
Abstract
The success of predators in species invasion will depend on their interactions with their own predators and competitors. The present study examined whether the predation of piscivorous fish among other piscivorous fish could be an active mechanism in species invasion. The diet of eleven piscivorous fish found in the upper Paraná River basin was analyzed during eight years. Seven of the fish species were native to the river basin, and four were invasive species. The diet composition of the studied species did not differ from each other, and a high value of niche overlap was found among invasive species with native species. Invasive species consumed higher amounts of piscivorous species, mainly Hoplias sp. 1 and C. kelberi. Salminus brasiliensis was the only native species with high values of predation over piscivorous fish. There were no significant differences between the consumption of piscivorous fish and their abundance. Overall, the consumption of piscivorous fish by invasive species can act as a mechanism for their success and maintenance in a new environment. Piscivorous invaders radically change the composition of their new environment more than other trophic levels; therefore, we recommend special care with the introduction of piscivores.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
In freshwater environments, invasive species are known to cause deleterious effects on diversity (Zaret & Paine, 1973; Pelicice & Agostinho, 2009) and to induce dramatic changes in community structure (Sharma et al., 2011) and functioning in assemblages (Baxter et al., 2004). The success of an invasive species is determined by its ability to tolerate and overcome the various pressures of the new environment (Diamond & Case, 1986), including direct interactions, such as competition and predation with native species (Dick, 1996; MacNeil & Prenter, 2000).
In aquatic systems, piscivorous invaders most likely change the composition of the novel area more radically than omnivorous or detritivorous invaders (Moyle & Light, 1996). Piscivorous species are relatively easy to introduce due to their aggressive behavior, and such invasions are recognized as one of the most powerful biological ways of transforming native communities (Luiz et al., 2011). Most predators have multispecific diet in a community (Hurd & Eisenberg, 1990; Sih et al., 1998) and this linkage results in several potential types of interactions between two coexisting predators species within an assemblage: (i) non interactive predators, (ii) predators that attack and kill each other, (iii) predators that influence either the foraging behavior or foraging range of another predator, and (iv) predators that influence prey behavior and thus affect prey susceptibility to other predators (Losey & Denno, 1998).
One key factor that likely governs emergent multipredator effects is an overlap between predators in their foraging habitat (Hochberg, 1996). The coexistence of two predators and their prey only occurs if one of the predators is a better competitor for the prey than the other (Sih et al., 1998). If the native predator species consume the invaders at a rate greater than or equal to its rate of arrival, it can prevent the establishment of a non-native species (Shinen et al., 2009). Thus, the success of predator invasions will depend on their interactions with their own predators and competitors (Sih et al., 2010).
Top predators may switch more readily from foraging on shared prey to foraging on intermediate predators in habitats where intermediate predators are more vulnerable to predation (Griffen & Byers, 2006). When a native species and an invasive species compete for the same feeding resource (prey), the invasive species can compensate this resource loss by consuming its competitors, which could increase the colonization success (Hall, 2011). However, identifying the main mechanisms that determine the success and establishment of invasive species is still a challenge in invasion ecology (Rice & Silverman, 2013). Thus, the objective of this study was to evaluate the consumption between native and non-native piscivorous fish. The working hypothesis was that invasive species will have higher consumption rates of their competitors than native piscivorous fish. If our hypothesis is supported, we can state that “eating the competition” may be a facilitator mechanism for the invasion of non-native predators.
Materials and methods
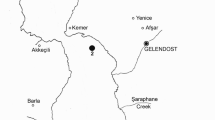
The upper Paraná River floodplain is located between the coordinates 22–22º50′ S and 53º15′–53º40′ W, in the third inferior part of the upper Paraná River (Fig. 1). There are approximately 182 fish species in the study area and, of this total, more than 50 species are non-native (Graça & Pavanelli, 2007). The origins of non-native species are diverse, including fish farming, sport fishing or facilitation by damming (Langeani et al., 2007). The upper Paraná is characterized by high diversity and endemic fauna, which is highly threatened due to hydrological alterations and species invasion (Agostinho et al., 2008). The floodplain where the study was conducted is the only remnant of the upper Paraná River with lotic waters within the Brazilian territory.

