
Abstract
Dormant propagule pools (egg banks) of zooplankton populations that accumulate in sediments provide biological archives of past conditions and enable the investigation of evolutionary changes in populations over relatively long periods of time (many decades). This study examined the egg bank of a Daphnia pulicaria population in a lake that has been stocked annually with rainbow trout (a zooplanktivore) since 1961. Resting eggs from sediments from the 1920s to 2001 (determined by 210Pb dating) were hatched and established as isofemale clonal lines. The phototactic behavior (a proxy for vertical migration behavior) of clones was assessed in the presence and absence of fish kairomones to evaluate the hypothesis that the heavy and consistent level of predation imposed by the stocked trout would have selected for clones that are more negatively phototactic. In addition, exposure to kairomones was expected to induce stronger negative phototaxis for all clones relative to control conditions. The behavior of clones derived from resting eggs from the trout era (after 1961) was significantly more negatively phototactic than it was for pre-trout era clones. Kairomone exposure induced a more negative phototactic response in clones from both eras, but the response was much greater for the pre-trout era clones. These results suggest that the consistently high level of predation by trout over several decades has selected for a conservative (less plastic) vertical migration strategy in which Daphnia maintain a deep daytime distribution with or without the presence of chemical cues from fish predators.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Resting embryos of zooplankton deposited in sediments (egg banks) provide a record of a population’s evolutionary history and are important reservoirs of biological diversity that may affect the genetic structure and evolutionary dynamics of a population (Hairston et al., 1999).
Experiments in which resting embryos are hatched or “resurrected” (Kerfoot et al., 1999) from sediments of different age provide the opportunity to examine evolutionary change in populations over longer time periods than is usually feasible (i.e., decades vs. years). For parthenogenetically reproducing organisms such as Daphnia, hatchlings from resting embryos may be used to establish isofemale clonal cultures. Individuals from these clonal cultures can then be used as replicates (of the same genotype) in experiments to evaluate whether a population has evolved over time. “Resurrection ecology” studies of this type have documented increased tolerance of Daphnia to toxic cyanobacteria in a lake that experienced eutrophication (Hairston et al., 1999), the evolution of increased phenotypic plasticity in phototactic behavior for Daphnia under changing levels of fish predation (Cousyn et al., 2001), changes in helmet and spine morphology of D. retrocurva in response to differing abundances of invertebrate and vertebrate predators (Kerfoot & Weider, 2004), “Red Queen” dynamics between Daphnia and the endoparasite Pasteuria ramosa (Decaestecker et al., 2007), an increased innate immune response in Daphnia during a period of intensified parasitism (Pauwels et al., 2010), and decreased interspecific competition over time among sympatric Daphnia species (Steiner et al., 2007).
This study uses resurrection ecology techniques to examine whether the annual stocking of the zooplanktivorous rainbow trout (Oncorhynchus mykiss Walbaum) over several decades (since 1961) has caused evolutionary change in the phototactic behavior of the Daphnia pulicaria Forbes population in Long Lake (northwestern Minnesota, USA). Previous research on this system (from 1996 to 1999) has shown that rainbow trout predation significantly affects the growth dynamics (Hembre & Megard, 2005) and genetic composition (Hembre & Megard, 2006) of the resident D. pulicaria population.
Rainbow trout preferentially prey on D. pulicaria in Long Lake and inhibit the growth of the population to the extent that spring “clear-water phases” (Lampert et al., 1986) are prevented when trout are abundant over winter (Hembre & Megard, 2005). Genetic (allozyme) analyses have indicated that the Daphnia population becomes genetically differentiated with respect to depth during the period of summer stratification with certain clones displaying affinities for deeper (hypolimnetic) or shallower (metalimnetic) depths during the daytime (Hembre & Megard, 2006). In addition, the genetic composition of Daphnia found in the stomachs of trout was similar to that of Daphnia from net samples collected from metalimnetic depths, but significantly different from Daphnia collected from the hypolimnion. This indicates that trout feed selectively on clones in the relatively shallow, well oxygenated (>5 mg l−1) portion of the metalimnion, while clones occupying deep-water refuges where it is darker and more hypoxic are less likely to be consumed by the trout (Hembre & Megard, 2006). Previous research (Hembre & Megard, 2006) also indicates that the genetic composition of the population is structured by a trade-off between a greater risk of mortality from trout predation in shallower water versus reproductive benefits associated with a shallower depth distribution (i.e., faster development at warmer temperatures and higher fecundity with more abundant food resources; Loose & Dawidowicz, 1994). During a 4-year monitoring period (1996–1999), metalimnetic clones predominated when trout densities were relatively low, and hypolimnetic clones comprised the majority of the population when trout were more abundant. The dramatic shifts in the dominant clones over a few years in the Long Lake D. pulicaria population illustrate that genetic change in clonally reproducing populations has the potential to be rapid. However, because of the “boom and bust” nature of zooplankton populations (e.g., Lampert et al., 1986), lasting changes in gene frequencies in cyclically parthenogenetic populations will depend upon the genetic composition of sexually derived individuals that are deposited in and emerge from diapausing embryos in the egg bank.
