
Abstract
Separating the effects of anthropogenic changes in freshwater delivery from that of sea level rise on the rate of salt water encroachment in the low relief Southeast Saline Everglades is important for understanding how the Anthropocene Marine Transgression might be best managed. We use stratigraphic and paleoecologic methods to calculate rates of salt water encroachment and biogenic sediment accumulation in the Southeast Saline Everglades. Our results suggest that sea level rise during the last century was accompanied by salt water encroachment, which is ultimately controlled by the elevation of high tide and varied by a factor of 14.8 in the five watersheds studied. These differences are attributed primarily to differences in freshwater delivery. The delivery of freshwater mitigated salt water encroachment in only one of the five watersheds. This difference is attributed to sufficient freshwater delivery to maintain a plant community with more rapid rate of sediment accumulation than other sites. Under conditions of diminishing freshwater availability and increasing rate of sea level rise, our data suggest that little can be done at a scale large enough to prevent loss of the Southeast Saline Everglades within the next 50–200 years.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The Southeast Saline Everglades (SESE) of South Florida, USA, historically consisted of a narrow belt of mangroves along the coast with freshwater marshes extending inland to the outcropping limestone of the Atlantic Coastal Ridge (Fig. 1). The Atlantic Coastal Ridge (Ridge) separates the Everglades from the SESE and gaps in the Ridge allow Everglades surface water delivery to the SESE. Delivery was historically sufficient to maintain freshwater marshes all the way to the coast. However, natural water flow through the Everglades is reduced by 70% (Perry, 2004) in response to increasing urbanization and associated alteration in water use and delivery patterns.

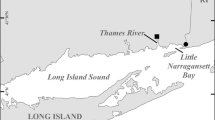
Location of field stations and transects: transects are bold lines with bold labels (TS Taylor Slough, JB Joe Bay, HC Highway Creek, TP Turkey Point, and MC Mowry Canal), small labeled circles are field station locations and the SESE is the white coastal band
Egler (1952) first applied the term SESE to the portion of the Everglades subject to marine influence as a consequence of proximity to two marine bodies of water, Florida and Biscayne Bays. Sedimentological data indicate that coastal wetlands (i.e., fringing Rhizophora mangle Linnaeus forests) in the SESE were in equilibrium with respect to maintaining surface elevation in response to SLR for at least the last 3165 years (Unpublished radiocarbon date # 1229, University of Miami Radiocarbon Laboratory). However, over the last century mangroves have retreated over the freshwater wetland prairie community at the expense of freshwater wetland (Ross et al., 2000, 2002). The landward migration and expansion of mangrove habitat is either a response to rising sea level, the decrease in freshwater delivery, or a combination of the two processes. Distinguishing between the effects of SLR and anthropogenic alterations in freshwater delivery to the coastal area is thus important in order to provide a better technical foundation for restoration activities.
In South Florida, the rate of late Holocene sea level rise (SLR) during the previous 3 k year averaged <1 mm year−1 (Wanless et al., 1994). Over the last century, the rate of SLR in South Florida accelerated to 2.4 mm year−1 (Maul & Martin, 1993). Since the year 2000, the rate of SLR in South Florida increased to between 5.9 and 9 mm year−1 (Park and Sweet, 2015; Wdowinski et al., 2016). This acceleration is consistent with global trends in SLR described by Nicholls & Cazenave (2010). The present rate of global SLR is 3.3 mm year−1 (Church & White, 2011; NASA, 2016) (Fig. 2). By the end of this century, the rate of global SLR is expected to continue accelerating, with estimates ranging between 9.6 mm year−1 (IPCC, 2013) and 20 mm year−1 (NOAA, 2012). Although there will surely be regional and local variations (Hay et al., 2015), it is logical to assume a continued acceleration in SLR along the South Florida coast driven in large part by global eustatic trends.

This historical acceleration in the rate of SLR is linked to global climate change. The rate and magnitude of SLR will continue to increase well beyond the turn of the century (c.f. Parkinson et al., 2015) and is significant because it determines coastal response (Wanless et al., 1994; Table 2 in Parkinson et al., 2015). Studies of the South Florida Holocene suggest three temporal subdivisions based upon the rate of SLR and associated coastal response (Davis, 1940; Scholl et al., 1969; Wanless, 1974; Spackman et al., 1976; Parkinson, 1989; Parkinson & Meeder, 1991; Wanless et al., 1994). The rate of SLR during the early Holocene was >10 mm year−1 and coasts underwent submergence and overstep. During the middle Holocene (the time interval between 6 and 3 k year B.P.) the rate of SLR slowed to between 1 and 2 mm year−1, resulting in the formation of barrier islands and other constructional coastal features that retreated landward to generate a transgressive facies succession. During the last 3 k year (late Holocene) the rate of SLR decreased to <1 mm year−1 resulting in stabilized coastlines, aggradation, and progradation. The conclusions drawn by these South Florida studies are further supported by investigations from other regions (Grand Cayman, Woodroffe, 1981; Cayman Islands, Ellison & Stoddart, 1991; Bermuda, Ellison 1993; Dieman Gulf, Australia, Woodroffe, 1995).
The rate and distance of salt water encroachment (SWE) and associated landward migration of the freshwater–saltwater ecotone within the SESE should increase in response to an acceleration in the rate of SLR. However, the natural hydrology of the SESE was altered by changes in freshwater delivery associated with major drainage projects for wetland reclamation (McVoy et al., 2011). The early, pre-1920 freshwater diversion projects reduced Lake Okeechobee’s stage and the hydrologic head of the Everglades watershed (from +7 m to +3 m), greatly reducing the water storage capacity in the Everglades Ecosystem and ultimately the volume of water moving southward. Increasing population after World War II along the east coast of South Florida was accompanied by an acceleration in surface water diversion activities. Several major drainage projects altering the delivery of freshwater to the SESE were completed by 1920. These reduced Lake Okeechobee stage, water storage capacity in the ecosystem, and ultimately the volume of water moving southward (McVoy et al., 2011). In 1948, as the population increased in South Florida, numerous additional water control construction projects were initiated, including the Water Conservation Areas, the L31W canal, and the South Dade Conveyance System and the C-111 system (Kotun & Renshaw, 2014). In 1968, the South Dade Conveyance System was completed to deliver water from the Water Conservation Areas southward towards eastern Miami-Dade County and Everglades National Park, and ultimately to Biscayne and Florida Bays. In addition, the South Florida Conveyance System drained areas to prevent flooding of agriculture and urban areas. One of the last major features constructed was the L31E storm levee and associated canal along the coast of Biscayne Bay.
These diversions resulted in changes in water delivery to different segments of the coastal wetlands at different times. One of the most obvious signs of changing freshwater delivery are changes in plant communities, sediment types, and loss of agriculture (Parker et al., 1955). Several actions were undertaken to stop or reverse SWE, including the installation of water control structures (Parker et al., 1955; Leach et al., 1972) and the closure of culverts (Craighead, 1966). The Rehydration Pilot Project, undertaken to restore Biscayne Bay coastal wetlands, was initiated in 1994 (Ross et al., 2003). Since 2000, levees were breached and freshwater delivery increased in both the C-111 and L31E canals. The results of these remedial actions are under study. How have these changes in freshwater delivery influenced SWE and ultimately SESE ecosystem response under conditions of historic SLR? If successfully enumerated, these observations will provide a powerful tool to help resource managers optimize ecosystem resilience to predicted SLR. This study was undertaken to distinguish between the effects of SLR and freshwater delivery on SWE in the SESE.
Materials and methods
Experimental design
Our experimental design utilized stratigraphic profiles from five coastal watersheds with different water delivery histories to determine if differences in SWE were detected. Three coastal watersheds along Florida Bay (Taylor Slough, Joe Bay, and Highway Creek) and two along Biscayne Bay (Turkey Point and Mowry Canal) were selected for access reasons or the presence of water level recorders (Fig. 1). Stratigraphic methods were applied to determine SWE rates. If differences in SWE were detected, the relationships between SWE, SLR, and changes in water delivery were examined. The stratigraphic profiles observed in each basin were also used to reconstruct the effects of SWE on SESE depositional environments and associated coastal basin topography.
Study location and field station selection
The SESE varies in width, slope, and water delivery histories and these variations were represented in the five watersheds (Fig. 1). Twenty-six sites were used to establish five transects crossing the salinity gradient in each watershed. Ten sites were located along the shore of Biscayne Bay and 16 sites at long-term water level recorder stations in Everglades National Park. The location of all field stations was determined using GPS (Supplemental Data (SD), Appendix A).
