
Abstract
More than half of the natural wetlands in the Illinois River valley (IRV) have been lost through conversion of floodplain wetlands and lakes to drainage and levee districts for agricultural production. During 2007–2013, we monitored the response of wetland vegetation communities to restoration at Emiquon Preserve, a former floodplain that was drained and farmed for more than 80 years. Spatial coverage of wetland vegetation and other cover types rapidly expanded from 252 ha in 2007 to 1,944 ha in 2013 (mean 1,512 ± 239 ha) with little supplemental planting or hydrological management. Restored vegetation emulated aquatic plant communities that were largely eliminated from the IRV, most importantly floating-leaved and submersed aquatic vegetation. Mean annual wetland cover included aquatic bed (44%), open water (20%), persistent emergent vegetation (10%), hemi-marsh (10%), and nonpersistent emergent vegetation (9%). Average moist-soil seed and tuber density was similar to managed wetlands in the IRV (mean 724 ± 224 kg/ha). A viable option for restoration of diverse aquatic macrophyte communities within degraded wetlands of large river systems includes passive restoration of hydrology and vegetation behind levees while maintaining infrastructure to facilitate drawdowns when necessary or to mimic historical conditions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Wetlands within and nearby floodplains of large rivers historically provided extensive and valuable habitats to migrating waterbirds and other wetland-dependent wildlife in the Upper Midwest (Havera, 1999). Drainage of wetlands for agriculture was particularly pervasive in the Midwestern states of Illinois, Indiana, Iowa, and Ohio, where losses have exceeded 85% (Dahl, 1990). According to Pavelis (1987), nearly 45 million ha of rural land in the United States had been drained by 1985. Moreover, Illinois led the nation with nearly 4 million ha of rural land area drained, while 50% of this area was served by drainage organizations (e.g., drainage and levee districts). Many of the natural wetlands in the Illinois River valley (IRV) have been lost due to large-scale anthropogenic changes, while sedimentation, fluctuating water levels, exotic and invasive species, and excessive nutrient loads have diminished the quality of current wetland conditions (Havera, 1999). For instance, drainage and levee districts were formed throughout the IRV during the early 1900s separating the river from floodplains and adjacent wetlands. Floodplain wetlands and backwater lakes were separated from the river by levees and drained by pumps through a network of ditches to convert the fertile floodplain soil to agriculture. By the early 1920s, approximately half of the Illinois River floodplain between La Salle and Grafton, IL was within drainage and levee districts (Mills et al., 1966; Bellrose et al., 1979, 1983; Havera et al., 1995).
The remaining lakes and wetlands of the IRV have received decades of excessive sediment deposition from agricultural runoff (Bellrose et al., 1983), and they are subjected to raised and extremely variable hydrology preventing growth of aquatic macrophytes (Sparks et al., 1998). Furthermore, the Illinois River contains persistent, invasive species which can degrade restored wetlands (Raibley et al., 1995; Bajer et al., 2009; Irons et al., 2009). Wetlands and lakes with open hydrologic connections to the Illinois River typically have low water transparency during the growing season and are devoid of aquatic macrophytes (Mills et al., 1966; Sparks et al., 1998; Yin & Langrehr, 2005; Moore et al., 2010). Aquatic macrophytes stabilize sediments and increase water clarity (Madsen et al., 2001), provide food and cover for fish and wildlife (Johnson & Jennings, 1998; Stafford et al. 2010), and provide nutrient cycling (Mitsch et al., 2012). While river-floodplain connectivity in the IRV may benefit nutrient cycling, flood storage, and provide access to spawning and overwinter habitat for some riverine fishes (Sparks, 1995), the long-term consequences of accretion from sedimentation and the loss of aquatic macrophytes illustrate the tradeoffs of river-floodplain connectivity in a highly modified river system (Jackson & Pringle, 2010).
Numerous projects have aimed to restore or enhance floodplain wetland quality throughout the IRV, but success has been variable and occasionally hampered by frequent flooding from the Illinois River (Lemke et al., 2016a). Bellrose et al. (1983) and Sparks et al. (1998) noted that conservation organizations could acquire existing drainage and levee districts, and conduct studies to advance our understanding of the structure and function of floodplain ecosystems. Restoring wetlands within drainage and levee districts has several advantages: (1) drained floodplains were once wetlands and hydric conditions will likely return quickly following return of hydrology; (2) existing agricultural drainage infrastructure will allow isolation from major floods and a buffer against sediment inputs; and (3) experiments can be conducted to better describe the effects of river-floodplain connectivity on aquatic vegetation communities. Following this impetus, The Nature Conservancy (TNC) acquired the Thompson Lake Drainage and Levee District along with some adjacent properties in 2000 and established the Emiquon Preserve (hereafter Emiquon) to restore an important floodplain along the Illinois River and advance restoration ecology (Walk et al., 2016). Wetland restoration began at Emiquon in 2007 when TNC turned off drainage pumps and allowed groundwater and precipitation to naturally fill the historical lake beds and adjacent wetlands, which remained separated from the Illinois River by the original drainage district levees. A unique feature of the restoration of Emiquon Preserve was maintenance of isolation from the Illinois River, and its position within a historical floodplain where several backwater lakes had been an important habitat for fish and wildlife prior to drainage in the 1920s (Mills et al., 1966).
Key ecological attributes (KEAs) were established by TNC to evaluate restoration success with a variety of floral and faunal metrics that could indicate when management actions were needed to produce target ecological conditions (Parrish et al., 2003). Among these, KEAs were elements of vegetation structure indicative of habitat conditions for wildlife and wetland quality (Lemke et al., 2016b). We monitored the response of wetland vegetation communities and other cover types to restoration at Emiquon during 2007–2013 to evaluate a set of KEAs relative to desired ecological conditions established by TNC and those important to wetland conservation planners (Soulliere et al., 2007; Lemke et al., 2016b). Our objectives were to (1) document the composition and arrangement of wetland vegetation communities and other cover types through geospatial covermapping, (2) estimate moist-soil plant seed and tuber density to index potential forage habitat quality for waterfowl, and (3) describe changes in cover types and moist-soil forage quality following restoration. We hypothesized that wetland vegetation communities would emulate characteristics of historical wetlands and floodplain lakes in the IRV due to isolation from the Illinois River. Furthermore, we hypothesized that former agricultural fields would be colonized by moist-soil vegetation early in the restoration and seed, and tuber densities would be similar to those of other contemporary moist-soil wetlands in the IRV.
