
Abstract
Biomanipulation via fish regulation combined with submerged plant introduction is an effective measure to restore eutrophic shallow lakes. Improved water quality and clarity promote growth of benthic algae, which with submerged plants may limit sediment phosphorus (P) release, thereby reinforce lake recovery. Our study sought to evaluate the effect of such a biomanipulation on water quality, benthic algal development and sediment P release in a shallow, tropical lake by (1) comparing porewater and lake water quality, light intensity and benthic algal development in restored and unrestored sections; (2) conducting a 32P radiotracer experiment to track P release from sediment cores sampled from both sections. The biomanipulation led to lower total P, total dissolved P, and soluble reactive P concentrations in lake water, lower phytoplankton biomass, and increased light intensity at sediment surface, stimulating benthic algal development. Moreover, sediment 32P release was lower in the restored than unrestored section. Concurrently, dissolved oxygen levels in upper layers of the sediment cores were higher in the restored section. Our study indicates that the biomanipulation improved water quality and enhanced growth of benthic algae, thereby reducing sediment P release, which may be one of the main mechanisms to create successful restoration.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Eutrophication is one of the most common water quality problems in lakes worldwide (Schindler, 2006). As a major nutrient source in the aquatic environment, phosphorus (P) is a key contributing factor to eutrophication and, with it, deteriorating water quality and loss of biodiversity. P reduction is therefore an urgent priority in most lake restoration efforts (Schindler, 1977; Carpenter et al., 1995). However, despite intensive research into how to effectively reduce external loading, P remains a problem in water quality control (Jeppesen et al., 2007; Søndergaard et al., 2013). The most efficient way of improving the ecological status of shallow eutrophic lakes is to reduce the external nutrient loading (Jeppesen et al., 2007). However, when external loading has been reduced, increased release of P from the sediment has been observed, and this internal P loading may decide the trophic status of the lakes (van de Molen & Boers, 1994; Søndergaard et al., 2003, 2013). P sequestered in porewater is of particular concern due to its high mobility (Holdren & Armstrong, 1986), and even after a substantial reduction of the external loading, release of sediment P can contribute to continued high nutrient levels in the water column (Søndergaard et al., 2013), especially in lakes with high density of fish stirring up the sediment when foraging (Zhang et al., 2016). P release from the sediment, latest summarized by Søndergaard et al. (2013), is therefore an important issue to consider in lake restoration strategies.
Submerged plant growth often leads to reduced release of sediment P (Carignan & Kalff, 1980; Jaynes & Carpenter, 1986) through the direct nutrient uptake by the plants and by oxidation of sediment, which increases its P-binding capacity (Barko & James, 1998). Also, the submerged plants stabilize the sediment and reduce resuspension (Jensen et al., 2016). Submerged plants can also enhance the sedimentation of particles in the water column and decrease phytoplankton growth by competing for nutrients and light resources (Carignan & Kalff, 1980; Blindow et al., 1993). Moreover, a reduction of the biomass of coarse fish obtained by removal or stocking of piscivores can also diminish disturbance of the sediment (Drenner & Hambright, 1999; Jeppesen et al., 2012). Consequently, biomanipulation (fish regulation sometimes combined with submerged plant introduction) has been proposed as an effective ecological measure for improving water quality and clarity and generally promoting the recovery of eutrophic shallow lake ecosystems after an external nutrient loading reduction (Drenner & Hambright, 1999; Qiu et al., 2001; Jin et al., 2006; Jeppesen et al., 2012; Yu et al., 2016).
The increased availability of light at the sediment surface following the restoration allows colonization of benthic algae. Benthic algae have a high affinity for nutrients (Stevenson & Stoermer, 1982; McCormick & O’Dell, 1996; Pérez-Martínz et al., 2010) and may together with rooted macrophytes oxidize the top sediment layers and thus help immobilize nutrients in the sediment (Wetzel, 2001; Zhang et al., 2013), leading to increase in P retention in sediments (Carlton & Wetzel, 1988; Christensen et al., 1997) and lower release of P into the overlying water (Dodds, 2003; Tyler et al., 2003). In a previous study, sediment without benthic algae exhibited P release rates of about 1 mg P m−2day−1, whereas sediment with benthic algae released only 0.1 mg P m−2day−1 (van Luijn et al., 1995).
