Modelling the impact of higher temperature on the phytoplankton of a boreal lake. Boreal Environmental Research 19: 66–78.
Radbourne, A. D., J. A. Elliott, S. C. Maberly, D. B. Ryves & N. J. Anderson, 2019. The impacts of changing nutrient load and climate on a deep, eutrophic, monomictic lake. Freshwater Biology 64: 1169–1182.
Reynolds, C. S., 1989. Physical determinants of phytoplankton succession. In Sommer, U. (ed.), Plankton Ecology. Brock-Springer, New York: 9–55.
Reynolds, C. S.,A. E. Irish & J. A. Elliott, 2005. A modelling approach to the development of an active management strategy for the Queen Elizabeth II reservoir. In K. Rouen & G. George (eds), New Tools for the Monitoring, Modelling & Management of Fresh Waters. Freshwater Forum 23: 105–125.
Schneidermann, E. M., D. C. Pierson, D. G. Lounsbury & M. S. Zion, 2002. Modeling of hydro-chemistry of the cannonsville watershed with Genearlized Watershed Loading Functions (GWLF). Journal of the American Water Resources Association 38: 1323–1347.
Thackeray, S. J., Sparks, T. H., Frederiksen, M., Burthe, S., Bacon, P. J., Bell, J. R., … S. Wanless, 2010. Trophic level asynchrony in rates of phenological change for marine, freshwater and terrestrial environments. Global Change Biology 16: 3304–3313.
Trolle, D., J. A. Elliott, W. M. Mooij, J. H. Janse, K. Bolding, D. P. Hamilton & E. Jeppesen, 2014. Advancing projections of phytoplankton responses to climate change through ensemble modelling. Environmental Modelling & Software 61: 371–379.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Guest editors: Judit Padisák, J. Alex Elliott, Martin T. Dokulil & Luigi Naselli-Flores / New, old and evergreen frontiers in freshwater phytoplankton ecology: the legacy of Colin S. Reynolds
Rights and permissions
About this article

Cite this article
Elliott, J.A. Modelling lake phytoplankton communities: recent applications of the PROTECH model. Hydrobiologia 848, 209–217 (2021). https://doi.org/10.1007/s10750-020-04248-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-020-04248-4