
Abstract
Groundwater may create refuges for Atlantic salmon egg survival during low flows in regulated rivers and thus play an important role for survival during winter. To investigate the links between the survival of salmon embryos and hyporheic hydrological processes during permanent winter drawdown, a 100-m-long and 50-m-wide gravel area in a regulated river, the River Suldalslågen, was used for an experimental study. Surface and subsurface water levels were monitored with 2-min time resolution by means of water pressure sensors placed in pipes. Temperature, conductivity and dissolved oxygen were also measured. Eight cylindrical boxes, each with two compartments (at 10- and 30-cm depth, respectively) containing 50 Atlantic salmon eggs, were placed in the river bed substrate of both the drawdown zone and the permanently wetted area as a reference. They were regularly checked for survival during winter from January to May, coinciding with egg development period for this river. Survival rates in boxes in the dewatered river bed were between 8 and 78% during winter, compared to 80 to 99% in the reference wetted area. The main driver for egg survival in the dewatered area was groundwater with sufficient oxygen levels.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Groundwater may constitute 40–100% of the total discharge in inland Norwegian rivers during low flow periods in late summer and winter (Colleuille et al., 2005). The total groundwater inflow is usually lower, or shows higher variation in western coastal rivers, due to steeper topography and glacial-alluvial valley deposits of coarser sand and gravels of high permeability (Koestler & Brabrand, 2001). Geological heterogeneity will produce a potential for local underwater sites of groundwater flux, with heterogeneous hyporheic substrates (Hayashi & Rosenberry, 2002; Schmidt & Hahn, 2012) that will determine microspatial influx sites (Heggenes et al., 2010), thus creating spatial variability in habitat and spawning sites for salmonids with regard to flow, temperature and oxygen (Power et al., 1999). The hyporheic zone is an important component of the lotic ecosystem (Ward, 1989) with variable flux of groundwater and surface water, creating high vertical heterogeneity with associated ecological implications. Both spatial and temporal fine scale variabilities of hyporheic hydrochemistry, in particular dissolved oxygen, appear to be common and may affect eggs’ revival in gravel spawning salmonids (Greig et al., 2007; Soulsby et al., 2009; Malcolm et al., 2009).
Suitability of salmonid-spawning sites varies with specific hydraulic conditions and substrate (Peterson, 1978; Fleming, 1996), and in many cases, their accessibility increases with artificial high discharge providing access to areas that are not wetted during low flows (Bauersfeld, 1978; Chadwick, 1982). Salmonids often spawn in habitats where groundwater inflows occur, and their spawning success may be dependent on the limited availability of such habitats (Hansen, 1975; Garrett et al., 1998; Baxter & McPhail, 1999; Barlaup et al, 2008). Eggs have the most restrictive winter niche of all life stages of Atlantic salmon (Cunjack et al., 1998). Eggs of fall-spawning salmonids may freeze and die during natural or regulated low flow periods in late winter (Skoglund et al. 2012). However, where there is groundwater influx, freezing of eggs in winter might be minimized with the provision of more stable temperatures and flow and ice-free habitats (Cunjak et al, 1998; Heggenes et al., 2010; Saltveit & Brabrand, 2013). A significantly higher survival rate for salmonid embryos was documented in habitats influenced by discharging groundwater with a clear selection of those sites for spawning (Garrett et al., 1998; Baxter & McPhail, 1999). Such groundwater influence, given no oxygen deficiency (Soulsby et al., 2005), has also been observed in regulated rivers, where Atlantic salmon eggs may survive in groundwater-fed substrates for months during artificial winter drawdowns (Saltveit & Brabrand, 2013) or when exposed to hydropeaking (Casas-Mulet et al., 2014).
Physical and chemical conditions in redds can be altered when spawning areas are dewatered during winter (Young et al., 2011). The extent and duration of flow alteration and the stage of development will influence the survival, with eggs being more tolerant than the newly hatched alevins (Becker & Neitzel, 1985; Neitzel & Becker, 1985). If not subjected to extreme temperatures (in this case warm water) or predation, laboratory studies have confirmed that salmonid eggs may survive for weeks in dewatered gravel if they are kept moisty (Becker et al., 1982, 1983; Becker & Neitzel, 1985; Reiser & White, 1983). Survival in dewatered gravels has also been illustrated in earlier field studies (Saltveit & Brabrand, 2013; Casas-Mulet et al., 2014). The aim of our study was to investigate the survival of Atlantic salmon eggs in an area of potential spawning grounds that is permanently dewatered (with no detectable surface flow) during most of the egg incubation period due to regulation. Specific objectives were to identify and measure environmental variables directly influencing potential egg survival and to investigate their relationships during several periods of the egg incubation.