Topography and surface hydrology
Water level recorders mounted on tables with known elevation (hereafter recorder tables) were used for elevation control and acquired from Everglades National Park, National Audubon Society, and U. S. Geological Survey. The elevation of sampling sites without recorder tables was estimated using a 15 cm contour interval map provided by Robert Fennema, Everglades National Park, or determined by interpolation between adjacent stations in areas of negligible slope. Coastal basin topography was delineated by photo documentation and measuring water depth when inundated. Water depths were also measured along each 100 m transect at 5 m intervals and the data only used in basin topography analysis. A Leitz level with accuracy of ±2 mm 100 m−1 was used and each shot closed. All 100 m shots that did not close to within ±2 mm were rejected. Sites along Biscayne Bay were tied to USGS benchmarks. We assumed that control elevations provided to us were correct. Location and distance to the coastline were calculated from GPS positions. Observations of incoming, slack and outgoing tide waters from the ground and helicopter over 20 field days resulted in the documentation of surface water inundation and flow patterns.
Coastal slope
Coastal slope was determined by calculation of elevation differences along the slope (m) and dividing elevation by distance (m) from the coastline to the toe of the Ridge along each transect.
Vegetation
Relative plant taxa abundance was determined by ranking abundance in 20 random 1 m2 plots sampled within a circle with a radius of approximately 50–100 m, centered on each station’s water level recorder when present. Vegetation standing crop was quantified by clipping 10 random 1/4 m2 plots along the 100 m elevation transect at every station (Ross et al., 2000). Standard statistical approaches were used to establish plant communities.
Sediment
The upper 2 cm of sediment from a 10 × 10 cm plot at each vegetation clip site was sampled and analyzed for sediment constituents after removal of surface floc and/or periphyton mat. These data were used to establish the relationship between sediment and plant community, and to characterize extant depositional environments. Sediment cores were collected in 1994 and 1995 at each station using a 7.5 cm diameter aluminum core tube. The core tube was hand pushed to bedrock at the beginning of each undisturbed elevation transect 10 m from the water level recorder. All cores with more than 5% compaction or loss were rejected. The cores were stored in the vertical position and frozen to maintain “in situ” pore water and to facilitate extraction. The sediment was extruded frozen, placed in a miter box, cut into 1 cm thick slabs, placed in pre-labeled Petri dishes, weighed for wet bulk density, and stored in the refrigerator until split for constituent and faunal analysis. Each refrigerated sample was then subdivided into three splits; one for sediment bulk density (dry and wet), one for compositional analysis (Dean, 1974), and the other for paleosalinity analysis as described below. Wet weight was measured immediately after cutting frozen core into 1 cm segments whereas other bulk density measurements were taken from dried (80°C until stable weight was achieved) sub-samples. Organic matter and carbonate content were determined by loss on ignition (LOI). Type of organic constituent (species of origin and constituents such as leaves, flowers, seeds, stems, cable or rhizomes, rootlets or fibrous, and amorphous material) was determined for all surface and core sediment samples using a 10–60 X binocular dissecting scope. Marine-influenced sediments were stained to determine the presence of unstable marine aragonite from stable freshwater low Mg calcite (Freidman, 1959).
Sediment accumulation rates
Sediment accumulation rates were calculated for each of the three major sediment types identified during this investigation; mangrove peat–marl, marl, and sawgrass (Cladium mariscus jamaicense (Crantz) Kük) peat–marl. Cores (12.5 cm in diameter) were collected from two mangrove peat–marl environments in two watersheds, four marl environments in three watersheds, and two sawgrass peat–marl environments in two watersheds (Fig. 1) and both 137Cs and 210Pb methods were utilized (Cutshall et al., 1983; Lynch et al., 1989). The Constant Rate of Supply (CRS) Model was used because cores were long enough to reach baseline Pb levels (Appleby & Oldfield, 1992). Analyses were done at the Wetland Center, Louisiana State University. Cores from Highway Creek and Joe Bay were selected for analysis because both transects have the best core coverage and the Turkey Point core was selected because the extensive marl prairies along Biscayne Bay were not represented. Because of limited funding and the fact that SWE front was past the mangrove peat belt, mangrove peat was only sampled along Biscayne Bay and not analyzed for radioisotopes. Therefore, a fringing and riverine mangrove peat 137Cs analyses were included from the upper Florida Keys for comparison as mangrove peat was a sediment end-member otherwise not represented (Callaway et al., 1997). We assumed that these accumulation rates were analyzed correctly and provided accurate data for the different sediment types.
Depositional environment
The relationship between plant community, sediment type, and sediment accretion rates were determined for each station and stations with similar relationships were defined as depositional environments (Table 3). Depositional environments (DEs) were used in the interpretation of stratigraphy.
Paleosalinity
The third sediment split was washed through a series of sieves (1 and 2 mm diameter) and all skeletal material collected, dried, identified and counted. Mollusks were, by far, the most abundant preserved invertebrate phyla and the salinity ranges for each species was determined from surface sample site salinities and the literature (Ladd, 1957; Tabb & Manning, 1961; Moore, 1964; Turney & Perkins, 1972; Abbott, 1974; Thompson, 1984). A salinity tolerance from 1 (restricted to freshwater environments) to 5 (species restricted to full marine salinity) was assigned to each mollusk species (SD, Appendix B). Paleosalinity was determined using a salinity index (SI) developed for use with diatoms (Blinn, 1993) and previously used with this molluscan assemblage (Meeder et al., 1996; Ross et al., 2002). A weighted averaging procedure was used to calculate sample SI using the equation:
where SIsp was a species SI, and DENSsp and DENSsamp was the density of individual species and all mollusks, respectively, in each sample. SIsamp ranged from 1 to 5, with SI of 1.0 representing freshwater conditions and SI values >1.5 were interpreted as marine-influenced sediments. The SI value of 1.5 was used because the most common gastropod, Littoridinops monroensis (Frauenfeld), was found in both fresh and brackish water and assigned the SI value of 1.5. The SI value was calculated for each individual interval. The marine-influenced freshwater contact in each core was established, and contacts between cores in each transect connected forming the isohaline line. The isohaline line was placed where the SI values remain constant and not at the contact of an individual strata with an SIsamp > 1.5 (All future references to individual SIsamp are indicated by SI without the subscript). Isohaline lines constructed within each stratigraphic profile were drawn through the center of the 1 cm sample interval, which represents a 3–7 year period depending upon sediment type.
Stratigraphy
A stratigraphic approach was used to reconstruct SWE history because the sediments contain a continuous record of salinity changes. The first step was to complete descriptions of sediment cores in each transect and then construct a stratigraphic profile using standard stratigraphic methods. Each stratigraphic profile documented the temporal and spatial relationships between the extant plant community and corresponding sediment type representing an evolving DE along the marine-to-freshwater transition. Timelines for the years 1900, 1940, 1968, and 1995, were then established based upon sediment type and respective accumulation rate. The 1900 timeline was selected because it coincided with the Flagler Railroad construction, which predated other major hydrologic alterations and was young enough to allow sediment accumulation to be quantified using the 210Pb methodology. The other three timelines were chosen to correspond with available complete aerial photography coverage. Most major alterations of water delivery occurred within 5 yrs of the aerial photos used in this study. Four timelines representing the depositional surface were established along all transects. For example, the 1900 timeline was established in a marl sediment by multiplying the interval of time passed between 1900 and 1995, or 95 years, times the average sediment accumulation rate of a marl sediment (1.35 mm year−1). This resulted in the 1900 timeline being drawn 128.3 mm below the present day sediment surface along each transect. The other three timelines were also constructed in this way using accumulation rates associated with each sediment type observed in the stratigraphic section present in each core.
Salt water encroachment
SWE was calculated in five steps: 1. The horizon in each core that denotes a change in salinity from freshwater to marine-influenced sediments was identified based upon calculated SI. 2. An isohaline line was drawn through all horizons along each transect. This line separates freshwater from marine-influenced sediments along each watershed transect. Marine-influenced sediments are found above this isohaline line. 3. This isohaline line was intersected by the timelines. 4. The distance between two successive intersections with timelines was the distance of SWE during that time interval. 5. This calculation was made for the distance between each two successive timelines for each transect and plotted. The elevation at each intersection of the isohaline and timeline was determined by interpolation between two points of known elevation along the transect. The relationship between the elevation of SWE was compared to sea level. The relationship between SWE, plant community change, and the tide stage was established. The error in calculation of SWE by placement of the timeline by either ±1 cm in a 20 cm sediment profile was demonstrated by the application of Pythagorean Theorem. We used the sediment thickness as the right triangle’s opposite side or height (20 cm) and assumed an adjacent side, SWE distance, of 1000 m. We solved for tangent (B) and used the angle reading (89.9885) to calculate changes in SWE by changing sediment thickness. We calculated for sediment thicknesses of 19 and 21 cm which resulted in an error of −54 and +46 m. We felt that this difference, ~0.4%, in SWE over a 20 or 40 years interval was sufficient for our purposes.
Hydrology Alterations
A chronological history of anthropogenic changes in water delivery affecting the SESE was produced from the literature. Project dates that potentially affected water delivery in the SESE were established. The dates of the projects were compared to SWE curves for the different watersheds and projects that coincided with changes in the rate of SWE were considered causative.