Although numerous studies have investigated the impact of submerged plants (Brenner et al., 1999; Rooney et al., 2003) and benthic algae on the P cycling between the sediment and the water column (Hansson, 1989; Dalsgaard, 2003; Spears et al., 2008; Zhang et al., 2013), little research has been undertaken to elucidate how the sediment P release is affected by biomanipulation in shallow lakes. This is unfortunate as changes in the P release may have profound effects on ecological state. In the present study, we hypothesized that lake biomanipulation (fish removal, stocking of piscivorous fish, plant introduction) would improve water quality and clarity, promoting growth of benthic algae. We further hypothesized that this increase in benthic algae may limit sediment phosphorus (P) release, thereby reinforcing lake recovery. We evaluated the effect of lake biomanipulation on water quality, benthic algae, and sediment P release by (1) comparing sediment porewater and lake water quality, light intensity as well as benthic algal development in restored and unrestored sections of a shallow lake and (2) conducting a 32P radiotracer experiment to track the P release from sediment cores sampled from restored and unrestored lake sections. We expected the P release to be substantially lower from the restored section.
Materials and methods
Lake description
Huizhou West Lake is a shallow eutrophic lake located in Huizhou City, Guangdong Province, South China. The lake is divided into several sub-lakes separated by bridges that do not block the movement of water. The lake covers 1.6 km2 and has an average depth of approximately 1.5 m. The average air temperature in the area is 22.0°C and the annual rainfall is 1889 mm. Nutrient levels in the sediment are high, for example in the lake section known as Fenghu TP and TN values are 1.31 and 2.39 mg g−1 as dry weight, respectively (Zhang et al., 2014).
Efforts to address the problem of eutrophication by the local government have been successful in cutting off point sources of external pollution, but the nutrient concentrations in lake water were still high with a Secchi depth of about 35 cm (Table 1) and submerged macrophytes were absent (Liu et al., submitted). In order to restore a section of the lake (as a demonstration), a total of 29.7 t (2.5 t ha−1) fish were removed in the Nanhu section by seine fishing, encompassing 65% tilapia (Oreochromis niloticus Linn.), 16% silver carp (Hypophthalmichthys molitrix Cuvier et Valenciennes) and bighead carp (Aristichthys nobilis Richardson), 12% common carp (Cyprinus carpio Linnaeus), and 1.6% long-tailed anchovy (Coilia grayii Richardson). Upon removal, transplantation of the submerged macrophytes Hydrilla verticillata Linnaeus f. Royle and Vallisneria natans Lour. Hara was conducted in 2007. V. natans ramets were planted in the sediment at a density of about 10–15 plants per m−2, and apical sections of young H. verticillata shoots were planted at a density of 20–30 plants m−2. Piscivorous fish (18,000 snakehead Channa argus Cantor and 16,000 mandarin fish Siniperca chuatsi Basilewsky, 1,500 and 1,333 ha−1, respectively) were also stocked (Liu et al., submitted).
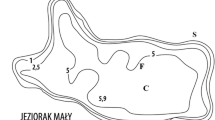
About 85% of the Nanhu section was covered by submerged plants in 2010, with concomitant improvements in water quality. The restored section was clear, incident light reaching all the way to the lake bottom (Fig. 1). For the purpose of the current study, sampling sites were established in the unrestored, unvegetated Fenghu section and in the restored, macrophyte-rich Nanhu section. Before restoration, water quality in the two sections was similar (Table 1).

West Lake in Huizhou City and the sampling sites in the two sections of the lake
Sampling and analysis
Sediment cores and P release experiment
Three 10-cm-deep sediment cores were taken along with overlying water from the selected sites in the unrestored Fenghu section and another three from open plant-free areas of the lake bed near the selected sites in the restored Nanhu section on March 12th, 2010. All cores were collected in perspex core tubes (15 cm length × 6.5 cm internal diameter). The tubes were immediately sealed at the top and bottom with silicon rubber stoppers to preserve the sediment structure. At the same time, 5 l lake water samples were collected in each section for use as experimental water. Both sediment cores and water samples were transported to the laboratory at Jinan University in boxes of ice.
The stoppers on the top of the sediment tubes were removed on arrival at the laboratory to allow gas exchange with air. At the start of the experiments, water overlying each core was siphoned off. The thickness of each sediment core was standardized to 5 cm from the bottom of each tube so that the sediment surface was 2.0 cm above the 5-mm-diameter 32P injection hole on the side of the tubes (Zhang et al., 2013). The holes were sealed with a silicon rubber stopper through which 32P could be injected into the sediment. Before initiating the experiment, 200 ml lake water was carefully added to each sediment tube.