Materials and methods
Study site
The river Suldalslågen (Western Norway) runs 22 km from Lake Suldalsvatnet to the inner part of the Ryfylke fjord. Due to regulation, the flow in Suldalslågen is reduced, with an instream flow ranging between c. 12 and 65 m3 s−1, depending on the time of year. A stable minimum flow of 12 m3 s−1 is released in winter (15 December to 1 May), whilst higher flows and artificial floods are released during the rest of the year, for smolt migration, angling and fine sediment flushing purposes.
In the Suldalslågen, Atlantic salmon spawn relatively late compared to other Norwegian rivers, with a peak in early January (Heggberget, 1988). Based on models on egg and alevin development (Crisp, 1981, 1988; Jensen et al., 1989, 1991), ‘swim up’ should then occur between 17 June and 4 July. It, however, occurs one month earlier in this river according to in situ observations (Saltveit et al., 1995). This clearly indicates the importance of groundwater with higher and more stable temperatures compared to surface water during winter (Saltveit & Brabrand, 2013). Moreover, it supports the popular historical knowledge on connecting early hatching of juvenile Atlantic salmon to groundwater influx areas within the river (Anders Slagstad, pers. com.).
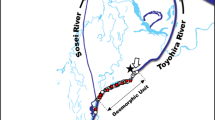
We selected the river Suldalslågen for this study given its low winter discharges both prior to, and due to regulation. With spawning occurring at higher flows, spawning redds can be dewatered, with egg mortality as a possible consequence of desiccation. An in situ incubation experiment was carried out 1 km below the Suldalsvatn dam. The study area was a 100-m-long and 50-m-wide side gravel bar on the southern side of the river (Fig. 1). Field experiments were undertaken from January to June 2012. The river discharge was stable during the study period, except for the two smolt migration floods in May, as illustrated in Fig. 2.

Illustration of the location of the sites in Suldalslågen and the experimental set-up

Water elevation and air temperature changes in the river Suldal, during the whole study period
In situ experimental set-up
Egg boxes and water quality
The experiments were conducted using eight cylindrical boxes, 24-cm height and 6.2-cm diameter, each divided into eight compartments. Eggs from Atlantic salmon were acquired from the local hatchery, fertilized 1 week ahead of the start of the experiments. Fifty eggs were placed in two of the box compartments: second from the top and bottom. These were protected with a 1-mm mesh net to avoid excess fine sediments and had a 0.5-m-long piece of surgical tubing connected to allow the extraction of water samples. The compartments above and below those with eggs were filled with small stones, pebbles and gravel from the river. All compartments were perforated with ten holes (diam. 5 mm) to allow water flux. Each cylinder was numbered and marked.
The cylindrical boxes were planted within the river bed on 18 January 2012 at four sites. One site (1) was located in the permanently wet area as reference for survival under permanent flow conditions. The other three sites (2–4) were located in the drawdown zone (Fig. 1). Two boxes were introduced at each site, c. 1 m apart, and placed in the river substrate, so that the uppermost compartment of each box was situated at the upper edge of the bed. When introduced, the dry river bed was covered by a 10-cm layer of snow, and the air temperature was −5°C.
Egg compartments were checked for survival and water samples were collected on three occasions during the egg incubation period: on 23 March (after period 1), 19 April (after period 2) and when terminating the experiments 24 May (after period 3). Dead eggs were removed to prevent fungal development. All eggs that hatched between April and May, including dead alevins, were considered as surviving the incubation period. Elevation changes due to reburial were recorded with a differential Global Positioning System (GPS; Topcon™ Legacy E+, Topcon Corporation, Japan) for each of the boxes at each of the control periods. When assessing survival, egg boxes were taken out of the river bed and reburied immediately to avoid disturbances. Water-quality samples were obtained pumping water from the egg compartments through the surgical tubing on four occasions (February, March, April and May). Oxygen, temperature, pH and conductivity were measured in situ with means of a WTW® Multi 3410 meter, and water samples were taken to the laboratory for turbidity analysis.
Water elevations, temperature and oxygen
Five Eijkelkamp® Diver water pressure transducers with integrated temperature loggers were inserted in pipes constructed of 32-mm inside diameter, Durapipe®. They were located next to each pair of egg boxes and provided 10-min resolution data on surface and subsurface water levels and temperatures in the ground (Fig. 1). One Eijkelkamp® Baro Diver was installed in the site to measure air temperature and air pressure to compensate for the absolute readings in the water pressure transducers.
Single-point water elevations were measured at the lowest and highest flows with a differential GPS and used as a reference to convert the continuous water-level data to elevations. Those were also linked to discharge data provided by the Norwegian Water Resources and Energy Directorate, NVE.