Results
Topography and surface hydrology
In general, coastal basins exhibited <40 cm of relief along the otherwise gently sloping shore-normal transects within the SESE (SD, Appendix A). Local relief within a DE exhibited little variation (<10 cm over 100 m; SD, Appendix C). Marl prairies exhibited the least variation in topography except in areas with mangrove clumps which developed small elevated hummocks. Specific observations included: 1. Levees were generally well developed along the coastline and covered with button wood (Conocarpus erectus Linnaeus) or salt marsh grasses along Florida Bay and red mangroves (Rhizophora mangle Linnaeus) along Biscayne Bay, 2. Levees were also present along tidal creeks and their elevation was proportional to tidal range (Fig. 2A), 3. The levee system created a network of coastal depressions or basins (Fig. 3c). Surface water flow during both the rainy and dry season was strongly influenced by topography (Fig. 3c). Tidal ingress into the basins was only observed during spring tides. Freshwater sheet flow filled the basins during the rainy season.

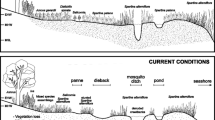
Coastal basin topography is displayed by a an aerial photograph of the coastal area along northern Florida Bay showing the coastal basins outlined by mangrove covered levees (photo by JFM), b an aerial photo displaying a closer view of a tidal creek levee system and the flooded basins to either side (photo by JFM), c a schematic plan view showing both wet (left side) and dry season (right side) both with flow path arrows) conditions and the general basin topography (closed contours) which holds water in the wet season and retains tidal water until evaporation during the dry season. The higher elevation coastal and tidal creek levees are displayed as dark shades, coastal basin periphyton as the medium shade, and sawgrass periphyton as the light shade, and d the distribution of surface sediment types is shown along cross section profiles displaying the coastal and riverine levees constructed of mangrove peat (darkest shades) grading basinward (mangrove peat–marl in dark shade) into marl (medium shade sediments of the basin interior shown inundated (light shade)
Coastal slopes
Coastal slope varied by two orders of magnitude among the five watersheds (Table 1). The coastal slope in Taylor Slough was an order of magnitude greater than the adjacent Joe Bay watershed. Joe Bay and Highway Creek had the lowest slopes. Slopes increased steadily from Joe Bay to Mowry Canal. Coastal width was greatest at Taylor Slough and decreased by >75% to Mowry Canal.
Vegetation
Four major ecoclinal plant communities were identified along the salt-to-freshwater gradient in all watersheds forming shore parallel bands (Fig. 4; SD, Appendix D: Fig. D5). Mangroves were located along the coastline and extend inland as a function of the landward limit of SWE. Mangrove density was greatest along the coast and both tree density and height decreased rapidly with increased distance inland. Mangroves were only minor contributors to biomass except near the coastline. As mangrove abundance decreased periphyton biomass increased. The periphyton community was found throughout the SESE and formed a thick mat of algae, diatoms, and Utricularia spp. and sparse spikerush, Eleocharis cellulosa Torrey (SD, Appendix D: Fig. 3; Appendix E: Fig. E1). The periphyton community also infringed into the sawgrass community reducing the dense sawgrass community to upper Taylor Slough and as small patches at higher interior elevations (SD, Appendix D: Fig. D4). The landward succession of plant communities did not change between watersheds but their width did (SD, Appendix D: Fig. D5). Sawgrass in the Taylor Slough watershed was observed much closer to the coast than in any other watershed and the periphyton prairie was barely present. The periphyton prairie was widest in the Joe Bay and Highway Creek watersheds and all but absent in the Turkey Point and Mowry canal watersheds.

Plant and sediment relationships along the shoreline to interior transition along the Turkey Point transect exhibiting a relative abundance of major plant species. (Mang mangrove, Perip periphyton, Saw sawgrass) and b biomass of major plant species and c Sediment constituents of upper 2 cm
Sediment
Five sediment types were found throughout the study area in shore parallel bands; mangrove peat, mangrove peat–marl, marl, sawgrass peat–marl, and sawgrass peat when present (Fig. 5; SD, Appendix E). Mangrove root material was the third most abundant sediment constituent and characterized both mangrove peat and peat–marl sediment types. The mangrove peat–marl distribution was well into the coastal basins from the natural levees. Periphyton marl prairie was the most common sediment throughout the SESE. Surface marls contained an average of 21% OM and marl samples from cores averaged ~10%, a result of very rapid surface decomposition. The interior border between the extensive marl prairies and the sawgrass peat along the interior edge of the SESE was gradational as marl sediments infiltrate the sawgrass marshes. The interior sawgrass peat was present at higher interior elevations and generally uncommon except in upper Taylor Slough. Although the landward sequence of the four sediment types occurred in the same order, the width of each sediment type band varied and DE 4 was missing from Taylor Slough.

Biogenic nature of SESE sediments is illustrated, arrow width is proportional to constituent abundance
Sediment accumulation rates
Based upon the results of the 210Pb analysis, each sediment type yielded a distinct sediment accumulation rate (Table 2; SD: Appendix F). The 137Cs data were rejected as a consequence of low activity levels, uncharacteristic profiles, or absence of distinct peaks (SD, Appendix F). The highest accumulation rates calculated during this investigation were generated from mangrove peat–marl. These averaged 3.15 mm year−1. The lowest rates were generated from marl samples averaged 1.35 ± 0.21 mm year−1. Intermediate values are associated with sawgrass peat–marl samples and averaged 2.2 mm year−1. Higher organic content was related to higher sediment accumulation rates.
Depositional environments
Six depositional environments (DE) were recognized along the marine-to-freshwater salinity gradient in all watersheds (Table 3). Each DE occurred along shore parallel bands which varied in width between watersheds.
DE 1 The coastal levee differed between Florida and Biscayne Bays. The vegetation and sediments of the Florida Bay levee along Taylor Slough, Joe Bay, and Highway Creek watershed coasts varied considerably in plant cover (from button wood, mangrove, salt marsh grasses, or a combination) and sediment was most frequently a peat with marine mud interbedded. In contrast, the coastal levee of Biscayne Bay along Turkey Point and Mowry Canal basin coasts consisted of red mangrove fibrous peat (DE 1), although the coastal levee along Turkey Point and other locations was nearly completely lost by erosion. Mangroves were found in all plant communities except DE 6 but only contributed significantly to sediment in depositional facies DE 1, DE 2, and DE 3 (Fig. 4). As mangrove relative abundance and biomass decreased with increased distance from the shoreline, the contribution to sediment decreased rapidly (Fig. 4). We assumed that mangrove peat accumulation rate was high (3.9 to 4.2 mm year−1) using Florida Key rates (Table 2).
DE 2 Riverine mangroves were found along the tidal creeks where tidal ingress transported propagules. Mangroves along tidal creeks were taller and denser than the scrub found away from the levees. Propagule settlement developed into a peat-forming environment which resulted in topographic high levee deposits separating coastal basins (Fig. 3). Mangrove levee landward retreat was keeping pace with SWE. Elevation difference between levee and marsh interior decreased with increased distance from the coast to the point of no tidal ingress. Several of our sites were located near tidal levees but none were sampled because they represent a very small fraction of the area. Mangrove-lined creeks were located in all basins and delineate basin boundaries in most cases. In Taylor Slough, the DE 2 has not retreated as far inland along the water course as Joe Bay and Highway Creek which extend inland more than 1 km. Many smaller, shorter tidal creeks which are not freshwater flow-ways do not form basin margins. DE 2 was well developed along Biscayne Bay, especially at Mowry Canal but are rendered functionless because of canal construction.
DE 3 Mangroves decreased in height and percent mangrove cover rapidly away from fringe and riverine mangrove communities producing a mangrove peat–marl (DE 3) that flanked DE 1 and 2. As mangrove cover decreased mangrove tree productivity decreased and root material was replaced by marl production by periphyton between mangrove trees (Fig. 4). Marl was >50%, by weight of mangrove peat–marl. The DE 3 accumulation rate was 3.2 mm year−1 (Table 2). DE 3 was narrowest at Taylor Slough, most extensive in Joe Bay and Highway Creeks and extended ~2.5 km inland at Turkey Point and 400 m in the Mowry Canal watershed.
DE 4 Coastal basins were characterized by low emergent plant cover and a well-developed periphyton mat producing a surface marl sediment with an average of ~21% OM. Scrub mangroves were present in clumps, many decades in age but exhibiting no expansion. The average sediment accumulation rate for DE 4 was 1.35 ± 0.21 mm year−1. DE 4 covered the greatest surface area in all watersheds except for Taylor Slough where it was greatly reduced or missing and Mowry Canal where DE 3 retreated to the toe of the L31E levee.