Methods
Study area
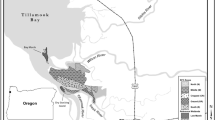
We monitored restoration of Emiquon Preserve, a 2,400-ha former floodplain of the Illinois River which contained the historical Thompson, Flag, and Siebs lake basins and was farmed for more than 80 years prior to restoration (Havera et al., 2003). Thompson Lake was the largest floodplain lake in the IRV and was the most productive backwater with respect to fish and wildlife populations prior to drainage in the early 1920s (Havera et al., 2003). Emiquon Preserve is located between Illinois River miles 121–126 in Fulton County, Illinois, USA (Fig. 1) and was designated a Ramsar Wetland of International Importance in 2012. Maximum water depth within Emiquon Preserve during our study was <6 m.

Location of the Emiquon Preserve (river miles 121–126) within La Grange Pool of the Illinois River, Fulton County, Illinois, USA
Wetland mapping
We delineated all wetland vegetation communities, mudflats, and areas containing surface water at Emiquon each autumn (late August to early November) during 2007–2013 to document annual changes in wetland area, vegetation communities, and cover type area. We traversed east–west transects spaced at 500-m intervals during early autumn each year and delineated changes in vegetation communities and other cover types (e.g., open water, mudflats), and recorded plant species encountered using a handheld global positioning system (GPS) combined with paper maps and data forms or field computer with Arc Pad software (Bowyer et al., 2005; Stafford et al., 2010). We used transects only as a guide for field personnel, as cover types between transect lines were also delineated. We digitized cover types using photointerpretation in ArcGIS 9.3, 10.0, and 10.1 using color aerial photos obtained from the U.S. Department of Agriculture’s Geospatial Data Gateway in 2007, high-resolution color aerial photographs from Sanborn Map Company (Chesterfield, MO) during 2008–2011, and color infrared aerial photographs from the U.S. Fish and Wildlife Service (Region 3 Office, Twin Cities, MN) and the U.S. Geological Survey (Upper Midwest Environmental Sciences Center, La Crosse, WI) in 2012 and 2013, respectively (Bowyer et al., 2005; Stafford et al., 2010). Aerial imagery was <1 m resolution.
Our nomenclature and classification rules generally followed Cowardin et al. (1979) and Suloway & Hubbell (1994). Woody vegetation was classified as bottomland forest if >6 m in height or scrub shrub if 1–6 m (Cowardin et al., 1979). Other wetland classifications included nonpersistent emergent vegetation [i.e., moist-soil plants (annual and perennial hydrophytic plants that colonize mudflats and produce seeds and tubers important to wetland wildlife as forage and cover; Fredrickson & Taylor, 1982)], persistent emergent vegetation (e.g., cattails [Typha L. spp.]), mudflats, floating-leaved aquatic vegetation (e.g., American lotus [Nelumbo lutea Willd.]), aquatic bed (i.e., submersed aquatic plants; e.g., coontail [Ceratophyllum demersum L.]), hemi-marsh (i.e., open water or aquatic bed interspersed with persistent emergent vegetation in a ratio ranging from 30 to 70% by ocular estimate; Weller & Spatcher, 1965), and open water (i.e., flooded habitat with no visible submersed or emergent vegetation). We also included a category to account for areas of herbaceous upland vegetation (e.g., goldenrod [Solidago L. spp.]) growing within the wetland basin that had been inundated.
We used a multivariate analysis of variance (MANOVA) to evaluate changes in vegetation communities and other cover types at Emiquon during our study period. Due to limited sample size, we selected the five most dominant cover types for analysis. We used the percent of wetland area comprising aquatic bed vegetation, open water, nonpersistent emergent vegetation, persistent emergent vegetation, and hemi-marsh vegetation as dependent variables and year as the independent variable (Crook et al., 2009). Similarly, we used simple linear regression to describe changes in frequency of occurrence of undesirable, invasive species in covermap polygons. We designated year as an independent variable and frequency of occurrence (i.e., percent of polygons where each taxa was encountered during covermapping) as the dependent variable. We did not include total wetland area because it was correlated with year (r = 0.78). We designated α = 0.10 since our sample size was small and our study was observational in nature and used plots of residuals to ensure a normalized distribution. We conducted all analyses in SAS v9.3 (SAS Institute, Inc., Cary, NC) and reported values as means ± standard error (SE).
We consolidated vegetation communities into eight categories: bottomland forest, nonpersistent emergent, open water, aquatic bed, floating-leaved aquatic, mudflat, persistent emergent (persistent emergent + hemi-marsh vegetation), and scrub shrub (Stafford et al., 2010; Table 3) to compare wetland characteristics at Emiquon to those of historical (1938–1942; Bellrose, 1941; Bellrose et al., 1979) and contemporary (2005–2006; Stafford et al., 2010) wetlands in the La Grange and Peoria reaches in the IRV. We used cover data from other contemporary wetlands in the IRV reported by Stafford et al. (2010) as our control, as wetland conditions during their study were similar to wetland conditions in the IRV during our study period. We omitted the area of upland vegetation that was flooded and ditched from this analysis, since areas comprised a small proportion of Emiquon Preserve during the majority of our study period and historical data were unavailable for these cover types. Finally, we computed an interspersion–juxtaposition index (IJI) after Stafford et al. (2010) as a measure of heterogeneity of wetland vegetative structure at Emiquon during 2007–2013 (Weller & Spatcher, 1965; McGarigal & Marks, 1995). We converted the wetland cover maps from polygons to rasters (10 m cells) in ERDAS IMAGINE 2015, and output IJI values using FRAGSTATS 4.2 (McGarigal et al., 2012). The IJI measures the extent to which wetland vegetation communities are interspersed, expressed as a percentage of the total number of cover types. Thus, higher IJI values correspond to increased interspersion of cover types (McGarigal & Marks, 1995).