For this study, D. pulicaria ephippia (capsules containing sexually produced resting embryos) were obtained from core samples of Long Lake sediment dating back to the 1920s. Hatchlings from the ephippia were used to establish isofemale clonal cultures, and individuals from the cultures were used in laboratory assays to assess the phototactic behavior (De Meester, 1991) of the clones in the presence and absence of fish kairomones. We hypothesized that increased levels of predation since the initiation of the trout stocking program (in 1961) would have selected for more negatively phototactic clones, and that the presence of fish kairomones would amplify this response.
Materials and methods
Study site and fisheries management
Long Lake is a 66.5 ha meso- to oligotrophic dimictic lake in northwestern Minnesota, USA. (latitude 47°17′N, longitude 95°17′W). The lake has a single basin and is steep-sided and relatively deep (mean depth 13 m, maximum depth 24 m). The lake is spring-fed, has no inflowing streams, and has a single outlet at its southeast end. Therefore, the resident D. pulicaria population is not likely to experience significant immigration from outside sources. Long Lake has been stocked annually with rainbow trout by the Minnesota Department of Natural Resources (MDNR) since 1961, and is one of more than 100 lakes in Minnesota managed as “two-story” fisheries in which stream trout (e.g., rainbow trout, or brown trout, Salmo trutta Linnaeus) are stocked to provide a cold-water sport fishery for anglers. Lakes managed as two-story fisheries by the MDNR have water quality that enables sufficient habitat for the trout, which require cold and well-oxygenated water (<21°C and >5 mg l−1 O2; Wang et al., 1996). Rainbow trout are pelagic planktivores that avoid warm surface waters during the summer. Instead, they inhabit deeper waters that are sufficiently cold and preferentially prey on D. pulicaria in Long Lake (Hembre & Megard, 2005). While the lake’s fish community also includes several native warm-water zooplanktivorous fishes such as yellow perch (Perca flavescens Mitchill), bluegill (Lepomis macrochirus Rafinesque) and pumpkinseed (L. gibbosus Linnaeus) sunfish, and minnow species (Cyprinidae) (2006 MDNR fish survey, unpublished data), the annual stocking of rainbow trout increases predation on Daphnia in the pelagic zone above natural levels.
Core collection and sediment processing
Four sediment cores (21–29 cm in length) were collected from the deepest part of the lake (water depth 24 m) on 24 July 2001 using a gravity corer (internal diameter 7 cm). Polycarbonate tubes containing the cores were wrapped in aluminum foil and immediately transported to the laboratory where sediments were extruded and sliced at 1 cm intervals. Sediments were placed in opaque containers and stored in the dark at 4°C for future analyses. Samples from three of the cores (21–24 cm in length) were used as sources of ephippia for enumeration and use in hatching experiments, and sediment samples from the longest core (29 cm) were used to determine the age of the sediment (via 210Pb dating).
Sediments from every other 1-cm interval to a depth of 29 cm (n = 15) were analyzed for 210Pb. 210Po distillation and alpha spectrometry methods (Eakins & Morrison, 1978) were used to determine 210Pb activity. Dates and sedimentation rates were calculated using the constant rate of supply dating model (Appleby & Oldfield, 1983). Dry density, water content, carbonate content, and organic content were determined for each sediment sample using loss-on-ignition methods (Dean, 1974).