DE 5 The transitional community between the coastal basin periphyton community and the interior sawgrass peat was covered by low density sawgrass with periphyton producing a sawgrass peat–marl with an accumulation rate of 2.2 mm year−1 (DE 5). DE 5 was an area where SWE was just beginning to change the plant community and mangrove propagule recruitment occurred occasionally with little success. Scrub mangroves were present in places but mangrove material did not contribute to sediment (Fig. 4). In Taylor Slough DE 5 was extensive, extending to within several hundred meters of the coast and DE 3. In Joe Bay and Highway Creek, DE 5 sawgrass peat–marl was not found within 4 km of the coast, ~3.6 km at Turkey Point, and was not present in the Mowry Canal watershed.
DE 6 was dominated by sawgrass with cover >50% forming a sawgrass peat in a freshwater environment. Sediment accumulation was not analyzed in the sawgrass community because of it’s limited distribution. DE 6 was only common in the Taylor Slough interior.
Paleosalinity analysis
In all cases the SI decreased inland and down core (SD, Appendix G: Fig. G2; Appendix H: Table H1). Isohaline lines created from these data (Fig. 6) depict a landward thinning of the stratigraphic section influenced by marine waters. Occasional high SI intervals (<2 cm in thickness) were found in otherwise freshwater intervals. The inland distance of marine-influenced sediments varied between watersheds. The shortest distance of SWE (~2.5 km) was found at Taylor Slough (Fig. 6; Table 2). SWE was ~5 km at Joe Bay and 6.7 km at Highway Creek, nearly all the way to the C-111 canal and levee. SWE was ~3.5 km at Turkey Point and terminated at the L31E levee. In Mowry Canal watershed the distance of SWE was the lowest (0.45 km) and limited by the presence of the L31E levee at 0.65 km.

Transect stratigraphic profiles: the small cross section shows the topography and all core locations along each transect. Mowry Canal transect is short and displayed in total. The larger cross section is an enlarged view of the area of the transect where the isohaline line (dark line) intersects the timelines and the distance between two successive intersections of the timelines (T1, T2, T3, T4) and the isohaline line is the distance of SWE during that time interval
Stratigraphy
The general stratigraphic sequence in all transects was transgressive, marine-influenced sediments overlying freshwater sediments (SD, Appendix I). Mangrove peat–marl was observed overlying marl sediments with minor marine influence. Further inland, marl was observed overlying sawgrass marl. Overstep occurred at Taylor Slough as DE 3 replaced DE 4. The shore-normal length of the transgressive sequence reflects the extent of SWE as described below.
Salt water encroachment
Salt water encroachment occurred in all watersheds as determined from landward penetration of DEs characterized by marine-influenced sediments and salt-tolerant vegetation over freshwater vegetation and associated sediment. The rate of SWE and the elevation of the corresponding isohaline line varied by a factor of 14.8 (Fig. 7). Taylor Slough had the least SWE. Between 1895 and 1940 SWE along the Joe Bay transect was 12 m year−1 and the rate doubled between 1940 and 1968 and again between 1968 and 1994. The elevation of SWE at Joe Bay ended up at the same elevation as high tide (Fig. 7b). Highway Creek transect had the earliest and most rapid rate of SWE between 1895 and 1940 with the highest rate observed between 1940 and 1968, 71.1 m year−1. The rate of SWE declined to the same rate calculated for the interval between 1895 and 1940. The elevation of SWE tracked sea level rise until 1968 when SWE tracked the elevation of high tide. The rate of SWE along the Turkey Point transect was greater than Taylor Slough but lower than Joe Bay and Highway Creek until 1968 when SWE accelerated to 64.8 m year−1, approximately the same as Joe Bay. Turkey Point exhibited the second lowest overall rate of SWE. The elevation of SWE ended up corresponding to the elevation of high tide as in Joe Bay and Highway Creek.

Graphs a displaying the change in distance of SWE over time for each transect (TS Taylor Slough, JB Joe Bay, HC Highway Creek, TP Turkey Point, SL Sea level). Changes in rate of SWE related to the following: 1 Flagler Railroad borrow canal, 2 C1, C102, C103 and Homestead-Ingraham Highway borrow canal, 3 C-111 Canal and Levee, 4 C1, C102, C103 and Homestead-Ingraham Highway borrow canal, 5 C-111 Canal and Levee, 6 L31E Canal and Levee, and 7 insignificant alteration. b Showing the relationship between the elevation of the inflection points between changes in the slope of the encroachment rate curve. The difference between SLR (1) and the elevation of SWE (3) since 1900 is one half the tidal range (2) in this micro-tidal setting
Hydrology alterations
All hydrological alterations of southeast Florida affecting water delivery to the coastal basins were identified in the literature and tabulated by date of occurrence (SD, Appendix J: Table J1, Fig J1). Hydrological alterations include decreased freshwater delivery such as in Taylor Slough, diversion of delivery away from a watershed such as in Joe Bay and Highway Creek, or elimination of sheet flow by construction of barriers such as the L31E at Turkey Point and Mowry Canal watersheds.
Discussion
Topography and surface hydrology
A coastal basin is defined by the area with lower elevation than the coastal levee and extends inland until the levee elevation is reached; this distance is the coastal basin width (Fig. 6). Present coastal basin topography is the result of different depositional environments accumulating sediment at different rates over a gentle sloping limestone bedrock (Fig. 3d). This topography results in extended periods of inundation after tidal (i.e., spring, wind driven) and heavy rainfall (i.e., storm) events (Fig. 3c). Tide stage had to exceed levee elevations (250 to 350 mm above mean sea level) before the basins were flooded. Coastal basin topography varied among watersheds related to extent and duration of SWE and ultimately by the changing rate of SLR. Coastal basin width, as distance along the transect, was shortest in Taylor Slough along Florida Bay ~2.5 km (Fig. 6). Coastal width in Joe Bay was ~8 km nearly the same as Highway Creek ~8.5 km. Both of these basins have the most extensive basin development associated with the earliest SWE. Along the steep coastal slope of Turkey Point the coastal basin was ~800 m in width and at the Mowry Canal ~600 m, terminating at the L31E levee.
The greater the sediment accumulation deficit in respect to SLR the faster coastal basin development occurs. The increase in the rate of SLR from <1 to 2.4 mm year−1 by the end of the study resulted in the basin morphology. The present rate of SLR is between 3.4 and 9 mm year−1 and may increase to 20 mm year−1 within a century (Fig. 2) producing a much greater sediment deficit resulting in predicted further coastal basin deepening and expansion until submergence creating estuarine conditions.
Coastal slope
Coastal slopes are not a function of coastal width. The Taylor Slough slope is an order of magnitude greater than the adjacent Joe Bay although Taylor Slough has the greatest coastal width. The reduced slope in Taylor Slough is associated with sufficient freshwater delivery to support sawgrass closer to the coast. Joe Bay and Highway Creek have the lowest, very similar coastal slope (~8.5 × 10−6) and width (~17.6 km) and both experienced the greatest distance of SWE as expected based upon low slope. However, the greatest rate of SWE is reported in Highway Creek watershed. The slope increases at Turkey Point as the coastal zone width decreases. With increasing coastal slope, less SWE was expected but that was not observed. These two cases clearly indicate other factors, such as freshwater delivery, influence SWE. The same is true for the Mowry Canal which has the greatest slope, least coastal width, and maximum SWE which terminated at the L31E canal.
Plant community and sediment-type relationships
Sediment types throughout the SESE are biogenic in origin corresponding to the plant community (Fig. 5) and are found in shore-normal bands along the marine-to-freshwater gradient; fringe mangroves are associated with peat sediment, mangrove periphyton with mangrove peat–marl, periphyton prairie with marl, sawgrass periphyton with sawgrass peat–marl, and sawgrass with sawgrass peat sediment. The width of the bands varied between watersheds. The association between plant community and sediment type is documented in the SESE (SD, Appendix K). This classification is very similar to the four plant communities previously utilized (Ross et al., 2000). The relationship between plant community and sediment type is well established for southern Florida (Davis, 1946; Jones, 1948; Gleason, 1972; Cohen & Spackman, 1974; Gleason & Spackman, 1974; Gleason et al., 1974; Duever et al., 1986; Gleason & Stone, 1994). Mangrove presence and associated sediment types decreased in abundance with increased distance from the coast (SD, Appendices D, E, K). The lack of mangrove above ground material documented the significance of mangrove root penetration in sediment production (McIvor et al., 2013) and we assume that root penetration is a continuous process in mangrove peat environments (Castañeda-Moya et al., 2011). Root expansion was found to occur at different rates in different environmental setting (Whelan et al., 2005) and we assume that root processes work the same in different DEs. Mangroves retreated much further inland than mangrove material is found in sediments, suggesting a time lag between mangrove settlement and peat production which is decades in duration. In this region where mangroves invaded periphyton prairie, the associated marl sediment is being replaced by mangrove peat–marl. Unlike roots in the peat sediment profiles scrub mangrove roots run horizontally, usually providing a distinct contact with underlying marl (SD, Appendix I).