Moist-soil seed density
During early autumn 2007–2013, we estimated above- and below-ground biomass of moist-soil plant seeds by extracting a soil core (10-cm diameter × 5-cm depth) in nonpersistent emergent vegetation. We collected soil cores from 16 to 20 random points each year near the terminus of vegetation transects along the western shore of Emiquon where moist-soil vegetation occurred consistently during our study period (Stafford et al., 2006, 2011). We froze soil cores in individually labeled bags until processing in a laboratory. We thawed core samples at room temperature and soaked them in a 3% solution of hydrogen peroxide (H2O2) to dissolve clays (Bohm, 1979; Kross et al., 2008), washed samples with water through a 250-μm sieve, and dried for 24 h at approximately 87°C (Greer et al., 2007; Stafford et al., 2011). We threshed dried materials over a series of 4–5 sieves (mesh sizes 1,400, 1,000, 500, 355, and 250 μm; Greer et al., 2007) and separated all seeds and tubers retained by the three largest sieves from debris by hand, while seeds retained by the two smallest sieves (e.g., nutgrasses [Cyperus L. spp.]) were subsampled (2.5–50% by mass) to increase processing efficiency (Stafford et al., 2011). All seeds and tubers were weighed to the nearest 0.1 mg. We adjusted estimates for seed recovery bias using size-specific correction factors and combined small and large seed masses to estimate total seed biomass (kg/ha, dry mass; Hagy et al., 2011). We used a one-way analysis of variance (Proc GLM in SAS v9.3) to test for an effect of year on total moist-soil seed density. We designated α = 0.10 since our sample size was small and our study was observational in nature and used plots of residuals to ensure a normalized distribution.
We used our overall estimates of seed density to estimate energetic carrying capacity for waterfowl, expressed as energetic use days (EUD). A EUD is defined as the number of days that a given area could support the energetic requirements of a duck in our study area (Reinecke et al., 1989; Stafford et al., 2011). EUDs are used as an indicator of wetland quality and are an important metric for use in conservation planning models (Soulliere et al., 2007; Gray et al., 2013). We used an average true metabolizable energy of 2.5 kcal/g for moist-soil plant seeds (Kaminski et al., 2003) and an average daily energy expenditure representative of dabbling ducks migrating through the IRV (337 kcal/day; Stafford et al., 2011) for EUD calculations (Gray et al., 2013). We presented seed abundance and EUD values as means ± SE.
Results
Wetland mapping
We identified more than 120 plant taxa and mapped 8 vegetation communities and other wetland cover types at Emiquon (Fig. 2). Spatial coverage of wetland or inundated vegetation ranged from 252 ha in 2007 to 1,944 ha in 2013 (mean 1,512 ± 239 ha; Table 1). The five dominant vegetation communities and cover types differed by year (F = 267.13, P = 0.046), with the proportion of aquatic bed (t = 2.14, P = 0.085) increasing and the proportions of hemi-marsh (t = −2.29, P = 0.070) and open water (t = −2.07, P = 0.093) decreasing during 2007–2013.

The change in wetland vegetation communities and other cover types at the Emiquon Preserve from fall 2007 (252 ha) to fall 2013 (1,944 ha). White space within the Emiquon boundary indicates nonwetland vegetation
Aquatic bed was the dominant wetland community at Emiquon, comprising an average of 44% (mean 784 ± 177 ha) of the wetland area since 2007 and 56% since 2009 (Table 1; Fig. 3). We found negative correlations between the proportion of aquatic bed and proportions of open water (r = −0.90, P = 0.01), nonpersistent emergent vegetation (r = −0.89, P = 0.01), and persistent emergent vegetation (r = −0.75, P = 0.08). Open water was the next largest cover type (mean 252 ± 27 ha, 20%; Tables 1, 3) at Emiquon, and proportionally remained relatively stable during 2009–2013 (12–18%; Fig. 3). The proportion of open water was positively correlated (r = 0.84, P = 0.03) with persistent emergent vegetation. The area of hemi-marsh (mean 141 ± 33 ha, 10%) increased more than eightfold from 2007–2009, but declined 53% during 2009–2013 (Table 1). During 2010–2013, hemi-marsh remained relatively constant in proportional area (4–7%). We detected a negative relationship (r = −0.78, P = 0.06) between proportions of hemi-marsh and persistent emergent vegetation. The area of persistent emergent vegetation steadily increased at Emiquon during our period of study, ranging from 33 ha in 2007 to 294 ha in 2013 and averaging 10% (mean 157.6 ± 44.3 ha) of the wetland area during the 7 years of study (Table 1; Fig. 3). The amount of nonpersistent emergent vegetation at Emiquon was variable, ranging from 24 ha in 2009 to 218 ha in 2010 (mean 108 ± 26 ha; 9%; Tables 1, 3; Fig. 3). The proportion of nonpersistent emergent vegetation was positively correlated with persistent emergent (r = 0.82, P = 0.04). Lastly, annual IJI values at Emiquon ranged from 59.5 to 66.3% and averaged 61.5 ± 1.2% (SE) during 2007–2013.

Changes in surface-water elevations (meters above mean sea level) and cover types at the Emiquon Preserve during 2007–2013
Emiquon comprised less bottomland forest (mean < 0.1%) and open water (mean 20.2 ± 4.0%) than historical (bottomland forest mean 8.8 ± 1.3%; open water mean 38.7 ± 3.9%) and other contemporary wetlands (bottomland forest mean 15.3 ± 2.2%; open water mean 37.6 ± 6.6%) in the IRV (Table 3). The proportions of nonpersistent emergent (mean 8.9 ± 2.4%) and scrub-shrub (mean 0.6 ± 0.4%) vegetation communities at Emiquon were similar to historical (nonpersistent emergent mean 12.4 ± 2.8%; scrub-shrub mean 1.3 ± 0.5%) wetlands, but less than those in contemporary (nonpersistent emergent mean 32.5 ± 4.8%; scrub-shrub mean 5.2 ± 0.8%) wetlands. Conversely, the proportions of aquatic bed (mean 43.6 ± 8.8%) and persistent emergent (mean 20.5 ± 1.2%) vegetation communities at Emiquon exceeded those in both historical (aquatic bed mean 11.2 ± 2.6%; persistent emergent mean 12.3 ± 2.0%) and contemporary (aquatic bed mean <0.1 ± 4.4%; persistent emergent mean 3.9 ± 3.4%) wetlands in the IRV. Finally, the proportions of floating-leaved aquatic vegetation (mean 0.2 ± 0.1%) and mudflats (mean 1.6 ± 0.8%) at Emiquon were similar to contemporary (floating-leaved mean <0.1 ± 3.7%; mudflats mean 1.7 ± 0.3%) wetlands. Historical wetlands contained more floating-leaved aquatic vegetation (mean 14.9 ± 2.2%) but fewer mudflats (mean 0.4 ± 0.2%) than Emiquon and contemporary wetlands (Table 3).