Ephippia were isolated from samples from the other three cores during the spring and summer of 2002 according to the methods of Cáceres (1998). Sediment samples were first rinsed over a fine (80-μm mesh) sieve to remove as much of the fine sediment as possible. The remaining sediment was placed in a 15-ml centrifuge tubes containing a 30% sucrose solution and centrifuged at 1,085 rpm for 1.5 min. This treatment caused most of the ephippia to become suspended in the supernatant. The supernatant was then poured onto an 80-μm mesh sieve and the ephippia were rinsed with dechlorinated tap water into opaque vials for later examination. In addition, pellets at the bottom of the centrifuge tubes were examined under a dissecting microscope and any remaining ephippia were removed and placed in the vials. Ephippia were inspected microscopically to determine the specific identity of the ephippia. The dorsal ridge of the D. pulicaria ephippium is lined with tiny hairs, while ephippia of D. mendotae (the other commonly observed daphnid in Long Lake) are smaller with a smooth dorsal ridge (Hebert, 1995). Open ephippial cases and those containing degenerated eggs were discarded, and ephippia containing two resting eggs that appeared to be in good condition were stored in the dark at 4°C for later use in hatching experiments.
Hatching and culturing methods
Ephippia were hatched in a flow-through tube system. The polycarbonate tubes (91-cm long, 15-mm-internal cross section) are open at the top and have 150 μm Nitex © mesh at the bottom. A peristaltic pump continuously delivers media (dechlorinated water in this case) into the tubes at the top, and the water exits the tubes through the mesh on the bottom. The system was illuminated from above with a light bank consisting of two 40-W cool-white fluorescent tubes positioned 0.5 m above the tubes and the temperature was maintained at 21°C. Twenty-five ephippia isolated from a given 1 cm sediment depth increment were placed on the mesh screen at the base of each tube, and four replicate tubes were used for each sediment increment from the top of the cores to a depth of 15 cm. Each ephippium contained two resting eggs, so the maximum number of hatchlings for each replicate tube was 50, and the maximum for each sediment depth was 200. Hatching was monitored daily over a 10-day period. Previous studies (e.g., Schwartz & Hebert, 1987; De Meester & De Jager, 1993) have shown that the vast majority of hatching occurs within 10 days of exposure to hatching cues. Each day, hatchlings were removed with a long glass pipette and placed in separate 50-ml flasks containing dechlorinated tap water to initiate isofemale clonal cultures. As initial cultures developed, additional cultures of each isofemale lineage were initiated in separate 0.5-l flasks and maintained under standardized conditions in a Percival © (model 1-30 B) growth chamber at 16°C with a 14:10 light:dark cycle and animals were fed with Scenedesmus acutus. Half of the culture medium was removed and refreshed with dechlorinated tap water every 2 days, and Scenedesmus concentrations were maintained at 5 × 105 cells ml−1. Daphnia concentrations in the cultures were maintained at moderate levels (~20 adults l−1), and any individuals producing ephippia were removed from the cultures to maintain pure clonal cultures.
Phototaxis assays
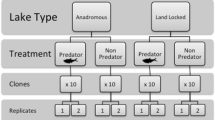
The phototactic behavior of clones was quantified using a laboratory assay. This assay is described in detail in De Meester (1991) and has subsequently been used in many published studies (e.g., De Meester, 1996; Cousyn et al., 2001; Decaestecker et al., 2002). The experimental setup for the assay consisted of a transparent cylinder (25 cm in height, 5 cm internal diameter) that was externally divided into three compartments: lower (L, 0–3 cm), middle (M, 3–12 cm), and upper (U, 12–25 cm). The column was placed in a darkened box and illuminated from above with a 150-W halogen fiber bulb. To minimize variability due to any endogenous rhythm, experiments were performed between 12 and 6 pm. The first phototaxis assays were performed 7 weeks after the hatching experiments were completed and after the clonal lineages had been cultured under standardized conditions in the growth chamber for multiple generations (to exclude maternal effects). Ten gravid adult females (1.6–2.0 mm in length—from top of head to base of tail spine) were randomly selected from different clonal cultures for each trial (to avoid common environment effects) and deposited into the column filled with treatment media (either dechlorinated tap water or kairomone water). Kairomone water was obtained from an 8-l tank containing three 6-cm-long goldfish (Carassius auratus Linnaeus). Goldfish were fed Tetrafin Goldfish Flakes (produced by Tetra ©). Media used in the experiments was filtered with a glass fiber filter (1-μm pore size) before it was added to the column. After a 5-min-dark acclimation period, the light source was turned on and the locations of the animals were recorded each minute for 10 min. The phototactic behavior of the animals was calculated using the phototactic index I = (U − L)/(U + M + L). U, M, and L are the number of animals observed in the upper, middle, and lower compartments of the column, respectively. Index values can range from −1 (all animals in the lower compartment) to +1 (all animals in the upper compartment). Values were averaged for each 10-min trial, and three replicate trials of the assay were performed (using ten different individuals from different cultures) for each isofemale clone. Ten isofemale clones from the pre-trout era and 26 clones from the trout era were assayed. Relatively low levels of hatching success from ephippia from deeper sediments prevented us from being able to assay as many pre-trout era clones as trout era clones.