Locations of sampling stations in the upper Paraná River floodplain, downstream from the Porto Primavera Dam
Samples were made in three distinct rivers of the floodplain (Baía, Ivinhema and Paraná rivers) totalizing 36 sampling stations (12 in each river) in different environments, including connected and non-connected lakes, channels, and river main channel. Fish species were captured quarterly from March 2005 to June 2013. Gill nets of different mesh sizes were used to capture fish (2.4–16 cm between opposite knots). The nets were exposed for a 24-h period and checked at 8:00 AM, 4:00 PM, and 10:00 PM. All sampled fish were identified, measured and eviscerated, and their stomachs were removed and preserved in 10% formalin for later analysis.
Fish stomach contents were examined under a stereoscopic microscope to determine their diets, and items were identified to the lowest possible taxonomic level. Fish were identified using the keys published by Graça & Pavanelli (2007). Fish preys were measured and the volumetric (Hyslop, 1980) and occurrence (Hynes, 1950) methods were used to express diet results.
The piscivorous fish species that had more than 40 adult individuals with content in the stomachs were analyzed and classified as native or invasive according to Pazza & Júlio-Jr (2003), Langeani et al. (2007), and Júlio-Jr et al. (2009). The native species were Acestrorhynchus lacustris (Lütken, 1875), Hoplias sp. 2, Hoplias sp. 3, Hemisorubim platyrhynchos (Valenciennes, 1840), Pseudoplatystoma corruscans (Spix & Agassiz, 1829), Rhaphiodon vulpinus (Spix & Agassiz, 1829), and Salminus brasiliensis (Cuvier, 1816), which correspond to 40% of the total native piscivorous abundance in the study area. The invasive species were Cichla kelberi Kullander & Fereira, 2006, Cichla piquiti Kullander & Fereira, 2006, Hoplias sp. 1 and Plagioscion squamosissimus (Heckel, 1840), representing more than 70% of the total abundance of invasive piscivores.
Feeding items were grouped taxonomically, at family and subfamily (for Characidae) levels, for analysis because they may reflect the shared ecomorphological and functional patterns of the prey species (Oliveira et al., 2010). Serrasalmus marginatus (Valenciennes, 1837) and Hoplerithrynus unitaeniatus (Agassiz, 1829) were considered as prey only because of the difficulty in analyzing S. marginatus stomachs that contained prey pieces that were hard to identify at a specific level and because of the low capture of H. unitaeniatus.
The proportion of fish consumed was obtained for each predator species to confirm the piscivorous diet. A Chi-square test was used to determine if the frequency of piscivorous fish consumption differed between native and invasive species, using as null hypothesis that the piscivorous consumption between native and invasive fish did not differ.
To calculate the feeding overlap between native and invasive species, the Schoener’s index (Schoener, 1970) was applied to the grouped data with all food resources. The significance of the overlapping was tested using a Chi-square test between the pair of species. The Schoener’s index is described as:
where, \( C_{xy} \) is the Schoener’s niche overlap Index and pxy and p yi are the estimated proportions of prey i in the diets of species x and y, respectively. This index ranges from 0 (no dietary overlap) to a maximum of 1 (all prey items are found in equal proportions).
A principal coordinate analysis (PCoA) was used with a Bray-Curtis similarity resemblance matrix (Legendre & Legendre, 1998) to summarize data on the diet of the native and invasive piscivorous species. A Permutational Analysis of Multivariate Dispersions (PERMDISP; Anderson, 2004) was used to test for differences between native and non-native species in the ordination, based on the distances of the samples relative to the group average (Anderson, 2006). This analysis determines if the diet is more variable among native or invasive species and if the group diets’ differ from each other. These analyses were tested by a Monte Carlo method with 999 randomizations.
To determine if the piscivore consumption was due to the abundance of available preys, the percentage of consumable preys was calculated for the environments, considering only the prey species and size range that were found in the stomachs; species or lengths not found in the stomachs content were omitted as they were deemed inaccessible to the studied species. Finally, to determine if the frequency of piscivorous consumption differences were due to abundance variations of the species, a Spearman’s correlation was used to analyze the associations between abundance (catch per unit of effort—CPUE; unit: number of individuals/1,000 m2 of nets set for 24 h) and the frequency of piscivorous fish consumption.
All analyses were performed with the R Programming Environment software, with the packs Vegan for PCoA and PERMDISP and spa for niche overlap (The R-Project for Statistical Computing, http://www.r-project.org/). The graphics were constructed in Statistica Statsoft 10.0.