We detected a positive relationship between year since restoration and frequency of occurrence of several invasive species, including common reed (Phragmites Adans. spp. [t = 5.96, P = 0.002]), reed canary grass (Phalaris arundinacea L. [t = 2.44, P = 0.059]), Eurasian watermilfoil (Myriophyllum spicatum L. [t = 4.23, P = 0.008]), and total invasive species (t = 4.69, P = 0.005). The frequency of occurrence of total invasive species ranged from 2 to 46% during our period of study. Detections of curly pondweed (Potamogeton crispus L.), purple loosestrife (Lythrum salicaria L.), and other undesirable, invasive species were too infrequent to analyze, but there were no apparent trends over time.
Moist-soil plant seed density
We collected 136 soil core samples on the west shore of Emiquon during autumns 2007–2013. Moist-soil plant seed density ranged from 256 kg/ha in 2009 to 1,246 kg/ha in 2011 (mean 724 ± 224 kg/ha; Table 2). Similar to seed density estimates, EUDs also were variable, ranging from 1,899 EUDs/ha in 2009 to 9,244 EUDs/ha in 2011 (mean 5,373 ± 634 EUDs/ha; Table 2).
Discussion
Aquatic cover types
The wetland area mapped at Emiquon increased more than 600% from 2007 to 2009, as the site developed into a mix of open water and diverse vegetation types (Table 1). Correspondingly, the average water elevation during our wetland mapping periods increased by 2.8 m. Subsequently, mean water elevations exhibited only a modest increase (0.1 m) from 2009 to 2013, but represented an 8% increase in wetland area (Table 1; Fig. 3). Rapid growth of the wetland area was likely enhanced by abundant precipitation during 2008–2009 (Changnon & Black, 2009; Changnon et al., 2010). During the initial year of restoration (2007), the wetland area was limited to the historic Thompson Lake basin, creating an open water environment with moist-soil plants (i.e., nonpersistent emergent) and cattails (i.e., persistent emergent) developing around the wet perimeter. By autumn 2008, water levels rose in Thompson Lake promoting the growth of submersed aquatic vegetation and inundating stands of cattails, which expanded the hemi-marsh community. Despite a lack of planting or water input from external sources which might have carried propagules, the aquatic bed community grew substantially from just 1% of the total wetland area in 2007 to 66% in 2009 (Fig. 3). Viable seeds of aquatic macrophytes can persist in the seed bank for decades, ensuring regeneration of aquatic communities following extended dry periods (Wienhold & van der Valk, 1989; McFarland & Rogers, 1998; Brock et al., 2003). Furthermore, Casanova (2012) demonstrated that seed banks of some aquatic plants in farmed wetlands exhibited resiliency to such extreme disturbances such as drought and tillage. As submersed aquatic and persistent emergent vegetation have substantially declined in the IRV since the mid-1900s (Stafford et al., 2010), it is noteworthy that these vegetation communities developed at Emiquon without supplemental seeding or planting by managers.
In just three years, the Emiquon Preserve plant communities naturally transitioned from shallowly flooded herbaceous plants and crop stubble to a diverse mix of facultative and obligate wetland plants that are uncommon in the IRV. Tradeoffs were apparent as hydrologic conditions produced environments more favorable to obligate wetland plants. For instance, the decline in suitable conditions for moist-soil plant development due to increasing area of shallow (<2 m) water during the growing season promoted rapid expansion of aquatic bed plant communities. Furthermore, as the area of aquatic bed expanded, the proportion of open water declined (Fig. 3). Following partial drawdowns in 2010 and 2012, due to pumping and a drought, respectively, mudflats expanded which provided habitat for migratory shorebirds (Smith et al., 2012; Fig. 3). Following these partial drawdowns, persistent emergent vegetation and nonpersistent emergent vegetation expanded. Generally, wet years favored the expansion of aquatic bed and hemi-marsh vegetation communities, whereas drier conditions encouraged the growth of nonpersistent emergent and persistent emergent vegetation communities. Clearly, hydrology was an important factor influencing vegetation community structure and cover type development, and increased water management capabilities in the future would enable management to target certain cover types or facilitate certain functions (Low & Bellrose, 1944; Fredrickson & Taylor, 1982; Bellrose et al., 1983).
Changes in vegetation structure observed at Emiquon were similar to those reported in previous research (Weller & Spatcher, 1965; Weller & Fredrickson, 1973; van der Valk & Davis, 1978). Van der Valk & Davis (1978) described four phases in the vegetation cycle of a prairie marsh: (1) dry marsh, (2) regenerating marsh, (3) degenerating marsh, and (4) lake marsh. During the dry marsh phase, mudflats are exposed allowing persistent emergent and nonpersistent emergent vegetation to germinate. In the regenerating phase, water returns to the marsh and terminates the germination of moist-soil plants and persistent emergent vegetation, while providing conditions favorable for submersed and floating-leaved aquatic vegetation. During the degenerating marsh phase, an abundance of submersed aquatic vegetation contributes to the seed bank as emergent vegetation declines due to a number of causes such as, disease, insects, high water, muskrat (Ondatra zibethicus L.) herbivory, and senescence. Finally, the lake marsh phase is defined by a dominance of submersed and floating-leaved aquatic vegetation, increasing muskrat populations, and persistent emergent vegetation confined to a narrow band around the perimeter. Eventually, submersed aquatic vegetation may decline due to increased turbidity from wave action, exacerbated by the loss of persistent emergent vegetation. Marshes remain in this climax state until the next drought or active drawdown to reset the cycle. During our study at Emiquon, we have witnessed parts of each phase of the vegetation cycle described by van der Valk & Davis (1978). At the start of restoration, persistent emergent and moist-soil vegetation flourished, but as water levels increased and were sustained during the growing seasons, hemi-marsh and aquatic bed vegetation communities expanded. In recent years of our study, we observed a dominance of submersed and floating-leaved aquatic vegetation, a decline in the hemi-marsh community, and an anecdotal increase in the number of muskrat lodges; thus, a substantial multiyear drawdown may be warranted in the near future as Emiquon continues to transition to the lake marsh phase.