10 μCi NaH 322 PO4 was injected into each sediment core using a syringe and a needle piercing the rubber stopper. A 0.5 ml water sample was collected from the middle of the water column of each tube 3, 6, 24, 48, and 72 h after NaH 322 PO4 addition. Water overlying the sediment was not stirred before sampling. The samples were transferred to a scintillation vial containing 10 ml scintillation cocktail (5.0 g 2,5-Diphenyloxazole + 0.5 g 1,4-bis (5-phenyloxazol-2-yl) benzene + 1,000 ml dimethylbenzene + 400 g tritonX-100). The 32P radioactivity (as counts per minute, dpm) of samples was assayed by liquid scintillation (Beckman Model LS6500, Beckman Coulter, Inc., Fullerton, CA) on March 15th and 18th, 2010, and corrected for the radioactive decay rate (Zhang et al., 2013).
The oxidation status of the sediment from both the restored and the unrestored sections was examined by measuring the dissolved oxygen (DO) concentration in the upper sediment layer using Unisense micro sensors (PA2000, Denmark).
Porewater and lake water
Samples from the sediment surface layer (0–5 cm depth) at three sites in the Fenghu and Nanhu sections were collected on March 12th, 2010, by a sediment sampler (Fig. 1) after which they put into sealed plastic bags and transported to the laboratory in iceboxes. Porewater was extracted from the sediments by centrifugation at 5,000 rpm for 15 min (Kalnejais et al., 2015) for nutrient analysis. Samples for total dissolved phosphorus (TDP) were filtered through GF/C filters (0.45 µm) and determined after persulphate digestion (APHA, 1992), by molybdenum-antimony-ascorbic acid colorimetry with Shimadzu UV 2550 spectrophotometry with 100 mm cuvette at 880 nm. Samples for soluble reactive phosphorus (SRP) determination were also filtered through GF/C filters (0.45 µm) without persulfate digestion.
One liter samples of lake water were collected 0.5 m beneath the surface at each sampling site and analyzed for a range of water quality parameters. All samples were kept cold and transported to the laboratory at the Jinan University in Guangzhou within 3 h of collection. The samples were analyzed for TP, TDP, and SRP, and phytoplankton biomass was measured as chlorophyll a (Chl a), being determined spectrophotometrically after ethanol extraction at room temperature according to Jespersen & Christoffersen (1987). Total phosphorus (TP) concentrations were determined after persulphate digestion (APHA, 1992). TDP and SRP were analyzed as described above for porewater.
Light and benthic algae
Light intensity at the bottom of the restored Nanhu and unrestored Fenghu sections was measured at all the sites using an underwater irradiance meter (ZDS-10W) between 9:00 to 12:00 am when collecting the sediment cores. Secchi depth (SD) was also measured using a Secchi disk.
Samples of sediment with benthic algae were taken from open areas of the Nanhu lake bed that were not directly colonized by submerged plants and situated close to the sediment sampling sites when collecting the sediment cores (Fig. 1). The algae were scraped from the sediment surface using a razor blade (Barbour et al., 1999), and their Chl a content was measured by spectrophotometry after ethanol extraction at room temperature according to Jespersen & Christoffersen (1987) to obtain a proxy for algal biomass. The dominant algal taxa were identified microscopically (Hu & Wei, 2006).
Statistical analyses
One-way ANOVA was performed to detect differences in TP, TDP, SRP, and Chl a concentrations and light intensity between samples from restored and unrestored sections. Repeated measurement ANOVAs were used to test for significant differences between restored and unrestored sections in 32P activity in the water column and DO concentrations in the sediment.
Results
P in porewater
Concentrations of porewater TDP and SRP were significantly lower (P < 0.05) in the restored section (TDP was 0.9 ± 0.3 μg l−1 and SRP was 0.3 ± 0.2 μg l−1) than in the unrestored site (TDP 20.2 ± 0.7 μg l−1 and SRP 14.6 ± 2.4 μg l−1). These restored section values for TDP and SRP represent about 4%, and 2% of those in the unrestored section, respectively.