Substrate temperature was monitored at sites 2, 3 and 4 at 1-h time resolution from 7 February at 0.15 and 0.3 m below the ground (the level of the top and bottom compartments, respectively) by means of HOBO® temperature loggers installed next to each compartment. In box 2D, logging devices included an AADI® Datalogger 3634 with two optopodes measuring temperature and dissolved oxygen.
Geometry and grain size distribution
A high-resolution (10 cm of maximum separation between individual xyz points) geometrical characterization of the study area was obtained by means of laser scan (Topcon™ GLS-100, Topcon Corporation, Japan) in dry areas, combined with the use of differential GPS in wet areas. Two subsurface and two subsurface substrate samples were collected at the upstream (between Sites 2 and 3) and downstream (Site 4) areas of the drawdown zone in April during low flows. A 0.5 by 0.5 m wooden frame and coloured spray was used to separate the surface substrate by collecting only the painted gravels. Subsurface substrate was collected manually by shovelling out material at 30-cm depth inside the wooden frame. Samples were taken to the lab for particle size distribution analysis. It was obtained through a standard method of analysis by sieving and weighing, with sieve sizes of 0.075, 0.15, 0.3, 0.6, 1.18, 1.7, 2.36, 3.35, 4, 4.75, 6.3, 9.3, 12.5, 19, 25 and 37.5 mm. Cumulative granulometry curves were drawn to derive representative particle size ranges or D values.
Data analysis
Subsurface water elevation data at sites 2, 3 and 4 were used to calculate Vertical Hydraulic Gradients (VHGs) in relation to the surface water elevation in order to indicate the upward (positive VHG or upwelling potential) or downwards (negative VHG or downwelling potential) flow in the hyporheic zone (see also Hanrahan, 2008). Water elevations were compared to the elevation of each compartment. Durations of exposure to dry and dry and freezing conditions (maximum, total and number of occasions) were computed for each of the sampling periods.
Both the percentage of survival and the cumulative percentage of survival were calculated for each individual compartment and for each of the sampling periods.
A total of ten field-collected environmental variables (Table 3) were considered for statistical analysis. A correlation analysis was carried out to select non-correlated variables. These were then individually compared to survival rates through linear regressions to see if a single variable could explain survival. Further, in order to see whether combinations of variables could explain survival, only variables showing significant linear regressions were selected and combined into several GLM models. Each GLM model combination was tested and the best-fitted one was selected by using significance and Akaike’s information criterion (AIC).
Data analyses were carried out in Microsoft Excel and the software package R, version 2.14.1 (R Core Team, 2013). Sigma Plot version 12.0 was used for graphical presentations.
Results
Flow variations
The discharge released from the dam was very stable throughout most of the study period (Fig. 2). From 1 January until 30 April, the average flow was 13.6 m3 s−1 with very little variation, meaning that the eggs were never inundated with river surface water during that period (Fig. 1). On 1 May, when the discharge from the dam was increased from the first artificial spring flow of 40 m3 s−1, the water elevation increased by c. 40 cm, inundating the eggs placed in the dry river bed until the experiment was terminated on 24 May.
Environmental conditions
River water temperatures were fairly stable during the experimental period, increasing only from 2 to 5.5°C. The air temperature during the study period varied from −7.7°C on 1 February to 17°C when terminating the experiments in May, with several periods below 0°C in January and February (Fig. 2).
Granulometry characteristics in Suldalslågen are summarized in Table 1. Aggregates ranged from coarse gravel (D 90), fine and medium gravel (D 50) to coarse sand (D 10), generally with coarser surface materials in Sites 2–3. Fine sediments (<1 mm) represented a low percentage of the surface samples and were 17 and 11% in the subsurface samples of Sites 2–3 and 4, respectively.
Survival
The percentage of surviving eggs and the cumulative survival in each compartment and for each surveyed period is illustrated in Fig. 3. Total average survival and average survival by periods is summarized in Table 2, for both the reference sites 1 and sites 2–4 and for each of the compartments and the overall box.

Percentage of survival for the top and bottom compartments in each of the boxes for each of the three sampling periods. n.d. refers to periods with no data after a period of zero survival
High variability in survival between individual compartments and periods was observed. However, as expected, the reference site 1 showed very high average survival rates with a total average of 95.5% and up to 100% in period 1. The top compartments showed a slightly lower survival than the bottom compartments, but with less than 4% difference.
Sites 2–4 also gave high average survival rates of an overall 72.2%, with differences between the top and bottom compartments of 5%.