Generally, obligate aquatic vegetation communities, such as aquatic bed and persistent emergent, were greater at Emiquon than during historical or contemporary periods in wetlands and lakes in the IRV. Although floating-leaved vegetation appeared less at Emiquon than historical periods, longleaf pondweed (Potamogeton nodosus Poir.) occurred extensively within the aquatic bed plant community concomitantly with submersed aquatic plants. Consequently, our estimate of floating-leaved vegetation is conservative because it was not practical to separately map longleaf pondweed. Contemporary wetlands in the IRV exhibited greater proportions of open water, nonpersistent emergent vegetation, and woody vegetation than those of Emiquon. In years when prolonged flooding does not occur during the growing season, most wetlands and lakes in the IRV are dewatered in mid-late summer to produce moist-soil vegetation, since hydrology prevents development of other plant communities (Stafford et al., 2010). Furthermore, bottomland forest and scrub-shrub communities have had little time to regenerate at Emiquon, following decades of row crop agriculture. Given the typical widespread development of moist-soil vegetation and lack of aquatic bed and persistent emergent vegetation communities, Emiquon fills an important niche in the IRV for migrating waterbirds and sportfish dependent upon these cover types for habitat (Hagy et al., 2016; VanMiddlesworth et al., 2016).
Stafford et al. (2010) noted a positive correlation between IJI value and total use days (UD) of mallards. Although our overall IJI value was slightly less than estimates reported by Stafford et al. (2010), Hagy et al. (2016) described that each incremental increase in IJI increased mallard use day density by approximately 16.5% (82.2 UD/ha) at Emiquon. The lower overall IJI value at Emiquon may have been attributable to the large proportion of the wetland area occupied by aquatic bed (44%) with less interspersion of other vegetation communities distributed throughout the basin. We propose that IJI values and waterbird use will decline concomitantly as Emiquon transitions towards the lake marsh stage described by van der Valk & Davis (1978).
Within aquatic bed, persistent emergent, and nonpersistent emergent vegetation communities, we observed trends among several invasive species. Curly pondweed increased annually until 2010, when we failed to find it on Emiquon, and encounters with the plant remained uncommon through 2013. Conversely, our encounters with Eurasian watermilfoil, reed canarygrass, and common reed increased throughout our study period. Eurasian watermilfoil increased rapidly throughout the aquatic bed and hemi-marsh communities during 2008–2012, but exhibited a slight decline in 2013. Similarly, Casper et al. (2016) showed Eurasian watermilfoil to be the dominant species in the aquatic bed community in 2012, but a decline may be related to competition with other species, such as coontail and longleaf pondweed. Encounters with reed canarygrass and common reed appear to be increasing despite some control efforts by TNC. Since a number of invasive species are now established, monitoring their response to a future drawdown will be important in case they out-compete native and desirable species.
Moist-soil plant seed density
The Upper Mississippi River and Great Lakes Region Joint Venture (UMRGLRJV) uses a moist-soil seed density estimate of 514 kg/ha for waterfowl conservation planning in the Upper Midwest (Soulliere et al., 2007; Stafford et al., 2011), which is similar to other regions important to nonbreeding waterfowl, such as the Mississippi Alluvial valley (550 kg/ha; Kross et al., 2008) and the Central Valley of California (634 kg/ha; Central Valley Joint Venture, 2006). Moist-soil plant seed densities at Emiquon were relatively high in 2007, the first year of restoration, and 2011, the year following a significant drawdown, exceeding published seed yields at Illinois Department of Natural Resources (IDNR) wetland management areas (mean 691 kg/ha; Stafford et al., 2011) and nearby Chautauqua National Wildlife Refuge (CNWR; mean 790 kg/ha; Bowyer et al., 2005; Table 2). During the two years when water levels increased from May to September (2008 and 2009), a period when drawdowns to encourage moist-soil plant growth typically occur, seed densities and EUDs were less than all other years. Nonetheless, seed and tuber densities from moist-soil plants surpassed the estimate used in the UMRGLRJV implementation plan in 6 of 7 years. Although Emiquon was not actively managed for production of moist-soil plant seeds and tubers as indicated by the relatively small proportion of nonpersistent emergent vegetation (mean 9%) compared to other contemporary IRV wetlands (mean 32%: Table 3), seed densities were comparable to those of more intensively managed moist-soil wetlands in the region. Emiquon exhibited the presence of a moist-soil seed bank and potential for expansion given proper hydrologic conditions.
Changing wetland conditions likely contributed to fluctuations in moist-soil plant seed yields during the period of study. The high yield observed at the start of restoration in 2007 was likely influenced by the extent of shallow surface and subsurface water around the perimeter of Thompson Lake during summer encouraging the growth of moist-soil plants. Moreover, recent soil tillage due to agricultural production resulted in nonpersistent emergent vegetation being dominated by early succession, annual species, which typically produce more seeds and tubers than late succession or perennial species (Naylor, 2002; Kross et al., 2008). Moist-soil plant seed and tuber density in 2011 were the highest recorded to date at the preserve (Table 2) when smartweeds (Polygonum L. spp), fall panicum (Panicum dichotomiflorum Michx.), and barnyardgrass (Echinochloa crus-galli [L.] P. Beauv.) produced an abundance of seed. An active drawdown through the use of pumps to repair infrastructure created large expanses of mudflats conducive for moist-soil plant development late in the growing season in 2010 and likely contributed to the abundant seed and tuber densities in 2011.