P and Chl a in lake water
TP concentrations in March were significantly lower (F = 11.027, df = 1, P = 0.029, Fig. 2) in the water column from the restored than from the unrestored section. However, there was no significant between-section difference in water column TDP or SRP (P > 0.05).

P and Chl a concentrations in lake water (mean ± SD) from restored and unrestored sections. Asterisk indicates significant (P < 0.05) differences between the two sections
The water column Chl a concentration was 59.9 ± 19.2 μg l−1 in the unrestored section compared with 6.1 ± 2.0 μg l−1 in the restored section (Fig. 2), indicating a significant reduction (F = 23.198, df = 1, P = 0.009) in phytoplankton biomass through biomanipulation.
Light and benthic algae on the sediment surface
A highly significant difference in light intensity at the sediment surface was observed between the sections (F = 61.714, df = 1, P = 0.001). In the unrestored location, the sediment surface was completely dark and lake water SD was 37 ± 3 cm, while in the restored section light reached the sediment surface at an intensity of 21.6 ± 4.8 μmol m−2 s−1 at the time of sampling.
Benthic algae growing on the sediment surface in the restored Nanhu section were dominated by three taxa: Oscillatoria tenuis Ag. (Cyanophycea), Rhizoclonium hieroglyphicum Ag. Kütz (Chlorophyceae), and Navicula spp. (Bacillariophyta). Mean benthic algal biomass measured as Chl a was 78.7 ± 52.2 mg m−2. Owing to the complete absence of light, no benthic algae were present on the sediment surface in the unrestored Fenghu section.
Sediment P release
32P activity in the water column was lower in samples overlying sediment cores from the restored section than in sediment from the unrestored section (F = 13.474, df = 1, P = 0.021, Fig. 3). Furthermore, 32P began to appear in the water above the unrestored sediment within 6 h of injection but was almost completely absent in the restored sediment water column until the 24th hour, and low until at least the 48th hour after injection, indicating delayed release of P from the sediments carrying a layer of benthic algae.

Appearance of 32P activity (mean ± SD) in the water overlying the sediment cores during a 72-h period. At time zero, 10 μCi NaH 322 PO4 was injected into the sediment in each core through an injection hole of 20 mm below the surface
DO concentrations in sediment
Average DO concentrations in the uppermost sediment layers (2.0 mm depth) were significantly higher in the restored section (5.03 ± 0.19 mg l−1) than in the unrestored section (1.09 ± 0.94 mg l−1; F = 58.740, df = 1, P < 0.001, Fig. 4). The maximum depth of the oxic layer in the unrestored sediment was less than 2.5 mm compared with more than 3.5 mm in the restored sediment. Thus, the upper layers of the restored section sediments contained considerably more oxygen than those from the unrestored section.

Microprofile of DO concentration (mean + SD) in sediments from unrestored and restored sections of the lake bed
Discussion
We found that the biomanipulation undertaken in parts of shallow eutrophic Huizhou West Lake led to lower TP and Chl a concentrations in the water column and higher water clarity, allowing flourishing of benthic algae on the sediment surface of the lake bed. We also found lower TDP and SRP concentrations in porewater and lower release of injected 32P, whereas DO concentrations in the surface sediment were higher in the restored than in the unrestored section.
It has long been known that submerged macrophyte can reduce the concentration of P in sediment porewater by direct absorption of nutrients (Carignan & Kalff, 1980; Shilla et al., 2006) and indirectly by release of oxygen from their roots, promoting formation of oxidized iron crusts in anaerobic sediment layers and enhancing the sorption of P (Hupfer & Dollan, 2003). More P is therefore often retained in the sediment as Fe-bound (Fe–P), thus reducing the P release (Wang et al., 2007). In the present study, P concentrations were substantially lower in the porewater in the restored section sediments −96% for TDP and 98% for SRP compared with the levels in the unrestored section. This reduction is important because porewater TDP and SRP is highly mobile and easily transferred to the overlying lake water (Holdren & Armstrong, 1986). In the restored section, sediment was collected in open plant-free areas, implying that plants can only indirectly (reduced sedimentation and, as discussed below, improved light conditions) have affected the oxygen and P concentrations in the porewater.