Periphyton and the resultant marl sediment is the most widespread plant community and sediment type. The lowest diversity and percent cover of emergent vegetation are found in the periphyton prairie community which is very reflective of light showing up as a white band just interior of the mangroves in aerial photography, this band is commonly referred to as a “White Zone” within the regional scientific community (Ross et al., 2002). The marl prairie retreats with SWE eventually replacing sawgrass peat with sawgrass peat–marl as documented by stratigraphy. This sawgrass peat–marl presently has the second largest distribution. Sawgrass abundance increases with increased distance from the coast (SD, Appendix D: Fig. D4). Most of the sawgrass peat was replaced by sawgrass peat–marl throughout the SESE.
The major differences among watersheds are related to the distance of mangrove and periphyton community retreat which are both related to the distance of SWE. In Taylor Slough mangroves are found 12.5 km into the interior but only contributed to sediments for a distance of 2.5 km. The marl prairie is not observed along the Taylor Slough transect because of overstep by mangrove retreat. Sawgrass, in contrast to mangroves, is found within 1 km of the coast but only contributed to sediments at a distance of ~2.5 km from the coast. Mangroves in both Joe Bay and Highway Creek are found >7.5 km from the coast but mangroves only contributed to Joe Bay sediments at a distance of ~4 km from the coast whereas mangrove contribution to sediments in Highway Creek corresponds to the distribution of mangroves. The more abundant mangrove material in the sediments in Highway Creek is the result of longer duration of mangrove presence because of earlier SWE. Sawgrass at Joe Bay and Highway Creek is found within 2.5 km of the coast but made contribution to sediments to a distance of 4 km at Joe Bay and no closer to the coast than 7.5 km at Highway Creek, also a result of SWE history. Mangroves are present in the Turkey Point basin to a distance of 4.5 km from the coast but only contributed to sediments at a distance of 750 m in response to rapid SWE in the last 30 years, not a long enough period for mangrove contribution to sediment. Sawgrass presence is found to be within 1.7 km of the coast but did not contribute to sediments along the 4.5 km transect. The lack of sawgrass contribution to the sediment is the result of the dominance of periphyton along this transect for the period of study. Sawgrass is not found at Mowry. The variation found in the distribution of mangrove, marl, and sawgrass among basins is associated with SWE as no other factors can explain observations.
Episodic deposition did not significantly alter the observed patterns of sediments within the study area although the region is prone to hurricane landfall (Duever et al., 1994). The sedimentological consequences of hurricane landfall were limited by the presence of shallow water bodies (Biscayne and Florida Bays), outer islands, the Florida Keys, and the Buttonwood Ridge that dampen storm surge and wave climate (Davis et al., 2004; Castañeda-Moya et al., 2010). Historical landfalls in this region are described (Craighead & Gilbert, 1962; Craighead, 1964; Whelan et al., 2009; Castañeda-Moya et al., 2010; Vlaswinkel & Wanless, 2012) but we found only one recognizable storm deposit and no depositional hiatuses that are interpreted as erosional surfaces. The observed marine mud storm layer deposited during Hurricane Andrew along the Turkey Point transect is no longer observable. The disappearance of storm deposits has been observed by others working in Florida as well (Harold R. Wanless, University of Miami, Personal communication). At least ~10 m of coastal mangrove fringe was eroded between 1968 and 1995 based upon submerged mangrove peat overlying bedrock.
Sediment accumulation rates
Sediment accumulation rates are different for each sediment type and are assumed to be applicable throughout each DE where they occur. Limited variation in sediment-type accumulation data suggests that the assumption is appropriate. In addition, the parameters controlling sediment accretion rate within a DE are the same such as plant community, soil type, hydroperiod, and exposure to salt water because of sulfur and phosphorous addition (Krairapanond et al., 1991; Dévai et al., 1996; DeLaune et al., 2002; Chambers et al., 2015). Post-depositional processes are not complete in these young sediments that affect long-term changes in elevation and therefore net accumulation (Parkinson et al., 1994). Post-depositional processes are found to vary with environment and include decomposition (Alongi et al., 2000; Poret et al., 2007; Huxham et al., 2010), compaction (Middleton & McKee, 2001), and surface subsidence (Cahoon & Lynch, 1997). However, these are poorly constrained within the SESE and were assumed to exert a uniform influence on the sediment succession within each depositional setting. The sediment accumulation calculations have not considered processes that have not occurred to completion and therefore are conservative estimates of SWE because on the decadal to century time scale SWE should increase with decreased surface elevation.
Depositional environments
The six DEs are found in each watershed in shore-normal bands in the same pattern along the marine-to-freshwater gradient. The distribution of these DEs should be the same if SLR and freshwater delivery were constant. However, distribution varies between watersheds. Therefore, changes in water delivery must be responsible for differences in DE distribution because the rate of SLR is the same for all watersheds. The other contributing factors are coastal slope and width which do not affect SWE uniformly among watersheds and therefore cannot explain observed differences in DE distribution among basins.
DE 1 is adjacent to open marine waters and is therefore the most exposed to salinity, sulfur, phosphorous, wave, and tidal energy. Mangroves are not affected by changing salinities under normal salinity ranges (Odum et al., 1982). Therefore, SWE only affects DE 1 by the associated SLR. Under conditions of this study DE 1 (accretion rate of 4.1 mm year−1) has maintained its relative position with the rate of SLR (2.4 mm year−1) as did the other plant species found on coastal levees. Turkey Point was the only basin to have lost DE 1 as a result of erosion. The rate of SLR is accelerating and expected to reach 9–20 mm year−1 well within this century (Fig. 2). At these rates under the local depositional regime it is doubtful that DE 1 will maintain its position because of submergence or erosion. That the rate of SLR controls the rate of mangrove peat accumulation has been postulated (McKee et al., 2007; McIvor et al., 2013); however, in this oligotrophic region with no allochthonous sediment influx it is most likely that DE 1 will submerge or erode at higher rates of SLR than during the study interval.
The mangroves of DE 2 are maintaining their relationship in respect to SLR while retreating along tidal creeks with increased tidal ingress by propagule recruitment. Mangrove along tidal creeks are taller and denser than the scrub found away from the levees, presumably because of nutrients associated with tidal ingress and freshwater delivery. Retreating mangroves are settling on nutrient-poor marl sediments with dense periphyton cover perhaps adversely affecting success and productivity. Mangroves of DE 2 decrease in height towards the interior and are reduced to a narrow, discontinuous band as the extent of tidal ingress is approached in all watersheds. In some watercourses mangroves are competing with freshwater herbaceous vegetation (Ross et al., 2002). The peat along the lower reach of the tidal creeks is gradually replaced upstream by mangrove peat–marl and in the region of greatest tidal extent marl sediment covered with young mangroves. The distance of tidal creek levee development is related to the distance of SWE as recorded (Fig. 5).
Mangroves of DE 3 (producing an average sediment accumulation rate of 3.2 mm year−1) expanded over sediments of DE 4 (1.2 mm year−1) in all watersheds from their original distribution along the coasts and creeks, forming landward-thinning transgressive stratigraphic sequences (SD, Appendix H). Mangroves of DE 3 are exposed to much less tidal action and longer periods of inundation, especially during rainy seasons than DE 1 or 2. Mangroves expanded into coastal basins but lag behind SWE. At Taylor Slough DE 3 overstepped the marl soils of the periphyton prairie and is retreating into the sawgrass community (DE 5). Mangrove retreat in Joe Bay and Highway Creek circumvented the deepening coastal basins, too deep for propagule settlement on the periphyton-covered marl surface (Odum et al., 1982; Koch, 1997; Chen & Twilley, 1999; Krauss et al., 2008) and moved inland into sparse sawgrass (DE 5). The retreat of mangroves into DE 5 is likely the first sign of overstep in these two watersheds. DE 3 is not well developed at Turkey Point as tidal creeks are very small with no water delivery because of the L31E levee or at Mowry Canal where canals and levees have replaced the creeks.
DE 4 (the White Zone) is covered by sparse emergent vegetation and a periphyton mat that is often >10 cm thick in all basins (SD, Appendix D: Fig. D3; Appendix E: Table 1). DE 4 is found in coastal basins which experience the longest hydroperiod and greatest salinity fluctuation. The marl sediments of DE 4 have the lowest accumulation rates of the sediment types (Table 2). The increasing marl accumulation deficit in respect to the rate of SLR explains coastal basin development by deepening and landward expansion, following the leading edge of the white zone (Ross et al., 2002). Much of DE 4 in Joe Bay and Highway Creek coastal basins is too deep for mangrove colonization. However, previously settled mangroves have formed small, dense clumps, the bases of which have accumulated sediment much faster than the surrounding marl sediment forming a hummocky topography on the otherwise very low relief marl surface. These mangrove clumps are decades old based upon aerial photographs but exhibit no evidence of increased cover. DE 4 retreated to the levee at Turkey Point and no longer exists at Mowry Canal. As the rate of SLR increases DE 4 sediment deficit in coastal basins will increase resulting in submergence leaving only mangrove levee or clumps of mangroves around their margins. We suggest that these coastal basins will become new estuarine bodies like those found along the present Florida Bay coastline.