Drawdown timing to expose bare mudflats and encourage growth of persistent and nonpersistent emergent macrophytes may be extremely important at Emiquon due to the extensive aquatic bed plant communities, especially Eurasian watermilfoil, which develops early in the growing season (van der Valk & Davis, 1980; Fredrickson & Taylor, 1982; Bowyer et al., 2005; Gray et al., 2013). Drought conditions prevailed throughout the growing season in 2012 (IDNR, 2013), resulting in a 19% reduction in wet area from fall 2011. However, the drawdown occurred during mid- and late summer after submersed and floating-leaved aquatic vegetation developed. As the water receded in 2012, stranded aquatic vegetation covered mudflats and possibly prevented moist-soil plants and other macrophytes from becoming established. Timing of drawdown is a critical component of moist-soil plant development, especially in a wetland complex like Emiquon where growth and expansion of competing plant communities must be considered.
Emiquon exhibited the potential to produce abundant moist-soil vegetation, but the lack of water control (i.e., drawdown timing and extent, reflooding) likely precluded moist-soil plants from contributing much to the energetic needs of waterbirds in years when water levels were too high for seed germination or in years when precipitation was lacking and moist-soil vegetation was not reflooded. Water control capabilities would allow TNC to manage Emiquon more adaptively and would ultimately benefit each of the major vegetation communities that make up the wetland complex over time. As Emiquon moves toward the lake marsh phase (van der Valk & Davis, 1978), the ability to remove water in the absence of drought to create the dry marsh phase is critical to resetting the cycle and maintaining productive aquatic vegetation communities over time. The ability to mimic wet and dry cycles through water control allows managers to encourage growth of some vegetation communities while setting back other, more dominant communities, creating a more heterogeneous vegetation structure which is desirable to waterbirds (Kaminski & Prince, 1981).
Conclusion
During the past several decades, wetlands in the IRV incurred many anthropogenically induced changes which contributed to a reduction in wetland heterogeneity and loss of submersed and floating-leaved aquatic vegetation (Mills et al., 1966; Bellrose et al., 1983; Stafford et al., 2010). Bellrose et al. (1979) demonstrated that the greater the separation of a backwater lake from the Illinois River the more aquatic vegetation it produced. Jackson & Pringle (2010) warned of the negative consequences associated with hydrologic connectivity in highly modified river systems such as altered hydrology, exotic species, excessive nutrient levels, toxins, and sedimentation. They further suggested that reduced rather than enhanced hydrologic connectivity could produce greater ecological benefits in severely altered aquatic systems. Galat et al. (1998) proposed coupling intensively managed wetlands (isolated from the river by levees, less subject to flooding) with passively managed wetlands (connected to the river or more subject to flooding) to sustain the long-term health of the lower Missouri River ecosystem. New research has demonstrated the value of wetland complexes to wildlife (Pearse et al., 2012; Beatty et al., 2014), and our results illustrate significant tradeoffs wetland managers must weigh when targeting wetland functions (e.g., hydrologically connected wetlands for nutrient processing and sediment trapping versus disconnected and restored wetlands for wildlife; Sparks, 1995).
The substantial loss of aquatic vegetation in the IRV stresses the importance of Emiquon to wildlife dependent on these habitats. For instance, Hagy et al. (2016) and VanMiddlesworth et al. (2016) demonstrated the exceptional response of waterbirds and fishes, respectively, to aquatic vegetation communities at Emiquon Preserve during our study period. Floodplain wetlands isolated from the deleterious effects of the Illinois River (i.e., sedimentation, invasive species, and altered hydrology) have the potential to restore habitats that were once common in the Illinois valley and benefit many wetland wildlife species (Havera et al., 2003). Furthermore, little active management was required to restore mostly native aquatic plant communities at Emiquon, which is encouraging for future wetland restorations within large river system floodplains. Unfacilitated revegetation of wetland and deepwater areas of Emiquon is a testament to the resiliency of river-floodplain ecosystems. Emiquon may serve as a model for restoration of diverse aquatic plant communities in highly altered river systems. We suggest future research continue to examine the changes in vegetation communities at Emiquon as the system changes and recommend a means of water control to allow for adaptive management and the ability to create disturbances (e.g., drawdowns, flooding) to reset the marsh cycle. Eventually, perturbations to the system, such as multiyear drawdowns, will likely be needed to increase productivity and restore emergent vegetation communities.
References
Bajer, P. G., G. Sullivan & P. W. Sorenson, 2009. Effects of a rapidly increasing population of common carp on vegetative cover and waterfowl in a recently restored Midwestern shallow lake. Hydrobiologia 632: 235–245.
Beatty, W. S., E. B. Webb, D. C. Kesler, A. H. Raedeke, L. W. Naylor & D. D. Humburg, 2014. Landscape effects on mallard habitat selection at multiple spatial scales during the non-breeding period. Landscape Ecology 29: 989–1000.
Bellrose, F. C., 1941. Duck food plants of the Illinois River valley. Illinois Natural History Survey Bulletin 21: 235–280.
Bellrose, F. C., F. L. Paveglio Jr. & D. W. Steffeck, 1979. Waterfowl populations and the changing environment of the Illinois River valley. Illinois Natural History Survey Bulletin 32: 1.
Bellrose, F. C., S. P. Havera, F. L. Paveglio, Jr. & D. W. Steffeck, 1983. The fate of lakes in the Illinois River valley. Illinois Natural History Survey Biological Notes 119.
Bohm, W., 1979. Methods of studying root systems. Springer, Berlin.
Bowyer, M. W., J. D. Stafford, A. P. Yetter, C. S. Hine, M. M. Horath & S. P. Havera, 2005. Moist-soil plant seed production for waterfowl at Chautauqua National Wildlife Refuge, Illinois. American Midland Naturalist 154: 331–341.
Brock, M. A., D. L. Nielsen, R. J. Shiel, J. D. Green & J. D. Langley, 2003. Drought and aquatic community resilience: the role of eggs and seeds in sediments of temporary wetlands. Freshwater Biology 48: 1207–1218.
Casanova, M. T., 2012. Does cereal crop agriculture in dry swamps damage aquatic plant communities? Aquatic Botany 103: 54–59.
Casper, A. F., T. D. VanMiddlesworth, N. N. McClelland & G. G. Sass, 2016. Response of submerged aquatic vegetation to 7 years of floodplain restoration. Hydrobiologia (In Review).
Central Valley Joint Venture, 2006. Central Valley Joint Venture implementation plan – conserving bird habitat. U.S. Fish and Wildlife Service, Sacramento, CA.