Results
As expected, a greater proportion of fish was found in the stomach content of the piscivorous species, with some variation in the intensity of consumption. Most of the species showed at least 80% of their total diet composed of fish, while the invasive C. piquiti consumed mostly other items, such as invertebrates, having only 39% of its diet composed of fish. Plagioscion squamosissimus and S. lima had an intermediate consumption of fish (70%), while the rest of their diets were dominated mostly by shrimp (Table 1). Native and invasive species presented high feeding overlap (0.73). However, the Chi-square test did not find significant differences between native and invasive and species feeding overlap (χ 2 = 3.27, df = 10, P = 0.97).
In total, the analyzed fish fed on 24 fish families. Characidae was consumed by all analyzed species, and C. piquiti was the only species that did not feed on Cheirodontinae and Erythrinidae. Anostomidae, Characidae, Cheirodontinae, Curimatidae, Serrasalminae, Sternopygidae and Cichlidae were the most consumed items while Apteronotidae, Hemiodontidae, Poeciliidae and Aphyocharacinae were the least consumed. The PCoA summarized the diet of the piscivorous fish. In the ordination, it is possible to verify three distinct groups of points, which are related by the fact that usually a single individual was registered in the stomachs of piscivores (Fig. 2). The preys consumed by each group belong to the same family, as fish do not select, resulting in the formation of the groups. But, besides the separation in groups, according to the ingested families, all of the analyzed species can be found in the three different groups. As a result of this, the PERMDISP did not find significant differences in the diet of native and invasive species (F = 1.33, P = 0.25) (Fig. 2).

Diet data ordination from native species and invasive species represented by the two axes of a principal coordinate analysis (PCoA 1 and PCoA 2)
Significant differences were found for the frequency of consumption of piscivorous fish between native and invasive species (χ 2 = 30.54, df = 9, P < 0.001). Generally, invasive species had 15% of their total diet dominated by piscivorous fish, while native species had approximately 10% of their total diet dominated by piscivorous fish. The species that were the most consumed by native species were C. kelberi and Hoplias. Cichla kelberi was also the most consumed by invasive species followed by S. marginatus (Fig. 3). The species that had the highest proportion of piscivorous fish consumption (almost 30%) was the invasive Hoplias sp. 1; Cichla kelberi had the second highest proportion (17%), but 11% were individuals of the same species. The native species, in general, had lower consumption of piscivores, with values lower than 10% of the total diet, except for S. brasiliensis with 26% of its total diet, of which 13% was composed by the invasive C. kelberi. The invasive C. piquiti was the only species that did not present any consumption of piscivorous fish and S. brasiliensis and R. vulpinus were not consumed by any of the predators (Table 2).

Percentage of piscivorous prey in the diet of native and invasive species
The availability of preys in the environments was high, at least twice of the piscivorous abundance, even for native or invasive piscivores (Fig. 4A). Thus, prey availability does not appear to be a determinant factor in the study area. The most abundant piscivorous species in the environment were A. lacustris, C. kelberi, Hoplias sp. 1, and Hoplias sp. 2 (Fig. 4B). The Spearman correlation coefficients between the frequency of piscivorous fish predation and species abundance were not significant (Native ρ = −0.06, P = 0.86; Invasive ρ = −0.02, P = 0.52), showing that the piscivorous fish consumption is independent of the piscivorous fish abundance for either native or invasive species.

Abundance (Capture per unit of effort = CPUE – unit: number of individual/1,000 m2 of net set for 24 h) of the analyzed piscivorous species. A Mean CPUE of the total analyzed species and the available preys in the environment. B Mean CPUE of each analyzed species in the environment. Vertical bars = Standard error
Discussion
In this study, native species and invasive species did not present significant differences in their diets, which indicate that invasive species are consuming the same feeding resources as native species. The high values of feeding overlap between native and invasive species corroborate this result. When a species invade a new environment, its successful establishment is reduced by the presence of competitors (Okubo et al., 1989; Svenning et al., 2014). However, invaders may compensate for food resources lost by direct consumption of competitors. This direct consumption of competitors may influence invasion speeds (Hall, 2011).
Hoplias sp. 1 had the highest piscivorous fish consumption among all studied species. This species was introduced in the upper Paraná River floodplain after the construction of Itaipu Dam (Pazza & Júlio-Jr, 2003). Currently, its abundance is nearly the same as its congener Hoplias sp. 2 and is the third in abundance in the floodplain of all the analyzed species. This great increase in abundance might be related to its high aggressiveness (showed by the high predation values), especially when considering that aggressiveness and voracity are basic behaviors that promote superiority competition among species (Holway & Suarez, 1999; Rehage & Sih, 2004; Pintor et al., 2008).