DE 5, the sawgrass peat–marl, developed over a precursor sawgrass peat that was invaded by periphyton and Eleocharis as SWE deteriorated sawgrass habitat. This environment is dominantly a freshwater environment that undergoes frequent SWE events sufficient to reduce sawgrass recovery. The boundary between DE 5 and DE 6 is the leading edge of SWE. At Taylor Slough DE 3 retreated directly into DE 5 because DE 4 could not compete or retreat into the faster sediment accreting sawgrass community, maintained by freshwater delivery. DE 5 at Joe Bay and Highway Creek was a very wide band which was expanding landward, while being replaced by marl (DE 4) and expanding basin on the coastal side. The typical sediment profile along this leading edge of SWE is sawgrass peat–marl overlying sawgrass peat, in contrast to, mangrove peat–marl overlying marl in sediment cores from closer to the coast (SD, Appendix I).
DE 6 is not common at the surface in the study area except in upper Taylor Slough watershed along the main flow-way and as small patches along the Ridge. However, sawgrass peat is found in the subsurface at other sites documenting the loss of dense sawgrass cover during the last century from SWE.
DEs are found in shore-normal bands because different plant communities respond to salinity differently. However, the widths of bands vary between watersheds in response to different rates and periods of SWE. Differences in SWE among watersheds is related to differences in water delivery because the rate of SLR was the same throughout the study area and differences in physical parameters were not consistent with SWE rates. Each DE comprised a unique plant community and sediment type that developed under a set of distinct environmental conditions such as salinity, hydroperiod, or nutrient regime. In the oligotrophic, extremely low relief, micro-tidal SESE with insignificant allochthonous sediment influx environmental controls of sediment accumulation in different DEs are assumed to affect post-depositional processes uniformly within a DE.
Paleosalinity analysis
The freshwater molluscan assemblage is very different and easily recognized from marine mollusks and are a good proxy for paleosalinity. Changes in SI are associated with changes in plant community and sediment type as documented. Small reversals in SI values in a core sequence of less than two cm are most likely the result of drought or storm deposition as the shells were often leached or broken and are only found close to the coast. Such short intervals of higher SI values are not used to determine placement of the isohaline lines.
Stratigraphy
All transects record transgressive sequence stratigraphy but the distance of the transgression varied among watersheds (Fig. 6; SD, Appendix I). In the majority of cores mangrove peat–marl is found overlying marl or marl overlying sawgrass peat or sawgrass peat–marl. Taylor Slough is the only deviation, as DE 4 is overstepped by DE 3 (SD, Appendix I, Fig. 1). This overstep was the result of sufficient water delivery to maintain sawgrass preventing retreat of the periphyton marl prairie and rapid mangrove retreat replacing the periphyton marl prairie. The longest transgressive sequences are found along the Joe Bay and Highway Creek transects both with very similar slopes and associated with the greatest SWE. However, DE 3 retreat was only 7.5 km in Joe Bay in contrast to >10 km in Highway Creek a result of more extensive SWE for a longer period of time in Highway Creek. The transgressive sequences at the Turkey Point and Mowry Canal are incomplete as transgression is terminated at the toe of the L31E levee. The distance of SWE controlled the length of the transgressive sequence. SWE and overstep have occurred at a SLR rate of 2.4 mm year−1 with a marl sediment deficit in respect to SLR of only 1.3 mm year−1. Under conditions of present and future rates of SLR of >9 mm year−1, overstep and submergence are expected throughout the SESE.
Salt water encroachment and hydrological alterations
SWE occurred in each watershed but at different rates among watersheds and time intervals within watersheds (Fig. 7a). These different SWE rates occurred during the same period of SLR (~20 cm) and therefore document that water delivery affected the rate of SWE both temporally and spatially. Taylor Slough had the lowest rate of SWE, the shortest distance of landward SWE, and sawgrass was found closest to the coast because of freshwater delivery. This does not suggest that water delivery was not reduced or altered historically but that there was sufficient delivery to eliminate the tidal signal (Fig. 7b) and maintain sawgrass at the SLR rate during the study. Freshwater delivery into Taylor Slough was sufficient to prevent 37% of the SWE experienced in the adjacent Joe Bay. The present rate of SLR is approximating or has passed the rate of sawgrass peat–marl accumulation which suggests that a sediment accumulation deficit should develop for DE 5, as well as, DE 4 increasing SWE in Taylor Slough.
Between 1895 and 1940 the rate of SWE at Joe Bay was 12.1 m year−1, approximately twice that of adjacent Taylor Slough, and coincides with pirating of water to the east by the Flagler Railroad borrow canal. SWE continued to increase because of additional water diversion associated with the C-111 canal and levee but at a slower rate than the adjacent Highway Creek with the same coastal slope and width. The difference in SWE between these two very similar basins can only be the result of different water delivery histories. SWE in both Joe Bay and Highway Creek reached the elevation of high tide (Fig. 7b). SWE at both Turkey Point and Mowry Canal was obviously associated with the construction of the L31E levee and SWE reached the elevation of high tide. The different rates of SWE among basins can only be explained by different water delivery histories. That the elevation of SWE reached the elevation of high tide in 4 out of 5 watersheds documents that sea level ultimately controls SWE.
SWE at Joe Bay and Highway Creek reached one half the distance to the Ridge in the last century at the SLR rate of 2.4 mm year−1. At present or future predicted rates of SLR SWE should reach the Ridge within decades and present coastal basins will become submerged and DEs overstepped. Delivery of additional water to counter future SLR is unlikely to succeed because at higher sea levels the residence time of freshwater is reduced and water resources are declining because of urban demand. Available water for delivery is already reduced by 70% (Perry, 2004) and restoration plans to increase delivery by use of recycled and ground water is postulated (Browder et al., 2005) with questionable success. However, a considerable increase in delivery is required to return conditions to 1900 and under conditions of increasing rate of SLR it’s doubtful that enough water is available to stop SWE and submergence.
Conclusion
Plant communities and associated sediment types have distributions that are shore normal along a salt-to-freshwater gradient based upon salinity tolerance: mangrove, periphyton-spike rush, and sawgrass. SWE creates landward shifts of plant communities resulting in contemporaneous shifts in associated sediment types. The relationship between plant communities and sediment types is demonstrated in this biogenic depositional setting with no allochthonous sediment influx.
Sea level rise is the primary cause of SWE although changes in freshwater delivery can influence the rate of SWE. This conclusion is documented by different rates of SWE in different watersheds occurring during the same condition of SLR. The elevation of SWE in Taylor Slough tracked rising mean sea level during the last century although the only watershed documenting overstep, the loss of the periphyton community by mangrove retreat. In contrast, SWE in the other watersheds reached the elevation of high tide documenting that sufficient water delivery existed to remove the tidal signl in Taylor Slough. Freshwater delivery reduced SWE in Taylor Slough by maintaining faster sediment accreting sawgrass community which maintained surface elevation in respect to sea level.
The rates of SWE and the elevation of the SWE front were produced under accelerating conditions of SLR, from <1 to 2.4 mm year−1 or ~20 cm during the previous century. This SWE was accompanied by local coastal erosion at the Turkey Point transect. The present rate of SLR in the region is between 3.4 and 9 mm year−1 and may reach as much as 20 mm year−1 within the next century. At present and greater rates of SLR, the SESE is expected to experience continued, more rapid SWE to the Ridge and continued inundation until submergence resulting in much of the SESE becoming estuarine in nature. The results of this study provide managers with an enhanced understanding of how the SESE will respond to future sea level rise.
References
Abbott, R. T., 1974. American Seashells: The Marine Molluska of the Atlantic and Pacific Coasts of North America, 2nd ed. Van Nostrand Reinhold, New York.
Alongi, D. M., F. Tirendi & B. F. Clough, 2000. Below-ground decomposition of organic matter in forests of the mangroves Rhizophorastylosa and Avicenniamarina along the arid coast of Western Australia. Aquatic Botany 68: 97–122.
Appleby, P. G. & F. Oldfield, 1992. Applications of lead-210 to sedimentation studies. In Ivanovich, M. & R. S. Harmon (eds), Uranium-series disequilibrium: applications to earth, marine, and environmental sciences, 2nd ed. Clarendon Press, Oxford: 732–783.
Blinn, D. W., 1993. Diatom community structure along physiochemical gradients in saline lakes. Ecology 74: 1246–1263.
Browder, J. A., R. Alleman, S. Markley, P. Ortner & P. A. Pitts, 2005. Biscayne Bay conceptual ecological model. Wetlands 25: 854–869.
Cahoon, D. R. & J. C. Lynch, 1997. Vertical accretion and shallow subsidence in a mangrove forest of southwestern Florida, USA. Mangroves and Salt Marshes 1: 173–186.