Water quality
The distribution values of the water quality data collected for each of the compartments is illustrated in Fig. 4. Dissolved oxygen varied from 6 to 14 mg l−1 and 60–110% between sites. Temperature variation (1–14°C) reflected the seasonal differences. Electrical conductivity values were between 6 and 90 µS cm−1 and pH between 6 and 8, both parameters having higher variability in the top compartments. The large spatial variation in electrical conductivity and pH in some of the compartments had no relevance for mortality compared with those having less variation. Turbidity was higher in the bottom compartments with values up to 400 NTU.

Distribution of the water quality variables values measured for each of the boxes (t top compartment, b bottom compartment) for all periods
Substrate temperatures in the boxes 1U and 1D, 2D, 3D and 4B were at all times above 0°C (Fig. 5). In the drained area, a vertical and lateral gradient in temperature changes was observed. Vertically, temperatures in the bottom compartments showed less variation, as expected. Larger fluctuations in temperature in the top compartments reflected a greater influence of air temperature. Within substrate, temperature showed minimal fluctuations in the wetted sites (1U and 1D), whilst the dewatered sites showed an increased influence of air temperature as they became further away from the river thalweg (4B).

Differences in temperatures between sites and between top and bottom compartments
Figure 6 shows continuous dissolved oxygen measurements at 2D top and bottom compartments, and several point measurements from the river. Dissolved oxygen levels in the subsurface water in the drained substrate were at all times lower than in the river. Changes in dissolved oxygen were directly linked to changes in groundwater level. However, the bottom compartments had higher dissolved oxygen concentrations than the top compartments during the majority of the low flow periods (except for very cold periods with temperatures below 0°C). In contrast, during the high flows in May, this is reversed with the top compartments having higher levels of dissolved oxygen indicating a greater influence of highly oxygenated surface water in the upper compartment areas. The dipping oxygen concentrations when the two flow peaks occur (Fig. 6), suggests that a different type of water, possibly older less rich in oxygen groundwater that was accumulated in the gravel, is mobilized during the peak, leading to a decreased oxygen concentration in the bottom compartments.

Continuous oxygen data from the top and bottom compartments of box 2D in comparison to point measurements in the river and the same compartments on four occasions
VHG and exposure to dewatering
During the low flow periods, a positive VHG at dewatered Sites 2 and 3 indicated upwelling potential (Fig. 7). Further downstream, at site 4, the negative VHG values indicated a downwelling potential. At high flows, VHG values were closer to zero, translating to a decrease of both upwelling and downwelling potentials.

Vertical hydraulic gradient (VHG) between the river and the subsurface water elevations at Sites 2, 3 and 4. VHG values presented are an average of the whole low flow and high flow period respectively. Upwelling potential is indicated by positive VHG values and downwelling potential by negative ones
The reference boxes at Site 1 (1U and 1D) were permanently covered by river surface water. The rest of the boxes, although located in an apparently dry area, had quite stable groundwater influx that prevented them from total desiccation. However, slight fluctuations in the groundwater influx and the relative position of the compartments in the study site, exposed some of them to desiccation, as shown in Fig. 8. Further, desiccation was combined with air temperatures below zero in some occasions, potentially leading to freezing. This is illustrated in Fig. 9, particularly for boxes 3D and 3U during period 1, with water levels below the top and bottom compartments when air temperatures were below zero.

Egg compartment elevations in comparison to water elevations for each of the sites. Ground temperatures are taken at the depth at which the piezometers were buried (see Fig. 1). Vertical lines denote sampling date. Egg compartment elevation may vary between sampling periods. No survival was measured in February

Duration of episodes with water levels above or below the egg compartment combined with air temperature above or below 0°C
Relationship between variables and survival
A correlation analysis was made between all the environmental variables considered on each compartment (Table 3). Total durations of exposure (to dry and dry and freezing conditions) and dissolved oxygen saturation levels were discarded in further analysis due to their high correlation (>95%) to maximum durations of exposure to desiccation and frost and dissolved oxygen concentration respectively.
The outputs (r 2 values and significance) of the individual linear regressions carried out are summarized in Table 4. Data are shown for the total dataset and for each of the periods. All regressions showed a normal distribution of the residuals. Several variables showed a significant relationship with survival rates during each of the individual periods and the overall experiment duration; however, only few were found with high r 2 values. Only three variables (turbidity, dissolved oxygen and maximum duration of exposure to dry conditions), showing a significant regression from the overall period analysis, were selected and combined into a total of 5 GLM models in order to understand their relationships with survival (Table 5). All models showed a normal distribution of the residuals. The best-fitted model was the combination of all three variables as it showed significance for all combinations and their interactions and the lowest AIC.