The high degree of cannibalism displayed by C. kelberi highlights the aggressive behavior exhibited by such species and this is reported for several locations (Fugi et al., 2008). Despite being an invasive species, C. kelberi had the second highest abundance and the second highest piscivorous fish consumption, unlike its congener C. piquiti. This last species did not consume any piscivorous fish in this study, did not present cannibalism but had the lowest abundance of all the analyzed species. When analyzing this case, piscivorous consumption can be one of the mechanisms responsible for the success of C. kelberi in establishing in the floodplain, while its congener, that did not consume fish, failed to establish in the floodplain.
The high abundance of the invasive C. kelberi is of concern due to the species ability to change the habitat where it is introduced. Pelicice & Agostinho (2009) observed that the introduction of this species in the Rosana Reservoir (Paranapanema River, a tributary of the upper Paraná River, Brazil) resulted in a 95% decline in native fish density and an 80% decline in richness in only 2 years. These authors also suggest that once the species is established, it is difficult to minimize the negative impacts in the receptor environment. More recently, C. kelberi had high capture in Capivara Reservoir (Brazil), which, as a consequence, increased the predation on native fishes (Orsi & Britton, 2014).
Invasive species face new ecological interactions in the invaded environment. Thus, the success of an invasion might be affected by the biotic resistance presented by competitors and predators (Bajer et al., 2012; Thompson et al., 2012). Santos et al. (2013) detected that juveniles S. brasiliensis can exert biotic resistance to invasive species in Neotropical ecosystems, highlighting the importance of the conservation of S. brasiliensis. Invasive species are also consumed by native piscivorous species. In the present study, the diet of the native S. brasiliensis was composed by piscivorous individuals (26%) and, among them, the invasive C. kelberi (13%). This fact demonstrates that this native is acting directly as a predator of the invasive species, corroborating the importance of such species in providing biotic resistance against invaders.
Most of the invasive species considered in this paper were introduced during dam construction or reservoir management. Therefore, besides the role of reservoirs facilitating invasions and the dispersion of aquatic species (Havel et al., 2005; Johnson et al., 2008), the increasing use of reservoirs by sport fish (stocking piscivorous fish; Agostinho et al., 2010) and aquaculture (caging in reservoirs; Pelicice et al., 2014) elevate the risk of biodiversity impacts. As an example, S. brasiliensis, a migratory species, has been highly affected by reservoir construction which, combined with intense human occupation and fishing, causes serious depletions in its abundance (Agostinho et al., 2007; Hahn et al., 2011; Petrere-Jr et al., 2002). Our results show that piscivorous fish predation occurs independent of prey species origins and even with high availability of possible preys and can actually act as a mechanism for the success in establishment and maintenance of invasive species in the new environment. Also, native species can promote biotic resistance, in certain degree, by predating the new arrived species.
References
Agostinho, A. A., E. E. Marques, C. S. Agostinho, D. A. de Almeida, R. J. Oliveira, J. Rodrigues & B. Melo, 2007. Fish ladder of Lajeado Dam: migrations on one-way routes? Neotropical Ichthyology 5: 121–130.
Agostinho, A. A., F. M. Pelicice & L. C. Gomes, 2008. Dams and the fish fauna of the Neotropical region: impacts and management related to diversity and fisheries. Brazilian Journal of Biology 68: 1119–1132.
Agostinho, A. A., F. M. Pelicice, L. C. Gomes & H. F. Júlio-Júnior, 2010. Reservoir fish stocking: when one plus one may be less than two. Natureza & Conservação 08: 103–111.
Anderson, M. J., 2004. PERMDISP: a FORTRAN computer program for permutational analysis of multivariate dispersions (for any two-factor ANOVA design) using permutation tests. Department of Statistics, University of Auckland, Auckland.
Anderson, M. J., 2006. Distance based tests for homogeneity of multivariate dispersions. Biometrics 62: 245–253.
Bajer, P. G., C. J. Chizinski, J. J. Silbernagel & P. W. Sorensen, 2012. Variation in native micro-predator abundance explains recruitment of a mobile invasive fish, the common carp in a naturally unstable environment. Biological Invasions 14: 1919–1929.
Baxter, C. V., K. D. Faush, M. Murakami & P. L. Chapman, 2004. Fish invasion restructures stream and forest food webs by interrupting reciprocal prey subsidies. Ecology 85: 2656–2663.
Diamond, J. & T. J. Case, 1986. Overview: introductions, extinctions, exterminations and invasions. In Diamond, J. & T. J. Case (eds), Community Ecology. Harper & Row, San Francisco: 65–79.