Callaway, J. C., R. D. DeLaune & W. H. Patrick Jr., 1997. Sediment accumulation rates from four coastal wetlands along the Gulf of Mexico. Journal of Coastal Research 13: 181–191.
Castañeda-Moya, E., R. R. Twilley, V. H. Rivera-Monroy, K. Zhang, S. E. Davis & M. Ross, 2010. Sediment and nutrient deposition associated with Hurricane Wilma in mangroves of the Florida Coastal Everglades. Estuaries and Coasts 33: 45–58.
Castañeda-Moya, E., R. R. Twilley, V. H. Rivera-Monroy, B. D. Marx, C. Coronado-Molina & S. M. Ewe, 2011. Patterns of root dynamics in mangrove forests along environmental gradients in the Florida Coastal Everglades, USA. Ecosystems 14: 1178–1195.
Chambers, L. G., S. E. Davis, T. G. Troxler & J. A. Entry, 2015. Sea level rise in the Everglades: plant-soil-microbial feedbacks in response to changing physical conditions. In Entry, J. A., A. D. Gottlieb, K. Jayachandran & A. Ogram (eds), Microbiology of the Everglades Ecosystem. CRC Press, Boca Raton: 89–112.
Chen, R. & R. R. Twilley, 1999. Patterns of mangrove forest structure and soil nutrient dynamics along the Shark River estuary, Florida. Estuaries 22: 955–970.
Church, J. A. & N. J. White, 2011. Sea-level rise from the late 19th to the early 21st century. Surveys in Geophysics 32: 585–602.
Cohen, A. D. & W. Spackman, 1974. The phytogenic organic sediments and sedimentary environments in the Everglades-mangrove complex. Part II. Palaentographica 162: 71–114.
Craighead Jr., F. C., 1964. Land, mangroves and hurricanes. Fairchild Tropical Garden, Miami.
Craighead Jr., F. C., 1966. Additional considerations of the experimental closing of the culverts along the Flamingo Road. Technical Report, National Park Service.
Craighead Jr., F. C. & V. C. Gilbert, 1962. The effects of Hurricane Donna on the vegetation of southern Florida. Quarterly Journal of the Florida Academy of Sciences 25: 1–28.
Cutshall, J. C., I. R. Larsen & C. R. Olsen, 1983. Direct analysis of Pb210 in sediment samples: self-absorption corrections. Nuclear Instruments and Methods 206L: 309–312.
Davis, J. H., Jr., 1940. The ecology and geologic role of mangroves in Florida. Papers Tortugas Laboratory, Carnegie Institute, Washington, Publication No. 517: 304–412.
Davis Jr., J. H., 1946. The peat deposits if Florida. Florida Geological Survey Bulletin 25: 1–311.
Davis III, S. E., J. E. Cable, D. L. Childers, C. Coronado-Molina, J. W. Day Jr., C. D. Hittle, C. J. Madden, E. Reyes, D. Rudnick & F. Sklar, 2004. Importance of storm events in controlling ecosystem structure and function in a Florida gulf coast estuary. Journal of Coastal Research 20: 1198–1208.
Dean Jr., W. E., 1974. Determination of carbonate and organic matter in calcareous sediments and sedimentary rocks by loss on ignition: comparison and other methods. Journal Sedimentary Petrology 44: 242–248.
DeLaune, R. D., I. Devai, C. R. Crozier & P. Kelle, 2002. Sulfate reduction in Louisiana marsh soils of varying salinities. Communications in soil science and plant analysis 33: 79–94.
Dévai, I. S. T. V. Á. N., K. R. Reddy, R. D. DeLaune & D. A. Graetz, 1996. Sulfate reduction and organic matter decomposition in a wetland soil and lake sediment. Acta Biologica Debrecina Supplementum Ecologica Hungarica 6: 13–23.
Duever, M. J., J. E. Carlson, J. F. Meeder, L. C. Duever, L. H. Gunderson, L. A. Riopelle, T. R. Alexander, T. R. Myers & D. P. Spangler, 1986. The big cypress national preserve. National Audubon Society Research Report 8: 1–455.
Duever, M. J., J. F. Meeder, L. B. Meeder & J. M. McCollom, 1994. The climate of South Florida and its role in shaping the Everglades Ecosystem. In Davis, S. M. & J. C. Ogden (eds), Everglades: The Ecosystem and its Restoration. St. Lucie Press, Delray Beach: 225–248.
Ellison, J. C., 1993. Mangrove retreat with rising sea-level, Bermuda. Estuarine, Coastal and Shelf Science 37: 75–87.
Ellison, J. C. & D. R. Stoddart, 1991. Mangrove ecosystem collapse during predicted sea-level rise: Holocene analogues and implications. Journal of Coastal Research 7: 151–165.
Egler, F. E., 1952. Southeast saline Everglades vegetation, Florida, and its management. Plant Ecology 3: 213–226.
Ezer, T., 2013. Sea level rise, spatially uneven and temporally unsteady: why the US East Coast, the global tide gauge record, and the global altimeter data show different trends. Geophysical Research Letters 40: 5439–5444.
Friedman, G. M., 1959. Identification of carbonate minerals by staining methods. Journal of Sedimentary Research 29: 87–97.
Gleason, P., 1972. The origin, sedimentation, and stratigraphy of a calcitic mud located in the southern fresh-water Everglades. Ph.D. Dissertation, Pennsylvania State University, University Park.
Gleason, P. J. & P. A. Stone, 1994. Age, origin, and landscape evolution of the Everglades peatland. In Davies, S. M. & J. C. Ogden (eds), Everglades: The Ecosystem and its Restoration. St Lucie Press, Delray Beach: 149–198.
Gleason, P. J. & W. Spackman, Jr., 1974. Calcareous periphyton and water chemistry in the Everglades. In Gleason, P.J. (ed.), Environments of South Florida: Present and Past. Miami Geological Society Memoir 2: 146–181.
Gleason, P. J., A. D. Cohen, W. G. Smith, H. K. Brooks, P.A. Stone, R. L. Goodrick & W. Spackman, Jr., 1974. The environmental significance of Holocene sediments from the Everglades and saline tidal plain. In Gleason, P. J. (ed.), Environments of South Florida: Present and Past. Miami Geological Society Memoir 2:287–341.
Hay, C. C., E. Morrow, R. E. Kopp & J. X. Mitrovica, 2015. Probabilistic reanalysis of twentieth-century sea-level rise. Nature Climate Change 517: 481–484.
Huxham, M., J. Langat, F. Tamooh, H. Kennedy, M. Mencuccini, M. W. Skov & J. Kairo, 2010. Decomposition of mangrove roots: effects of location, nutrients, species identity and mix in a Kenyan forest. Estuarine, Coastal and Shelf Science 88: 135–142.
IPCC, 2013. Summary for policymakers. In Stocker T, Qin D et al. (eds). Climate change 2013: the physical science basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA [available on internet at https://www.ipcc.ch/report/ar5/wg1/]. Accessed 20 Feb 2014.
Jones, L. A., 1948. Soils, geology, and water control in the Everglades region. University of Florida Agricultural Experimental Station Bulletin 442: 1–168.
Koch, M. S., 1997. Rhizophora mangle L. Seedling Development into the Sapling Stage across Resource and Stress Gradients in Subtropical Florida. Biotropica 29: 427–439.
Kotun, K. & A. Renshaw, 2014. Taylor Slough hydrology. Wetlands 34: 9–22.
Krairapanond, N., R. D. DeLaune & W. H. Patrick, 1991. Seasonal distribution of sulfur fractions in Louisiana salt marsh soils. Estuaries and Coasts 14: 17–28.
Krauss, K. W., C. E. Lovelock, K. L. McKee, L. López-Hoffman, S. M. Ewe & W. P. Sousa, 2008. Environmental drivers in mangrove establishment and early development: a review. Aquatic Botany 89: 105–127.
Ladd, H. S., 1957. Paleoecological evidence. In Ladd, H. S. (ed.), A treatise on marine Ecology and paleoecology. Geological Society of America Memoirs 67: 599–640.
Leach, S. D., H. Klein & E. R. Hampton, 1972. Hydrologic effects of water control and management of southeastern Florida. Florida Bureau of Geology, Report of Investigations 60: 1–115.
Lynch, J. C., J. R. Meriwether, B. A. McKee, F. Ver-Herrera & R. R. Twiley, 1989. Recent accretion in mangrove ecosystems based on Cs137 and Pb210. Estuaries 12: 284–299.
Maul, G. A. & D. M. Martin, 1993. Sea level rise at Key West, Florida, 1846–1992: America’s longest instrument record? Geophysical Research Letters 20: 1955–1958.
McIvor, A. L., T. Spencer, I. Möller & M. Spalding, 2013. The response of mangrove sediment surface elevation to SLR. The Nature Conservancy and Wetlands International, Natural Coastal Protection Series: Report 3. Cambridge Coastal Research Unit Working Paper 42: 1–59.