Discussion
Numerous spawning locations in regulated rivers are only accessible during limited high-flow periods with a potential dewatering of redds when the flow declines after spawning, potentially leading to high egg mortality due to desiccation or frost (Barlaup et al., 1994; Young et al., 2011; Skoglund et al., 2012; Vollset et al., in review). A relative high percentage of eggs, however, may survive in dewatered areas (Brabrand & Saltveit, 2013; Casas-Mulet et al., 2014). In the present study, both survival rates for each of the three periods and also for the whole study period were relatively high. Despite relatively high survival rates (72%), eggs in the drawdown compartments showed lower survival than those in the compartments permanently covered with surface river water (95.5%).
Variables having a significant individual effect on egg mortality included durations of exposure to desiccation and to desiccation and frost simultaneously, temperature, dissolved oxygen, electrical conductivity and turbidity. However, differences were found between periods with regard to such controlling parameters, and only maximum duration of exposure to desiccation, dissolved oxygen and turbidity showed a significant relationship for the whole of the study period. Vertical and horizontal differences in survival rates were also observed between and within egg boxes in the drawdown area. Such variabilities cannot be explained by a single variable. The GLM model combining these three variables was the best-fitted one to explain egg mortality in the Suldalslågen. However, spatial and temporal variabilities may have not been fully explained by the mode but by local factors such as streambed heterogeneity (e.g. Malard et al, 2002; Boulton, 2007) that can create a variable horizontal and vertical mosaic of interstitial flow, humidity, temperatures and dissolved oxygen in the river bed.
Subsurface water in Norway generally originates from very shallow aquifers in coarse river deposits, resulting in usually well-oxygenated groundwater (Brabrand et al., 2002). During low flows, the groundwater becomes more important for river water quality, including levels of dissolved oxygen. Except for the compartments 3Ub and 4Bb, the level of oxygen was never below 7 mg l−1 close to the egg boxes. In the River Lundesokna (mid-Norway), the level of oxygen in the river bed was never below 10 mg l−1 resulting in high egg survival (75–100%) during long dewatering periods due to hydropeaking (Casas-Mulet et al., 2014). No eggs survived in redds where average oxygen levels were less than 7 mg l−1 in Scottish rivers (Malcolm et al., 2003). Differences in reported critical values in dissolved oxygen could reflect differences in methods (including sampling frequency), salmonid species and water temperature between studies (Malcolm et al., 2002). Dissolved oxygen in bottom substrate plays a critical role in the development of the juvenile stages of benthic spawning fish and salmonids in particular. Factors influencing the dissolved oxygen regime within spawning gravels include the accumulation of fine sediment, penetration of groundwater or surface water into the gravels, the thermal regime and the consumption of oxygen by organic fractions in sediments (Jones et al., 2012).
Differences in local substrate composition and distribution might affect survival and this is illustrated from the high rates of survival in box 3D (top and bottom) during periods 1 and 2, in comparison to the high mortality in box 3U, only 1 m apart. However, substrate composition at such small spatial scale was not measured in this study, and only differences between upstream and downstream (through a representative sample) areas are shown. There are no indications that the effect of substrate manipulation during sampling had effects on survival, as great care was taken and the lapse of time between sampling periods was long enough to allow recovery between periods. In addition, no effects were detected in the river Lundesokna (see Casas-Mulet et al., 2014), where the same methods were applied. The relative position of some of the compartments to the slightly fluctuating groundwater elevation and then the duration of exposure to desiccation and freezing might explain the high mortality in period 1 in some of the compartments such as the tops of 3U, 2U and 2D, which were exposed to long-lasting desiccation and frost periods. However, other compartments were equally exposed during this period, and showed high survival, such as 3D and 4T tops (100 and 95% survival, respectively, in period 1). Therefore, micro-scale local conditions of groundwater influence, clogging and dynamic processes occurring in the compartments, including possible variation between eggs in incubation time, although there is no evidence of strong intraspecific variation in incubation time and egg survival within Atlantic salmon populations (Jonsson & Jonsson, 2011).
Despite the bottom compartments being never exposed to dry or frost condition, the final survival rates in these compartments of 4B, 3U and 3D were 0, 0 and 18%, respectively. Turbidity levels were relatively high, and in addition, large amounts of fine organic sediments were noticed inside these compartments during sampling, potentially leading to critically low dissolved oxygen levels in the micro environment close to the eggs not detected from the water samples or the loggers. As such, high level of turbidity is probably not a mortality factor if not settling on egg surface preventing oxygen supply. Similar high turbidity level were found not only in the top compartments 2U and 4B, with low survival rates of 36 and 29%, respectively, but also in the bottom compartments 2D and 2U with high survival rates (74 and 82% respectively). This variability in results illustrates that the local conditions around the boxes and the complex groundwater dynamics affecting each compartment may have affected the final results. Fine sediments were not the primary factor determining within-redd mortality rates in the Newmills Burn (Soulsby et al., 2001). However, variations of only a few percent of silt content can strongly decrease survival to emergence (Lapointe et al., 2005). Increasing hydraulic gradients has a positive effect on median survival, but the effect depends both on sediment composition and the height gradient. There is no single threshold interstitial flow velocity that ensures survival to emergence. Even when maintaining a constant interstitial velocity, survival tended to be reduced in substrate with a finer content (Olsson & Persson, 1986; 1988; Lapointe et al., 2005).