An enhanced benthic algal growth in the restored section may have contributed to the reduced sediment P release, as seen in other studies (Carlton & Wetzel, 1988; Hansson, 1990; Wetzel, 2001). In shallow lakes, even a small increase in light intensity at the sediment surface can promote the growth of benthic algae, some of which can tolerate remarkably low light levels (Steinman & McIntire, 1987). A light compensation point of 4.7 μmol m−2 s−1 has been recorded for benthic algal communities (Wiencke, 2011); however, the light compensation point may vary for different species of algae within the benthic communities. For example, for Mougeotia, the light compensation point falls below 10 μmol m−2 s−1 (Graham et al., 1996), whereas Cladophora requires 29 μmol m−2 s−1 (Lorenz et al., 1991). In our study, we measured 21.6 ± 4.8 μmol m−2 s−1 at the time of sampling in the restored section, while no light reached the bottom in the unrestored section. The light reaching the sediment surface in the restored section was sufficient to induce benthic algal growth, as evidenced by benthic algae occurrence on the sediment surface in the restored but not in the unrestored section. Benthic algal growth may have contributed to the lower P release in the restored section by enhancing the oxygen concentration in the sediment and by releasing extracellular material that acts as a glue to stabilize the sediment surface, the latter enhancing cohesion and adhesion of sediment particles and thus limiting sediment resuspension (Tolhurst et al., 2008; Lubarsky et al., 2010) and sediment P release. A previous 32P lab study (Zhang et al., 2013) conducted at ambient room light intensity (daytime average: 15.8 ± 9.4 μmol m−2 s−1, darkness at night), with a dark/light cycle of about 12:12 h, showed greater P retention capacity in sediment with benthic algae than in controls: 93% of 32P being retained in the sediment with benthic algae compared with 84% in sediment without benthic algae. Our 32P tracing experiment, conducted under similar light conditions as the previous 32P lab study, concur with these results. Lower release means less phytoplankton and higher water clarity, which, in turn, stimulates benthic primary production further (Liboriussen & Jeppesen, 2003; Vadeboncoeur et al., 2003), and reduce P release from the sediment (Hansson, 1989).
Besides macrophytes and benthic algae, reduced disturbance by benthivorous fish and reduced sedimentation due to increased zooplankton grazing on phytoplankton (which means less oxygen consumption in the sediment and thus less P-release) may potentially contribute to the lower concentration of the various P fractions found in the restored section, as judged from temperate lakes (Jeppesen et al., 2012). These factors may, however, be of less importance in subtropical and tropical lakes. In the restored section in our study, the fish community was still quantitatively dominated by omni-benthivorous fish (tilapia and crucian carp Carassius auratus Linnaeus). Moreover, no notable difference was traced in total fish biomass or fish size among the sections (Gao et al., 2014). Also, there was no clear change in the zooplankton community: the abundance of cladocerans remained low and small species such as Chydorus sp., Alona spp., and Moina micrura Kurz continued to dominate, with low potential grazing on phytoplankton in both sections as judged from a low zooplankton:phytoplankton biomass ratio, likely due to high abundance of juvenile fish (Liu et al., submitted), indicating that the role of these alternative factors in changing the P concentrations and the sediment P release were of less importance.
Although TP concentrations in overlying water were lower in the restored than unrestored section, no significant differences of TDP and SRP concentrations were recorded between the two sections, indicating that the main change in TP was in particulate phosphorus (PP). The lower release of injected 32P from sediment cores in the restored section than unrestored section indicates that the restoration reduces P release from the sediment either by uptake by the more abundant benthic algae community or by complex binding to, for example, iron in the more oxidized sediment. It is important to notice, however, that accumulation of 32P in overlying water does not necessarily represent a net release of P from the sediment as isotopic exchange (with 31P) occurs after 32P addition in the sediment (Jensen et al., 2009).
In conclusion, biomanipulation including adjustment of the fish assemblage and introduction of submerged macrophytes can decrease TDP concentrations in porewater and P release from the sediment, thereby reducing the P concentration in the lake water. These effects may contribute to creating an improved water quality after restoration. Oxidation by benthic algae and macrophytes, and perhaps lower sedimentation of algae were likely the most influential factors in the restored section of Huizhou West Lake, but more detailed studies are needed to draw firm conclusions about the mechanisms involved and the contribution of the different factors.
References
APHA, 1992. Standard methods: for examination of water and wastewater, 18th ed. American Public Health Association.