McKee, K. L., D. R. Cahoon & I. C. Feller, 2007. Caribbean mangroves adjust to rising sea level through biotic controls on change in soil elevation. Global Ecology and Biogeography 16(5): 545–556.
McVoy, C. W., W. P. Said, J. Obeysekera, J. A. Van Arman & T. W. Dreschel, 2011. Landscapes and hydrology of the predrainage Everglades. University Press of Florida, Gainesville.
Meeder, J. F., M. S. Ross, G. Telesnicki, P. L. Ruiz, & J. P. Sah, 1996. Vegetation Analysis in the C-111/Taylor Slough Basin. In two parts: (1) Document One, The Southeast Saline Everglades revisited: a half century of coastal vegetation change; (2) Document Two, Marine Transgression in the southeast Saline Everglades, Southeast Environmental Research Center Research Report 6 [available on internet at http://digitalcommons.fiu.edu/sercrp/6].
Middleton, B. A. & K. L. McKee, 2001. Degradation of mangrove tissues and implications for peat formation in Belizean island forests. Journal of Ecology 89: 818–828.
Moore, D. R., 1964. Mollusca of the Mississippi coastal waters. Gulf Coast Bulletin 1.
NASA, 2016. Global Climate Change: Vital Signs of the Planet [available on internet at http://climate.nasa.gov/vital-signs/sealevel/].
Nicholls, R. J. & A. Cazenave, 2010. Sea-Level Rise and Its Impact on Coastal Zones. Science 328: 1517–1520.
NOAA, 2012. CO2 data of US National Oceanic and Atmospheric Administration [available on internet at ftp://ftp.cmdl.noaa.gov/ccg/co2/trends/co2_annmean_gl.txt].
Odum, W. E., C. C. McIvor & T. J. Smith, III, 1982. The ecology of the mangroves of South Florida: A community profile. FWS/OBS-81/24: 1–145.
Park, J. & W. V. Sweet, 2015. Accelerated sea level rise and Florida Current transport. Ocean Science 11: 607–615.
Parker, G. G., G. E. Ferguson & S. K. Love, 1955. Water resources of southeastern Florida with special reference to the geology and ground water of the Miami area. United States Geological Survey Water-Supply Paper 1255: 1–965.
Parkinson, R. W., 1989. Decelerating Holocene SLR and its influence on southwest Florida coastal evolution: a transgressive-regressive stratigraphy. Journal of Sedimentary Petrology 59: 960–972.
Parkinson, R. W. & J. F. Meeder, 1991. Mud-bank destruction and the formation of a transgressive sand sheet, southwest Florida inner shelf. Geology Society of America Bulletin 103: 1543–1551.
Parkinson, R. W., R. D. DeLaune & J. R. White, 1994. Holocene sea-level rise and the fate of mangrove forests within the wider Caribbean region. Journal of Coastal Research 10: 1077–1086.
Parkinson, R. W., P. W. Harlem & J. F. Meeder, 2015. Managing the Anthropocene Marine Transgression to the 2100 and beyond in the State of Florida USA. Climatic Change 128: 83–98.
Perry, W., 2004. Elements of south Florida’s comprehensive Everglades restoration plan. Ecotoxicology 13: 185–193.
Poret, N., R. R. Twilley, V. H. Rivera-Monroy & C. Coronado-Molina, 2007. Belowground decomposition of mangrove roots in Florida coastal Everglades. Estuaries and Coasts 30: 491–496.
Rohling, E. J., I. D. Haigh, G. L. Foster, A. P. Roberts & K. M. Grant, 2013. A geological perspective on potential future sea-level rise. Scientific Reports 3. doi:10.1038/srep03461.
Ross, M. S., J. F. Meeder, J. P. Sah, P. L. Ruiz & G. J. Telesnicki, 2000. The southeast saline Everglades revisited: 50 years of coastal vegetation change. Journal of Vegetation Science 11: 101–112.
Ross, M. S., E. E. Gaiser, J. F. Meeder & M. T. Lewin, 2002. Multi-taxon analysis of the “white zone”, a common ecotonal feature of South Florida coastal wetlands. In Porter, J. W. & K. G. Porter (eds), The Everglades, Florida Bay and Coral Reefs of the Florida Keys. CRC Press, Boca Raton: 205–238.
Ross, M. S., J. F. Meeder, E. Gaiser, P. L. Ruiz, J. P. Sah, D. L. Reed, J. Walters, G. T. Atlas, A. Telesnicki, M. Wachnicka, J. Jacobson, J. Alvord, M. Byrnes, C. Weekley, M. T. Lewin, B. Fry & A. Renshaw, 2003. The L-31E Surface Water Rediversion Project Final Report: Implementation, Results, and Recommendations. Southeast Environmental Research Center Research Report 1 [available on internet at http://www.digitalcommons.fiu.edu/sercrp/1].
Scholl, D. W., F. C. Craighead & M. Stuiver, 1969. Florida submergence curve revised: its relation to coastal sedimentation rates. Science 163: 562–564.
Spackman, W., A. D. Cohen, P. H. Given & D. J. Casagrande, 1976. A field guidebook to aid in the comparative study of the Okefenokee Swamp and the Everglades-mangrove swamp-marsh complex of southern Florida. A short course presentation, June 1976, Coal Research Section, The Pennsylvania State University (No. 6).
Tabb, D. C. & R. B. Manning, 1961. A checklist of the flora and fauna of northern Florida Bay and adjacent brackish waters of the Florida mainland collected during the period July 1957 through September 1960. Bulletin Marine Science 1: 550–647.
Thompson, F. G., 1984. Freshwater Snails of Florida: A Manual for Identification. University of Florida Press, Gainesville, FL.
Turney, W. J. & B. F. Perkins, 1972. Molluscan distribution in Florida Bay. Comparative Sedimentology Laboratory, Sedimenta III: 1–37.
Vlaswinkel, B. M. & H. R. Wanless, 2012. Rapid recycling of organic-rich carbonates during transgression: a complex coastal system in southwest Florida. Perspectives in Carbonate Geology: A Tribute to the Career of Robert Nathan Ginsburg. International Association of Sedimentologists Special Publication 41(98): 91–112.
Wanless, H. R., 1974. Mangrove sedimentation in geological perspective. In Gleason, P.J. (ed.), Environments of South Florida: Present and Past. Miami Geological Society Memoir 2: 190–200.
Wanless, H. R., R. Parkinson & L. Tedesco, 1994. Sea level control on stability of Everglades wetlands. In Davis, S. & J. Ogden (eds), The Florida Everglades: the Ecosystem and its Restoration. St Lucie Press, Delray Beach: 199–224.
Wdowinski, S., R. Bray, B. P. Kirtman & Z. Wu, 2016. Increasing flooding hazard in coastal communities due to rising sea level: case study of Miami Beach, Florida. Ocean & Coastal Management 126: 1–8.
Whelan, K. R., T. J. I. Smith III, D. R. Cahoon, J. C. Lynch & G. H. Anderson, 2005. Groundwater control of mangrove surface elevation: shrink and swell varies with soil depth. Estuaries 28: 833–843.
Whelan, K. R., T. J. Smith III, G. H. Anderson & M. L. Ouellette, 2009. Hurricane Wilma’s impact on overall soil elevation and zones within the soil profile in a mangrove forest. Wetlands 29: 16–23.
Woodroffe, C. D., 1981. Mangrove swamp stratigraphy and Holocene transgression, Grand Cayman Island, West Indies. Marine Geology 41: 271–294.
Woodroffe, C. D., 1995. Response of tide dominated mangrove shorelines in Northern Australia to anticipated sea level rise. Earth Surface Processes and Landforms 20: 65–85.
Acknowledgements
We would like to thank the SFWMD for funding the C-111 study (C-4244), L31E Pilot Project (C-12409), the Historic Creek Study (SFWMD 11679), and the Black Point Rehydration project (SFWMD x204114). Thanks to Gordon Anderson for providing elevation data for the Highway Creek and Joe Bay sites without recorders, Dr. Jerry Lorenz for providing elevation estimates, and Dr. Robert Fennema for providing contour maps of the western SESE. Special thanks to Dr. R.E. Turner for providing sediment accretion data. All Supporting Data are published as Electronic Supplementary Material. This is SERC Publication # 841.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: K. W. Krauss, I. C. Feller, D. A. Friess, R. R. Lewis III / Causes and Consequences of Mangrove Ecosystem Responses to an Ever-Changing Climate
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Meeder, J.F., Parkinson, R.W., Ruiz, P.L. et al. Saltwater encroachment and prediction of future ecosystem response to the Anthropocene Marine Transgression, Southeast Saline Everglades, Florida. Hydrobiologia 803, 29–48 (2017). https://doi.org/10.1007/s10750-017-3359-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-017-3359-0