Suitable reproductive habitats are a prime necessity for population sustainability, and river regulation may reduce the abundance and quality of spawning habitat, thus directly affecting recruitment of salmonid populations. Even though the importance of groundwater for salmon redd site selection and egg survival appears obvious (Soulsby et al., 2005), and that the use of groundwater upwelling sites for spawning has been reported for several salmonid species (e.g. Garett et al., 1998), there is little data to substantiate the idea that groundwater outflows directly affect spawning site selection (Baxter & McPhail, 1999). Varying patterns of interactions between groundwater and river surface water may generate a spatial and temporal mosaic and consequently complex conditions for egg survival (Malcolm et al., 2009), egg development and spawning time. During low flow periods in regulated rivers, there may therefore be an increase in the relative importance of groundwater for salmonid survival.
Conclusions
A certain proportion of Atlantic salmon eggs located in dewatered redds can survive during winter even when covered with ice and snow. However, this survival is lower in comparison to survival in permanently wetted locations.
Survival rate of eggs in the dewatered redds can vary with both the relative horizontal position along the gravel bar. The main drivers for survival were found to be linked to groundwater influx with regard to water level and water-quality characteristics such as oxygen and turbidity as a potential indicator of fine sediments.
Such findings are important for the management of regulated rivers by emphasizing the importance of considering groundwater influx when assessing the management needs for the conservation of Atlantic salmon populations.
References
Baxter, J. S. & J. D. McPhail, 1999. The influence of redd site selection, groundwater upwelling, and over-winter incubation temperature on survival of bull trout (Salvelinus confluentus) from egg to alevin. Canadian Journal of Zoology 77: 1233–1239.
Barlaup, B. T., H. Lura, H. Sagrov & R. C. Sundt, 1994. Inter- and intra-specific variability in female salmonid spawning behaviour. Canadian Journal of Zoology 72: 636–642.
Barlaup, B. T., S. E. Gabrielsen, H. Skoglund & T. Wiers, 2008. Addition of spawning gravel-a means to restore spawning habitat of Atlantic salmon (Salmo salar L.), and anadromous and resident brown trout (Salmo trutta L.) in regulated rivers. River Research and Applications 24: 543–550.
Bauersfeld, K. 1978. Stranding of juvenile salmon by flow reductions at Mayfield Dam on the Cowlitz River, 1976 Report. Washington State Department of Fisheries, Olympia: 36.
Becker, C. D. & D. Neitzel, 1985. Assessment of intergravel conditions influencing egg and alevin survival during salmonid redd dewatering. Environmental Biology of Fishes 12: 33–46.
Becker, C. D., D. Neitzel & D. H. Fickeisen, 1982. Effects of dewatering on Chinook salmon redds: tolerance of four developmental phases to daily dewaterings. Transactions of the American Fisheries Society 111: 624–637.
Becker, C. D., D. Neitzel & C. S. Abernethy, 1983. Effects of dewatering on Chinook salmon redds: tolerance of four development phases to one-time dewatering. North American Journal of Fisheries Management 3: 373–382.
Boulton, A. J., 2007. Hyporheic rehabilitation in rivers: restoring vertical connectivity. Freshwater Biology 52: 632–650.
Brabrand, Å., A. G. Koestler & R. Borgstrøm, 2002. Lake spawning of brown trout related to groundwater influx. Journal of Fish Biology 60: 751–763.
Casas-Mulet, R., S. J. Saltveit & K. Alfredsen, 2014. The Survival of Atlantic Salmon (Salmo salar) Eggs During Dewatering in a River Subject to Hydropeaking. River Research and Applications. Early view, New York.
Chadwick, E. M. P., 1982. Stock-recruitment relationship for Atlantic salmon (Salmo salar) in Newfoundland Rivers. Canadian Journal of Fisheries and Aquatic Sciences 39: 1496–1501.
Colleuille, H., P. Dimakis & W.K. Wong, 2005. Elv og grunnvann. Sluttrapport-Oppsummering og anbefalinger. NVE rapport Miljøbasert vannføring 8-2005: 39 (in Norwegian).