Barbour, M. T., J. Gerritsen, B. D. Snyder & J. B. Stribling, 1999. Rapid bioassessment protocols for use in streams and wadeable rivers USEPA, Washington.
Barko, J. W. & W. F. James, 1998. Effects of submerged aquatic macrophytes on nutrient dynamics, sedimentation, and resuspension. In Jeppesen, E., M. Søndergaard, M. Søndergaard & K. Christoffersen (eds), The Structuring Role of Submerged Macrophytes in Lakes. Springer, New York: 197–214.
Blindow, I., G. Andersson, A. Hargeby & S. Johansson, 1993. Long-term pattern of alternative stable states in two shallow eutrophic lakes. Freshwater Biology 30: 159–167.
Brenner, M., T. J. Whitmore, M. A. Lasi, J. E. Cable & P. H. Cable, 1999. A multi-proxy trophic state reconstruction for shallow Orange Lake, Florida, USA: possible influence of macrophytes on limnetic nutrient concentrations. Journal of Paleolimnology 21: 215–233.
Carignan, R. & J. Kalff, 1980. Phosphorus sources for aquatic weeds: water or sediments? Science 207: 987–989.
Carlton, R. G. & R. G. Wetzel, 1988. Phosphorus flux from lake sediments: effects of epipelic algal oxygen production. Limnology and Oceanography 33: 562–570.
Carpenter, S. R., D. L. Christensen, J. J. Cole, K. L. Cottingham, X. He, J. R. Hodgson, J. F. Kitchell, S. E. Knight, M. L. Pace, D. M. Post, D. E. Schindler & N. Voichick, 1995. Biological control of eutrophication in lakes. Environmental Science & Technology 29: 784–786.
Christensen, K. K., F. O. Andersen & H. S. Jensen, 1997. Comparison of iron, manganese and phosphorus retention in freshwater litorral sediment with growth of Litorella uniflora and benthic microalgae. Biogeochemistry 38: 149–171.
Dalsgaard, T., 2003. Benthic primary production and nutrient cycling in sediments with benthic microalgae and transient accumulation of macroalgae. Limnology and Oceanography 48: 2138–2150.
Dodds, W. K., 2003. The role of periphyton in phosphorus retention in shallow freshwater aquatic systems. Journal of Phycology 39: 840–849.
Drenner, R. W. & K. D. Hambright, 1999. Biomanipulation of fish assemblages as a lake restoration technique. Archiv für Hydrobiologie 146: 129–165.
Gao, J., Z. Liu & E. Jeppesen, 2014. Fish community assemblages changed but biomass remained similar after lake restoration by biomanipulation in a Chinese tropical eutrophic lake. Hydrobiologia 724: 127–140.
Graham, J. M., P. Arancibia-Avila & L. E. Graham, 1996. Physiological ecology of a species of the filamentous green alga Mougeotia under acidic conditions: light and temperature effects on photosynthesis and respiration. Limnology and Oceanography 41: 253–261.
Hansson, L. A., 1989. The influence of a periphytic biolayer on phosphorus exchange between substrate and water. Archiv für Hydrobiologie 115: 21–26.
Hansson, L. A., 1990. Quantifying the impact of periphytic algae on nutrient availability for phytoplankton. Freshwater Biology 24: 265–273.
Holdren, G. C. & D. E. Armstrong, 1986. Interstitial ion concentrations as an indicator of phosphorus release and mineral formation in lake sediments. In Sediments and Water Interactions. Springer, New York: 133–147
Hu, H. J. & Y. X. Wei, 2006. The Freshwater Algae of China: Systematics, Taxonomy and Ecology. Science Press, Beijing.
Hupfer, M. & A. Dollan, 2003. Immobilisation of phosphorus by iron-coated roots of submerged macrophytes. Hydrobiologia 506: 635–640.
Jaynes, M. L. & S. R. Carpenter, 1986. Effects of vascular and nonvascular macrophytes on sediment redox and solute dynamics. Ecology 67: 875–882.
Jensen, H. S., O. L. Nielsen, M. S. Koch & I. de Vicente, 2009. Phosphorus release with carbonate dissolution coupled to sulfide oxidation in Florida Bay seagrass sediments. Limnology and Oceanography 54: 1753–1764.