Changnon, S. A. & A. Black, 2009. 2008: a record wet and stormy year in Illinois. Report of Investigation 117. Illinois State Water Survey, Champaign, IL.
Changnon, S. A., J. Angel & D. Kristovich, 2010. The unusual weather of 2009 in Illinois created major economic impacts. Report of Investigation 120. Illinois State Water Survey, Champaign, IL.
Cowardin, L. M., V. Carter, F. C. Golet & E. T. LaRoe, 1979. Classification of wetlands and deepwater habitats of the United States. U.S. Fish Wildlife Service FWS/OBS 79/31. U.S. Government Printing Office, Washington, DC.
Crook, S. L., W. C. Conway, C. D. Mason & K. J. Kraai, 2009. Winter time-activity budgets of diving ducks on eastern Texas reservoirs. Waterbirds 32: 548–558.
Dahl, T. E., 1990. Wetlands losses in the United States, 1780s to 1980s. U.S. Fish and Wildlife Service, Washington, DC.
Fredrickson, L. H. & T. S. Taylor, 1982. Management of seasonally flooded impoundments for wildlife. United States Department of the Interior, Fish and Wildlife Service Resource Publication 148, Washington, DC.
Galat, D. L., L. H. Fredrickson, D. D. Humburg, K. T. Bataille, J. R. Bodie, J. Dohrenwend, G. T. Gelwicks, J. E. Havel, D. L. Helmers, J. B. Hooker, J. R. Jones, M. F. Knowlton, J. Kubisiak, J. Mazourek, A. C. McColpin, R. B. Renken & R. D. Semlitsch, 1998. Flooding to restore connectivity of regulated, large-river wetlands. BioScience 48: 721–733.
Gray, M. J., H. M. Hagy, J. A. Nyman & J. D. Stafford, 2013. Management of wetlands for wildlife. In Anderson, J. T. & C. A. Davis (eds), Wetland Techniques, Vol. 3: Applications and Management: 121–180.
Greer, A. K., B. D. Dugger, D. A. Graber & M. J. Petrie, 2007. The effects of seasonal flooding on seed availability for spring migrating waterfowl. Journal of Wildlife Management 71: 1561–1566.
Hagy, H. M., J. N. Straub & R. M. Kaminski, 2011. Estimation and correction of seed recovery bias from moist-soil cores. Journal of Wildlife Management 75: 959–966.
Hagy, H. M., C. S. Hine, A. P. Yetter, M. M. Horath, R. V. Smith & J. D. Stafford, 2016. Response of waterbirds to wetland restoration in the Illinois River valley: a case study at Emiquon Preserve. Hydrobiologia (In Review).
Havera, S. P., A. P. Yetter, C. S. Hine & M. M. Georgi, 1995. Some misconceptions about conflicts between waterfowl and fisheries management. In Proceedings of the 51st Annual Meeting of the Upper Mississippi River Conservation Committee: 26–41.
Havera, S. P., 1999. Waterfowl of Illinois: status and management. Illinois Natural History Survey Special Publication 21: 436.
Havera, S. P., K. E. Roat & L. L. Anderson, 2003. The Thompson Lake/Emiquon story: the biology, drainage, and restoration of an Illinois River bottomland lake. Illinois Natural History Survey Special Publication 25.
Illinois Department of Natural Resources, 2013. The drought of 2012: a report of the governor’s drought response task force.
Irons, K. S., S. A. DeLain, E. Gittinger, B. S. Ickes, C. S. Kolar, D. Ostendorf, E. N. Ratcliff & A. J. Benson, 2009. Nonnative fishes in the Upper Mississippi River System. U.S. Geological Survey Scientific Investigations Report.
Jackson, C. R. & C. M. Pringle, 2010. Ecological benefits of reduced hydrologic connectivity in intensively developed landscapes. BioScience 60: 37–46.
Johnson, B. L. & C. A. Jennings, 1998. Habitat associations of small fishes around islands in the Upper Mississippi River. North American Journal of Fisheries Management 18: 327–336.
Kaminski, R. M. & H. H. Prince, 1981. Dabbling duck and aquatic macroinvertebrate responses to manipulated wetland habitat. Journal of Wildlife Management 45: 1–15.
Kaminski, R. M., J. B. Davis, H. W. Essig, P. D. Gerard & K. J. Reinecke, 2003. True metabolizable energy for wood ducks from acorns compared to other waterfowl foods. Journal of Wildlife Management 67: 542–550.
Kross, J., R. M. Kaminski, K. J. Reinecke, E. J. Penny & A. T. Pearse, 2008. Moist-soil seed abundance in managed wetlands in the Mississippi Alluvial valley. Journal of Wildlife Management 72: 707–714.
Lemke, M. J., A. F. Casper, H. M. Hagy & H. Chen, 2016a. Floodplain wetland restoration in the Midwest. In Ecological Restoration in the Midwest: Building on the Legacy. University of Iowa Press, Iowa City.
Lemke, A. M., J. Herkert., D. Blodgett & J. Walk, 2016b. Restoration planning: developing targets and key ecological attributes to guide ecological restoration & management. Hydrobiologia (In Review).
Low, J. B. & F. C. Bellrose, 1944. The seed and vegetative yield of waterfowl food plants in the Illinois River valley. Journal of Wildlife Management 8: 7–22.
Madsen, J. D., P. A. Chambers, W. F. James, E. W. Koch & D. F. Westlake, 2001. The interactions between water movement, sediment dynamics and submersed macrophytes. Hydrobiologia 444: 71–84.
McFarland, D. G. & S. J. Rogers, 1998. The aquatic macrophyte seed bank in Lake Onalaska, Wisconsin. Journal of Aquatic Plant Management 36: 33–39.
McGarigal K. & B. J. Marks, 1995. FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. General Technical Report PNW-GTR-351. USDA, Forest Service, Pacific Northwest Research Station, Portland.
McGarigal, K., S. A. Cushman & E. Ene, 2012. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. University of Massachusetts, Amherst [available on internet at http://www.umass.edu/landeco/research/fragstats/fragstats.html].
Mills, H. B., W. C. Starrett & F. C. Bellrose, 1966. Man’s effect on the fish and wildlife of the Illinois River. Illinois Natural History Survey Biological Notes 57.