Data analysis
Phototactic index data from 216 trials (36 clones × 3 replicate trials/clone × 2 treatments) were found to be normally distributed and were subsequently analyzed with linear mixed effect (lme) models using the statistical package R (2011). A lme model analysis approach was employed for several reasons: (1) there were random and fixed factors in our experiment, (2) experimental units were nested in a hierarchy (clones within eras), and (3) because the design was unbalanced (different numbers of clones from the two eras). Clone was treated as a random factor nested within era (pre-trout vs. trout era). The interaction of clone and treatment was also treated as a random factor. Era, treatment (control vs. kairomone media), and the interaction between era and treatment were treated as fixed factors. Parameters were estimated using maximum likelihood.
Results
Core dating
210Pb analysis of the long (29 cm) core shows a monotonic decrease in 210Pb activity (except for a discontinuity at 8–9 cm) from 1.19 Bq g−1 in surface sediments to stable background (supported) values of 0.028 Bq g−1 below 21 cm (Fig. 1, left panel). Ages calculated by the constant rate of supply model (Appleby & Oldfield, 1983) give a date of 1832 (±33 year) for the 20- to 21-cm interval, and 1962.7 (±2.7 year) at the 8- to 9-cm interval. These results indicate that sediments shallower than 9 cm were deposited after the initiation of the rainbow trout stocking program, and those below 9 cm were deposited before the trout era (Fig. 1, right panel). The decrease in 210Pb activity in the 8–9 cm increment corresponds to unusually high sedimentation rates (Fig. 2) and the subsequent dilution of 210Pb at that depth. Loss-on-ignition data indicate that the increased sedimentation rate at this interval was caused by increased deposition of clastic inorganic material (~2-fold increase over baseline) at this depth in the core and suggest that an erosional event occurred in the lake’s watershed during that period. Activities related to the establishment of a county park (i.e., tree removal, creation of a swimming beach, and construction of a parking lot and concrete boat ramp) at the northwest end of Long Lake in 1965 are the likely cause for the increased deposition of inorganic sediments during that time period.

Total 210Pb activity (left) and sediment date (right) versus sediment depth from a core collected from Long Lake in 2001 (error bars ±1 SD). Dating indicates that sediments below 9 cm were deposited before the initiation of the rainbow trout stocking program (before 1961). The dashed line on the left-hand side panel indicates the level of supported 210Pb (estimated as the mean 210Pb activity of the four samples from the bottom of the core). The dashed line on the right-hand side panel marks the year in which rainbow trout were first stocked in Long Lake by the Minnesota Department of Natural Resources

Sediment accumulation rate versus 210Pb date for the core (error bars ±1 SD). Note: The spike in the sedimentation rate around the 1960 210Pb date (8–9 cm depth in the core) is likely due to erosion from the watershed during the construction of a county park in the 1960s
Ephippia densities and hatching rates
Densities of D. pulicaria ephippia in the sediment cores were highest between 5 and 8 cm and lowest in sediments below 13 cm (Fig. 3, left panel). The average hatching success of ephippia (Fig. 3, right panel) obtained from sediments that were more recently deposited (shallower than 5 cm) was considerably higher (12.5–34.5%), than for ephippia extracted from sediments deeper than 5 cm (all ≤5%).