Jensen, M., Z. W. Liu, X. F. Zhang, K. Reitzel & H. S. Jensen. 2016. The effect of biomanipulation on phosphorus exchange between sediment and water in shallow, tropical Huizhou West Lake, China. Limnologica (Accepted)
Jeppesen, E., M. Søndergaard, M. Meerhoff, T. L. Lauridsen & J. P. Jensen, 2007. Shallow lake restoration by nutrient loading reduction-some recent findings and challenges ahead. Hydrobiologia 584: 239–252.
Jeppesen, E., M. Søndergaard, T. L. Lauridsen, T. A. Davidson, Z. W. Liu, N. Mazzeo, C. Trochine, K. Özkan, H. S. Jensen, D. Trolle, F. Starling, X. Lazzaro, L. S. Johansson, R. Bjerring, L. Liboriussen, S. E. Larsen, F. Landkildehus, S. Egemose & M. Meerhoff, 2012. Biomanipulation as a restoration tool to combat eutrophication: recent advances and future challenges. Advances in Ecological Research 47: 411–488.
Jespersen, A. M. & K. Christoffersen, 1987. Measurements of chlorophyll a from phytoplankton using ethanol as extraction solvent. Archiv für Hydrobiologie 109: 445–454.
Jin, X. C., Q. J. Xu & C. Z. Yan, 2006. Restoration scheme for macrophytes in a hypertrophic water body, Wuli Lake, China. Lakes & Reservoirs: Research and Management 11: 21–27.
Kalnejais, L. H., W. R. Martin & M. H. Bothner, 2015. Porewater dynamics of silver, lead and copper in coastal sediments and implications for benthic metal fluxes. Science of the Total Environment 517: 178–194.
Liboriussen, L. & E. Jeppesen, 2003. Temporal dynamics in epipelic, pelagic and epiphytic algal production in a clear and a turbid shallow lake. Freshwater Biology 48: 418–431.
Liu, Z., P. Zhong, X. Zhang, J. Ning, S. E. Larsen & E. Jeppesen. Successful restoration of a tropical shallow eutrophic lake by biomanipulation: strong bottom-up but weak top-down effects recorded (Submitted)
Lorenz, R. C., M. E. Monaco & C. E. Herdendorf, 1991. Minimum light requirements for substrate colonization by Cladophora glomerata. Journal of Great Lakes Research 17: 536–542.
Lubarsky, H. V., C. Hubas, M. Chocholek, F. Larson, W. Manz, D. M. Paterson & S. U. Gerbersdorf, 2010. The stabilisation potential of individual and mixed assemblages of natural bacteria and microalgae. PLoS ONE 5: e13794.
McCormick, P. V. & M. B. O’Dell, 1996. Quantifying periphyton responses to phosphorus in the Florida Everglades: a synoptic-experimental approach. Journal of the North American Benthological Society 15: 450–468.
Pérez-Martínez, C., P. Sánchez-Castillo & M. Z. Jiménez-Pérez, 2010. Utilization of immobilized benthic algal species for N and P removal. Journal of Applied Phycology 22: 277–282.
Qiu, D. R., Z. B. Wu, B. Y. Liu, J. Q. Deng, G. P. Fu & F. He, 2001. The restoration of aquatic macrophytes for improving water quality in a hypertrophic shallow lake in Hubei Province, China. Ecological Engineering 18: 147–156.
Rooney, N., J. Kalff & C. Habel, 2003. The role of submerged macrophyte beds in phosphorus and sediment accumulation in Lake Memphremagog, Quebec, Canada. Limnology and Oceanography 48: 1927–1937.
Schindler, D. W., 1977. Evolution of phosphorus limitation in lakes. Science 195(4275): 260–262.
Schindler, D. W., 2006. Recent advances in the understanding and management of eutrophication. Limnology and Oceanography 51: 356–363.
Shilla, D. A., T. Asaeda, S. Kian, R. Lalith & J. Manatunge, 2006. Phosphorus concentration in sediment, water and tissues of three submerged macrophytes of Myall Lake, Australia. Wetlands Ecology and Management 14: 549–558.
Søndergaard, M., J. P. Jensen & E. Jeppesen, 2003. Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 506: 135–145.
Søndergaard, M., R. Bjerring & E. Jeppesen, 2013. Persistent internal phosphorus loading during summer in shallow eutrophic lakes. Hydrobiologia 710: 95–107.
Spears, B. M., L. Carvalho, R. Perkins & D. M. Paterson, 2008. Effects of light on sediment nutrient flux and water column nutrient stoichiometry in a shallow lake. Water Research 42: 977–986.