Crisp, D. T., 1981. A desk study of the relationship between temperature and hatching time for the eggs of five species of salmonid fishes. Freshwater Biology 11: 361–368.
Crisp, D. T., 1988. Prediction, from temperature, of eyeing, hatching and “swim up” times for salmonid embryos. Freshwater Biology 19: 41–48.
Cunjak, R. A., T. D. Prowse & D. L. Parrish, 1998. Atlantic salmon (Salmo salar) in winter: the season of parr discontent? Canadian Journal of Fisheries and Aquatic Sciences, Supplement 1(55): 161–180.
Fleming, I. A., 1996. Reproductive strategies of Atlantic salmon: ecology and evolution. Reviews in Fish Biology and Fisheries 6: 379–416.
Garrett, J. W., D. H. Bennett & L. R. Clarke, 1998. Enhanced incubation success for kokanee spawning in groundwater upwelling sites in a small Idaho stream. North American Journal of Fisheries Management 18: 925–930.
Greig, S. M., D. A. Sear & P. A. Carling, 2007. A review of factors influencing the availability of dissolved oxygen to incubating salmonid embryos. Hydrological Processes 21: 323–334.
Hanrahan, T. P., 2008. Effects of river discharge on hyporheic exchange flows in salmon spawning areas of a large gravel-bed river. Hydrological Processes 22(1): 127–141.
Hansen, E. A., 1975. Some effects of groundwater on brown trout redds. Transactions of the American Fisheries Society 104: 100–110.
Hayashi, M. & D. O. Rosenberry, 2002. Effects of groundwater exchange on the hydrology and ecology of surface water. Groundwater 40: 309–316.
Heggberget, T. G., 1988. Timing of spawning in Norwegian Atlantic salmon (Salmo salar). Canadian Journal of Fisheries and Aquatic Sciences 45: 845–849.
Heggenes, J., G. Bremset & Å. Brabrand, 2010. Groundwater, critical habitats, and behavior of Atlantic salmon, brown trout and Arctic char in streams. NINA Report 654: 28 pp.
Jensen, A. J., B. O. Johnsen & L. Saksgård, 1989. Temperature requirements in Atlantic salmon (Salmo salar), brown trout (Salmo trutta), and Arctic char (Salvelinus alpinus) from hatching to initial feeding compared with geographic distribution. Canadian Journal of Fisheries and Aquatic Sciences 46: 786–789.
Jensen, A. J., B. O. Johnsen & T. G. Heggberget, 1991. Initial feeding time of Atlantic salmon, Salmo salar, alevins compared to river flow and water temperature in Norwegian streams. Environmental Biology of Fishes 30: 379–385.
Jones, J. I., J. F. Murphy, A. L. Collins, D. A. Sear, P. S. Naden & P. D. Armitage, 2012. The impact of fine sediment on macro-invertebrates. River Research and Applications 28: 1055–1071.
Jonsson, B., Jonsson, N. 2011. Ecology of Atlantic salmon and brown trout. In Habitat as a Template for Life Histories. Springer, Dordrecht: 701. doi:10.1007/978-94-007-1189-1.
Koestler, A. G. & Å. Brabrand, 2001. Grunnvann som mulig årsak til mislykkede rotenonbehandlinger. Vann 1: 29–35. (in Norwegian).
Lapointe, M. F., N. E. Bergeron, F. Bérubé, M. A. Pouliot & P. Johnston, 2005. Interactive effects of substrate sand and silt contents, redd-scale hydraulic gradients, and interstitial velocities on egg-to-emergence survival of Atlantic salmon (Salmo salar). Canadian Journal of Fisheries and Aquatic Sciences 61: 2271–2277.
Malard, F., K. Tockner, M. J. Dole-Olivier & J. V. Ward, 2002. A landscape perspective on surface-subsurface hydrological exchanges in river corridors. Freshwater Biology 47: 621–640.
Malcolm, I. A., C. Soulsby & A. F. Youngson, 2002. Thermal regime in the hyporheic zone of two contrasting salmonid spawning streams: ecological and hydrological implications. Fisheries Management and Ecology 9: 1–10.
Malcolm, I. A., A. F. Youngson & C. Soulsby, 2003. Survival of salmonid eggs in a degraded gravel-bed stream: effects of groundwater–surface water interactions. River Research and Applications 19: 303–316.
Malcolm, I. A., C. Soulsby, A. F. Youngson & D. Tetzlaff, 2009. Fine scale variability of hyporheic hydrochemistry in salmon spawning gravels with contrasting groundwater–surface water interactions. Hydrogeology Journal 17: 161–173.