Mean density of D. pulicaria ephippia (left), and mean hatching success (right) of incubated eggs with respect to sediment depth (error bars 1 SE). The dashed line indicates the sediment depth that corresponds to the year (1961) that the rainbow trout stocking program was initiated in Long Lake
Phototaxis assay results
Results of a lme model analysis indicate that a significant amount of the observed variation in the phototactic index data was accounted for by the random factors of clone [likelihood ratio (LR) test statistic 58.0, df 1, P < 0.0001), and of the interaction between clone and treatment (LR 96.0, df 1, P < 0.0001). Accounting for these random factors, all of the fixed factors in the lme analysis were also significant (Table 1). The phototactic behavior of clones from the trout era differed significantly from that of the pre-trout era clones (P = 0.005), and the phototactic response of Daphnia exposed to fish kairomones was significantly different than it was for the control treatment (P < 0.001). While the mean phototactic response of clones from both eras was more negative when exposed to fish kairomones, there was significantly greater plasticity in this response for clones from the pre-trout era. The decreased plasticity observed in the clones from the trout era was a result of a more negative phototactic response in the absence of fish kairomones relative to that response among the pre-trout era clones (Fig. 5). This difference in plasticity between clones from the different eras accounts for the significant interaction between era and treatment (P = 0.002).
Exposure to fish kairomones was less likely to induce a more negatively phototactic response in clones from the trout era versus those from the pre-trout era (Fig. 4). All 10 of the pre-trout era clones assayed were more negatively phototactic in the presence of kairomones, while a smaller proportion (18 of 26) of the clones from the trout era showed this response. For clones from the trout era, 13 of the 17 (76%) clones from ephippia derived from sediments deposited in the 1970s and 1980s were more negatively phototactic in the presence of fish kairomone, while only five of the nine (56%) clones from ephippia from sediments deposited since 1990 had this response.

Mean phototactic behavior (±1 SD) of isofemale clones in the presence (filled symbols) and absence (open symbols) of fish kairomone. Clones are plotted from left to right by increasing sediment depth (age). Approximate ages (accounting for error estimates for the 210Pb dates—see Fig. 1) of the sediments in these groups: 0–2 cm (1990–2001), 3–5 cm (1980s), 6–8 cm (1970s), and 9–14 cm (1920–1960). The solid vertical line indicates the sediment depth that corresponds to the date (1961) that the rainbow trout stocking program began in Long Lake. Dashed vertical lines delineate the sediment depth groupings within the trout era (after 1961)
Four of the 36 clones assayed in the experiment were derived from ephippia from sediments near the transition between the pre-trout era and the trout era (i.e., sediments with 210Pb dates within 2 SD of 1961). One clone was from the 7- to 8-cm core depth (assigned to the trout era in the original analysis) and three were from the 9- to 10-cm core depth (assigned to the pre-trout era in the original analysis). To evaluate the effects of these four clones on the statistical results, lme model analyses were performed with different combinations of assignments for the clones and with their omission from the data set. Results of these analyses did not differ appreciably from each other or from the original analysis. All main effects and interaction effects were still highly significant (P < 0.01).
Discussion
The phototactic behavior of the clones assayed differed significantly between eras and between treatments (Figs. 4, 5). There was also a marked difference in the plasticity of the phototactic response of clones from the different eras (Fig. 5) when exposed to the different treatments (control vs. kairomone media). In the absence of fish kairomone, the phototactic behavior of trout era clones was clearly more negatively phototactic than it was for pre-trout era clones (Fig. 5). This finding supports our hypothesis that predation by the annually stocked rainbow trout would have selected for Daphnia that are more negatively phototactic than those that inhabited the lake during the pre-trout era (before 1961). In addition to the significant change in the “baseline” phototactic behavior of the population, the other striking difference between the clones from the two eras is that Daphnia from pre-trout era were much more plastic in their phototactic response to the presence of fish kairomones than were the trout era clones. The average phototactic behavior of the trout era clones when exposed to kairomones was only slightly more negative than their baseline behavior, while pre-trout era clones displayed a more consistent (Fig. 4—all pre-trout era clones assayed were more negatively phototactic in the kairomone treatment) and much larger (Fig. 5) response to the kairomone treatment.