Steinman, A. D. & C. D. McIntire, 1987. Effects of irradiance on the community structure and biomass of algal assemblages in laboratory streams. Canadian journal of fisheries and aquatic sciences 44: 1640–1648.
Stevenson, R. J. & E. F. Stoermer, 1982. Luxury consumption of phosphorus by benthic algae. Biological Science 32: 682–683.
Tolhurst, T. J., M. Consalvey & D. M. Paterson, 2008. Changes in cohesive sediment properties associated with the growth of a diatom biofilm. Hydrobiologia 596: 225–239.
Tyler, A. C., K. J. McGlathery & I. C. Anderson, 2003. Benthic algae control sediment – water column fluxes of organic and inorganic nitrogen compounds in a temperate lagoon. Limnology and Oceanography 48: 2125–2137.
Vadeboncoeur, Y., E. Jeppesen, M. Zanden, H. H. Schierup, K. Christoffersen & D. M. Lodge, 2003. From Greenland to green lakes: cultural eutrophication and the loss of benthic pathways in lakes. Limnology and Oceanography 48: 1408–1418.
van der Molen, D. T. & P. C. M. Boers, 1994. Influence of internal loading on phosphorus concentration in shallow lakes before and after reduction of the external loading. Hydrobiologia 275(276): 379–389.
van Luijn, F., D. T. van der Molen, W. J. Luttmer & P. C. M. Boers, 1995. Influence of benthic diatoms on the nutrient release from sediments of shallow lakes recovering from eutrophication. Water Science and Technology 32: 89–97.
Wang, S. R., X. C. Jin, H. C. Zhang, X. N. Zhou & F. C. Wu, 2007. Effects of Hydrilla verticillata on Phosphorus Retention and Release in Sediments. Water Air Soil Pollution 181: 329–339.
Wetzel, R. G., 2001. Limnology: Lake and River Ecosystems. Academic Press, San Diego.
Wiencke, C., 2011. Biology of Polar Benthic Algae. Walter de Gruyter, Berlin.
Zhang, X., Z. Liu, R. D. Gulati & E. Jeppesen, 2013. The effect of benthic algae on phosphorus exchange between sediment and overlying water in shallow lakes: a microcosm study using 32P as a tracer. Hydrobiologia 710: 109–116.
Zhang, X., Z. Liu, E. Jeppesen & W. D. Taylor, 2014. Effects of deposit-feeding tubificid worms and filter-feeding bivalves on benthic-pelagic coupling: implications for the restoration of eutrophic shallow lakes. Water Research 50: 135–146.
Zhang, X., Z. Liu, E. Jeppesen, W. D. Taylor & L. G. Rudstam, 2016. Effects of benthic-feeding common carp and filter-feeding silver carp on benthic-pelagic coupling: implications for shallow lake management. Ecological Engineering 88: 256–264.
Yu, J., Z. Liu, K. Li, F. Chen, B. Guan, Y. Hu, P. Zhong, Y. Tang, X. Zhao, H. He, H. Zeng & E. Jeppesen, 2016. Restoration of shallow lakes in subtropical and tropical China: response of nutrients and water clarity to biomanipulation by fish removal and submerged plant transplantation. Water. doi:10.3390/w8100438.
Acknowledgements
We thank Anne Mette Poulsen for valuable editorial work and Henning Skovgaard Jensen for constructive comments of this manuscript. The study was sponsored by the National Natural Science Foundation of China (No. 31570456 and 31000219) and the Natural Science Foundation of Guangdong Province (No. 2016A030313103) and was completed while the first author was a visiting associate professor at Cornell University. EJ was supported by the MARS project (Managing Aquatic ecosystems and water Resources under multiple Stress) funded under the 7th EU Framework Programme, Theme 6 (Environment including Climate Change), Contract No.: 603378 (http://www.mars-project.eu), and the Danish project CLEAR (a Villum Kann Rasmussen Centre of Excellence project).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Karl E. Havens
Rights and permissions
About this article

Cite this article
Zhang, X., Tang, Y., Jeppesen, E. et al. Biomanipulation-induced reduction of sediment phosphorus release in a tropical shallow lake. Hydrobiologia 794, 49–57 (2017). https://doi.org/10.1007/s10750-016-3079-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-016-3079-x