Mitsch, W. J., L. Zhang, K. C. Stefanik, A. M. Nahlik, C. J. Anderson, B. Bernal, M. Hernandez & K. Song, 2012. Creating wetlands: primary succession, water quality changes, and self-design over 15 years. BioScience 62: 237–250.
Moore, M., S. P. Romano & T. Cook, 2010. Synthesis of upper Mississippi River system submersed and emergent aquatic vegetation: past, present, and future. Hydrobiologia 640: 103–114.
Naylor, L. W., 2002. Evaluating moist-soil seed production and management in Central Valley wetlands to determine habitat needs for waterfowl. Master’s Thesis. University of California–Davis.
Parrish, J. D., D. P. Braun & R. S. Unnasch, 2003. Are we conserving what we say we are? Measuring ecological integrity within protected areas. BioScience 53: 851–860.
Pavelis, G. A., 1987. Economic survey of farm drainage. In Farm drainage in the United States: history, status, and prospects. Miscellaneous Publication 1455. U.S. Department of Agriculture, Washington, DC.
Pearse, A. T., R. M. Kaminski, K. J. Reinecke & S. J. Dinsmore, 2012. Local and landscape associations between wintering dabbling ducks and wetland complexes in Mississippi. Wetlands 32: 859–869.
Raibley, P. T., D. Blodgett & R. E. Sparks, 1995. Evidence of grass carp (Ctenopharyngodon idella) reproduction in the Illinois and Upper Mississippi Rivers. Journal of Freshwater Ecology 10: 65–74.
Reinecke, K. J., R. M. Kaminski, K. J. Moorehead, J. D. Hodges & J. R. Nassar, 1989. Mississippi Alluvial valley. In Smith, L. M., R. L. Pederson & R. M. Kaminski (eds.), Habitat management for migrating and wintering waterfowl in North America. Texas Tech University Press, Lubbock: 203–247.
Smith, R. V., J. D. Stafford, A. P. Yetter, M. M. Horath, C. S. Hine & J. P. Hoover, 2012. Foraging ecology of fall-migrating shorebirds in the Illinois River valley. PLoS One 7(9): e45121. doi:10.1371/journal.pone.0045121.
Soulliere, G. J., B. A. Potter, J. M. Coluccy, R. C. Gatti, P. W. Brown, C. L. Roy Nielsen, M. W. Eichholz & D. R. Luukkonen, 2007. Upper Mississippi River and Great Lakes Region Joint Venture Waterfowl Habitat Conservation Strategy. U.S. Fish and Wildlife Service, Fort Snelling, MN.
Sparks, R. E., 1995. Need for ecosystem management of large rivers and their floodplains. BioScience 45: 168–182.
Sparks, R. E., J. C. Nelson & Y. Yin, 1998. Naturalization of the flood regime in regulated rivers. BioScience 48: 706–720.
Stafford, J. D., R. M. Kaminski, K. J. Reinecke & S. W. Manley, 2006. Waste rice for waterfowl in the Mississippi Alluvial valley. Journal of Wildlife Management 70: 61–69.
Stafford, J. D., M. M. Horath, A. P. Yetter, R. V. Smith & C. S. Hine, 2010. Historical and contemporary characteristics and waterfowl use of Illinois River valley wetlands. Wetlands 30: 565–576.
Stafford, J. D., A. P. Yetter, C. S. Hine, R. V. Smith & M. M. Horath, 2011. Seed abundance for waterfowl in wetlands managed by the Illinois Department of Natural Resources. Journal of Fish and Wildlife Management 2: 3–11.
Suloway, L. & M. Hubbell, 1994. Wetland resources of Illinois: an analysis and atlas. Illinois Natural History Survey Special Publication 15.
van der Valk, A. G. & C. B. Davis, 1978. The role of seed banks in the vegetation dynamics of prairie glacial marshes. Ecology 59: 322–335.
van der Valk, A. G. & C. B. Davis, 1980. The impact of a natural drawdown on the growth of four emergent species in a prairie glacial marsh. Aquatic Botany 9: 301–322.
VanMiddlesworth, T. D., N. N. McClelland & A. F. Casper, 2016. Emiquon fish community response to six years of restoration. Hydrobiologia. doi:10.1007/s10750-016-2696-8.
Walk, J. W., D. Blodgett, and A. M. Lemke, 2016. Emiquon: overview of a large-scale floodplain restoration. Hydrobiologia (In Review).
Weller, M. W. & C. E. Spatcher, 1965. Role of habitat in the distribution and abundance of marsh birds. Iowa State University, Agriculture and Home Economics Experiment Station Special Report 43.
Weller, M. W. & L. H. Fredrickson, 1973. Avian ecology of a managed glacial marsh. Living Bird 12: 269–291.
Wienhold, C. E. & A. G. van der Valk, 1989. The impact of duration of drainage on the seed banks of northern prairie wetlands. Canadian Journal of Botany 67: 1878–1884.
Yin, Y. & H. Langrehr, 2005. Multiyear synthesis of the aquatic vegetation component 1991– 2002 for the long term resource monitoring program. U.S. Geological Survey, La Crosse.
Acknowledgments
Funding for this project was provided by The Nature Conservancy’s Illinois River Project Office, contract number C07-032. We would like to thank D. Blodgett, J. Beverlin, T. Hobson, M. Lemke, S. McClure, J. Herkert, and J. Walk of The Nature Conservancy for their input and guidance in our research activities. We also appreciate our colleagues at the Illinois Natural History Survey’s Illinois River Biological Station for use of field and laboratory equipment. Finally, we thank G. Fretueg and K. Hardy for their assistance in the field and laboratory. All required permissions and permits were acquired to conduct this research in accordance with guidelines established by the University of Illinois and state and federal policies.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: Michael J. Lemke, A. Maria Lemke & Jeffery W. Walk / Large-Scale Floodplain Restoration in the Illinois River valley
Rights and permissions
About this article

Cite this article
Hine, C.S., Hagy, H.M., Horath, M.M. et al. Response of aquatic vegetation communities and other wetland cover types to floodplain restoration at Emiquon Preserve. Hydrobiologia 804, 59–71 (2017). https://doi.org/10.1007/s10750-016-2893-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-016-2893-5