Dick, J. T. A., 1996. Post-invasion amphipod communities of Lough Neagh, Northern Ireland: influences of habitat selection and mutual predation. Journal of Animal Ecology 65: 756–767.
Fugi, R., K. D. G. Luz-Agostinho & A. A. Agostinho, 2008. Trophic interaction between an introduced (peacock bass) and a native (dogfish) piscivorous fish in a Neotropical impounded river. Hydrobiologia 607(143): 150.
Graça, W. J. & C. S. Pavanelli, 2007. Peixes da planície de inundação do alto rio Paraná e áreas adjacentes. Maringá, Eduem.
Griffen, B. D. & J. E. Byers, 2006. Partitioning mechanisms of predator interference in different habitats. Oecologia 146: 608–614.
Hahn, L., A. A. Agostinho, K. K. English, J. Carosfeld, L. F. Câmara & S. J. Cooke, 2011. Use of radiotelemetry to track threatened dorados Salminus brasiliensis in the upper Uruguay River, Brazil. Endangered Species Research 15: 103–114.
Hall, R. J., 2011. Eating the competition speeds up invasions. Biology Letters 7: 307–311.
Havel, J. E., C. E. Lee & M. J. Vander Zanden, 2005. Do reservoirs facilitate invasions into landscapes? BioScience 55: 518–525.
Hochberg, M. E., 1996. Consequences for host population levels of increasing natural enemy species richness in classical biological control. American Naturalist 147: 307–318.
Holway, D. A. & A. V. Suarez, 1999. Animal behavior: an essential component of invasion biology. Trends in Ecology and Evolution 14: 328–330.
Hurd, L. E. & R. M. Eisenberg, 1990. Arthropod community responses to manipulation of a bitrophic predator guild. Ecology 71: 2107–2114.
Hynes, H. B. N., 1950. The food of freshwater sticklebacks (Gasterosteus aculeatus and Pygosteus pungitius) with a review of methods used in studies of the food of fishes. Journal of Animal Ecology 19: 36–58.
Hyslop, E. J., 1980. Stomach contents analysis, a review of methods and their application. Journal of Fish Biology 17: 411–429.
Johnson, P. T. J., J. D. Olden & M. J. Vander Zanden, 2008. Dam invaders: impoundments facilitate biological invasions into freshwaters. Frontiers in Ecology and the Environment 6: 357–363.
Júlio-Júnior, H. F., C. Dei Tos, A. A. Agostinho & C. S. Pavanelli, 2009. A massive invasion of fish species after eliminating a natural barrier in the upper rio Paraná basin. Neotropical Ichthyology 7: 709–718.
Langeani, F., O. T. Oyakawa, O. A. Shibatta, C. S. Pavanelli & L. Casatti, 2007. Diversidade da ictiofauna do Alto Rio Paraná : composição atual e perspectivas futuras. Biota Neotropica 7: 181–197.
Legendre, P. & L. Legendre, 1998. Numerical Ecology. Elsevier Science, Amsterdam.
Losey, J. E. & R. F. Denno, 1998. Positive predator-predator interactions: enhanced predation rates and synergistic suppression of aphid populations. Ecology 79: 2143–2152.
Luiz, T. F., M. R. Velludo, A. C. Peret, J. L. Rodrigues & A. M. Peret, 2011. Diet, reproduction and population structure of the introduced Amazonian fish Cichla piquiti (Perciformes : Cichlidae) in the Cachoeira Dourada reservoir (Paranaíba River, central Brazil). International Journal of Tropical Biology 59: 727–741.
MacNeil, C. & J. Prenter, 2000. Differential microdistributions and interspecific interactions in coexisting native and introduced Gammarus spp. (Crustacea: Amphipoda). Journal of Zoology 251: 377–384.
Moyle, P. B. & T. Light, 1996. Biological invasions of fresh water: empirical rules and assembly theory. Biological Conservation 78: 149–161.
Okubo, A., P. K. Maini, M. H. Williamson & J. D. Murray, 1989. On the spatial spread of the grey squirrel in Britain. Proceedings of the Royal Society B: Biological Sciences 238: 113–125.
Oliveira, E. F., E. Goulart, L. Breda, C. V. Minte-Vera, L. Ricardo, D. S. Paiva & M. R. Vismara, 2010. Ecomorphological patterns of the fish assemblage in a tropical floodplain : effects of trophic, spatial and phylogenetic structures. Neotropical Ichthyology 8: 569–586.