Neitzel, D. A. & C. D. Becker, 1985. Tolerance of eggs, embryos, and alevins of Chinook salmon to temperature changes and reduced humidity in dewatered redds. Transactions of the American Fisheries Society 114: 267–273.
Olsson, T. I. & B. Persson, 1986. Effects of gravel size and peat material concentrations on embryo survival and alevin emergence of brown trout, Salmo trutta L. Hydrobiologia 135: 9–14.
Olsson, T. I. & B. Persson, 1988. Effects of deposited sand on ova survival and alevin emergence in brown trout (Salmo trutta L.). Archiv für Hydrobiologie 113: 621–627.
Peterson, R.H., 1978. Physical characteristics of Atlantic salmon spawning gravel in some New Brunswick streams. Fisheries Marine Service Technical Report 785: 28 pp.
Power, G., R. S. Brown & J. G. Imhof, 1999. Groundwater and fish—insights from northern North America. Hydrological Processes 13: 401–422.
R Core Team, 2013. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0 [http://www.R-project.org/]. Accessed May 2013.
Reiser, D. W. & R. G. White, 1983. Effects of complete redd dewatering on salmonid egg-hatching success anddevelopment of juveniles. Transactions of the American Fisheries Society 112: 532–540.
Saltveit, S. J., T. Bremnes & O. R. Lindaas, 1995. Effect of sudden increase in discharge in a large river on newly emerged Atlantic salmon (Salmo salar) and brown trout (Salmo trutta) fry. Ecology of Freshwater Fish 4: 168–174.
Saltveit, S. J. & Å. Brabrand, 2013. Incubation, hatching and survival of eggs of Atlantic salmon (Salmo salar) in spawning redds influenced by groundwater. Limnologica 43: 325–331.
Skoglund, H., B.T. Barlaup, S.E. Gabrielsen, G.B. Lehmann, G.A. Halvorsen, T. Wiers, B. Skår, U. Pulg & K.W. Vollset, 2012. Fiskebiologiske undersøkelser i Eidfjordvassdraget—sluttrapport for perioden 2004–2012. LFI-Unifob rapport 203: 108 (in Norwegian).
Schmidt, S. I. & H. J. Hahn, 2012. What is groundwater and what does this mean to fauna?—An opinion. Limnologica—Ecology and Management of Inland Waters 42: 1–6.
Soulsby, C., A. F. Youngson, H. J. Moir & I. A. Malcolm, 2001. Fine sediment influence on salmonid habitat in a lowland agricultural stream: a preliminary assessment. Science of the Total Environment 265: 295–307.
Soulsby, C., I. A. Malcolm, A. F. Youngson, D. Tetzlaff, C. N. Gibbins & D. M. Hannah, 2005. Groundwater–surface water interactions in upland Scottish rivers: hydrological, hydrochemical and ecological implications. Scottish Journal of Geology 41: 39–49.
Soulsby, C., I. A. Malcolm, D. Tetzlaff & A. F. Youngson, 2009. Seasonal and inter-annual variability in hyporheic water quality revealed by continuous monitoring in a salmon spawning stream. River Research and Applications 10: 1304–1319.
Vollset, K., B. Barlaup, H. Skoglund, S. Gabrielsen & T. Wiers. 2014. Effects of hydropeaking on the spawning behaviour of Atlantic salmon (Salmo salar) and brown trout (Salmo trutta). Manuscript submitted for publication.
Ward, J. V., 1989. The four-dimensional nature of lotic ecosystems. Journal of the North American Benthological Society 8: 2–8.
Young, P., J. Cech & L. Thompson, 2011. Hydropower-related pulsed-flow impacts on stream fishes: a brief review, conceptual model, knowledge gaps, and research needs. Reviews in Fish Biology and Fisheries 21: 713–731.
Acknowledgments
The Research Program Centre for Environmental Design of Renewable Energy (CEDREN)—financed by major Norwegian energy companies and the Research Council of Norway under RCN Contract 201779—provided the financial support. The authors would like to thank Sigmund Vårvik at the Suldal River Owner’s Association, for providing the eggs for this study, John E. Brittain for comments and language improvement, and Prof. Sigurd Einum for his suggestions on statistics interpretation. The authors are also grateful to the three anonymous reviewers for their constructive comments and help they provided during the editorial process.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Odd Sandlund
Rights and permissions
About this article

Cite this article
Casas-Mulet, R., Alfredsen, K., Brabrand, Å. et al. Survival of eggs of Atlantic salmon (Salmo salar) in a drawdown zone of a regulated river influenced by groundwater. Hydrobiologia 743, 269–284 (2015). https://doi.org/10.1007/s10750-014-2043-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-014-2043-x