Orsi, M. L. & J. R. Britton, 2014. Long-term changes in the fish assemblage of a neotropical hydroelectric reservoir. Journal of Fish Biology 44: 1–7.
Pazza, R. & H. F. Júlio-Jr, 2003. Occurrence of three sympatric cytotypes of Hoplias malabaricus (Pisces, Erythrinidae) in the upper Paraná river floodplain (Brazil). Cytologia 68: 159–163.
Pelicice, F. M. & A. A. Agostinho, 2009. Fish fauna destruction after the introduction of a non-native predator (Cichla kelberi) in a Neotropical reservoir. Biological Invavasions 11: 1789–1801.
Pelicice, F. M., J. R. S. Vitule, D. P. Lima Junior, M. L. Orsi & A. A. Agostinho, 2014. A serious new threat to Brazilian freshwater ecosystems: the naturalization of nonnative fish by decree. Conservation Letters 7: 55–60.
Petrere-Jr, M., A. A. Agostinho, E. Okada & H. F. Júlio-Junior, 2002. Review of the fisheries in the Brazilian portion of Paraná River. In Cowx, I. G. (ed.), Management and Ecology os Lake and Reservoir Fisheries. Wiley, New York: 123–143.
Pintor, L. M., A. Sih & M. L. Bauer, 2008. Differences in aggression, activity and boldness between native and introduced populations of an invasive crayfish. Oikos 117: 1629–1636.
Rehage, J. S. & A. Sih, 2004. Dispersal behavior, boldness and the link to invasiveness: A comparison of our Gambusia species. Biological Invasions 6: 379–391.
Rice, E. S. & J. Silverman, 2013. Propagule pressure and climate contribute to the displacement of Linepithema humile by Pachycondyla chinensis. PloS One 8: 1–11.
Santos, A. F. G. N., E. García-Berthou, C. Hayashi & L. N. Santos, 2013. When habitat complexity increases predation risk : experiments with invasive and neotropical native fishes. Marine and Freshwater Research 64: 752–760.
Schoener, T. W., 1970. Non-synchronous spatial overlap of lizards in patchy habitats. Ecology 51: 408–418.
Sharma, S., M. J. V. Zanden, J. J. Magnuson & J. Lyons, 2011. Comparing climate change and species invasions as drivers of coldwater fish population extirpations. PlosOne 6: 1–9.
Shinen, J. S., S. G. Morgan & A. L. Chan, 2009. Invasion resistance on rocky shores: direct and indirect effects of three native predators on an exotic and a native prey species. Marine Ecology Progress Series 378: 47–54.
Sih, A., G. Englund & D. Wooster, 1998. Emergent impacts of multiple predators on prey. Trends in Ecology & Evolution 13: 350–355.
Sih, A., D. I. Bolnick, B. Luttbeg, J. L. Orrock, S. D. Peacor, L. M. Pintor, E. Preisser, J. S. Rehage & J. R. Vonesh, 2010. Predator-prey naïveté, antipredator behavior, and the ecology of predator invasions. Oikos 119: 610–621.
Svenning, J. C., D. Gravel, R. D. Holt, F. M. Schurr, W. Thuiller, T. Münkemüller, K. H. Schiffers, S. Dullinger, T. C. Edwards, T. Hickler, S. I. Higgins, J. E. M. S. Nabel, J. Pagel & S. Normand, 2014. The influence of interspecific interactions on species range expansion rates. Ecography. doi:10.1111/j.1600-0587.2013.00574.x.
Thompson, K. A., J. E. Hill & L. G. Nico, 2012. Eastern mosquitofish resists invasion by nonindigenous poeciliids through agonistic behaviors. Biological Invasions 14: 1515–1529.
Zaret, T. M. & R. T. Paine, 1973. Species introduction in a tropical lake. Science 182: 449–455.
Acknowledgments
We thank João Carlos Barbosa da Silva for contributions and support in the manuscript, Luiz Fernando Caserta Tencatt for the fish identification and CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) for financial support. AAA and LCG are researchers from CNPq (Bolsa Produtividade).
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: Sidinei M. Thomaz, Katya E. Kovalenko, John E. Havel & Lee B. Kats / Aquatic Invasive Species
Rights and permissions
About this article

Cite this article
Pereira, L.S., Agostinho, A.A. & Gomes, L.C. Eating the competitor: a mechanism of invasion. Hydrobiologia 746, 223–231 (2015). https://doi.org/10.1007/s10750-014-2031-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-014-2031-1