Mean phototactic behavior of clones from the trout and pre-trout eras in Long Lake in the presence (filled symbols) and absence (open symbols) of fish kairomone. Error bars ±SE
The change in the Long Lake food web due to the annual stocking of rainbow trout since 1961 is a plausible cause for the change in the plasticity of the phototactic response of clones between eras (Table 1; era × treatment interaction in the lme model, P = 0.002). Before 1961, only naturally occurring populations of zooplanktivores were present in the lake. Reproductive success for fish populations is known to vary considerably among years (Rudstam et al., 1993). So, during the pre-trout era in Long Lake when levels of zooplanktivorous fish were controlled by natural reproduction, Daphnia clones able to adjust their vertical migration behavior according to the perceived risk of predation (via detection of kairomones) would likely have had selective advantage over those with a more fixed behavior. Our finding that the phototactic response to fish kairomones by pre-trout era clones was more plastic than it was for the trout era clones implies that Daphnia from the pre-trout era would have migrated to deeper daytime depths when the perceived risk of predation (via detection of kairomones) was high, and would have likely maintained a shallower daytime distribution when the perceived risk of predation was low to avoid incurring the costs (slower growth and reproduction) of inhabiting the colder deeper water (Loose & Dawidowicz, 1994). In contrast, clones from the trout era were similarly negatively phototactic both in the presence and absence of fish kairomones (Fig. 5) and appear to have evolved a more conservative vertical migration strategy. This suggests that phenotypic plasticity in phototactic behavior has been selected against over the many generations during which Daphnia in Long Lake have been subjected to consistently high levels of zooplanktivory by rainbow trout. De Meester (1993) observed a similar lack of plasticity in phototactic behavior for clones isolated from environments with high levels of zooplanktivorous fish, noting that the baseline negative phototactic behavior of clones was not dramatically altered by a fish cue. While either adaptive phenotypic plasticity or genetic adaptation can enable a population to persist when faced with a rapidly changing environment (Latta et al., 2007), our results suggest that the plastic response in phototactic behavior to increased predation was not sufficient to enable the survival of Daphnia with that strategy, and that genetic change in the standard phototactic response occurred.
Interestingly, our finding that the extent of phenotypic plasticity in phototactic behavior became less when the Daphnia population in Long Lake was subjected to higher levels of predation (i.e., since 1961) differs from the results of a similar study (Cousyn et al., 2001; and reviewed in Hairston & De Meester, 2008) on a Daphnia population from a small manmade fish rearing pond (Oud Heverlee Pond) in Belgium. In that study, Daphnia clones were “resurrected” from pond sediments that had been deposited during two different time periods when levels of fish stocking were low (1970–1972 and 1982–present) and an intervening period (1973–1982) when zooplanktivorous fish were stocked at a high level. For clones derived from ephippia during the two time periods when predation pressure was low, exposure to fish kairomones did not substantially alter phototactic behavior. However, clones derived from sediments from the 9-year period (1973–1982) when fish stocking levels were high displayed considerably more plasticity in their phototactic behavior and typically became more negatively phototactic in the presence of fish kairomones. The opposite effect of increased fish predation pressure on the plasticity of phototactic behavior in the Daphnia populations of Long Lake and Oud Heverlee Pond is likely a consequence of different historical circumstances in the two water bodies. Long Lake is a natural lake that was formed by glacial processes ~12 thousand years ago, while Oud Heverlee Pond is manmade and was constructed in 1970. Therefore, the Daphnia population in Long Lake would very likely have experienced considerable variation in predation intensity over the millennia before the trout stocking program began and possessing flexibility in phototactic behavior would likely have been adaptive. Conversely, Daphnia present in Oud Heverlee Pond in the first few years of its creation were not subjected to zooplanktivorous predators, so responsiveness to fish kairomones would not have conferred a selective advantage. It was only once zooplanktivorous fish were stocked in Oud Heverlee Pond that clones in that Daphnia population evolved to become negatively phototactic in response to fish kairomones (Cousyn et al., 2001).
The evolution of the conservative vertical migration strategy among most clones in the contemporary D. pulicaria population in Long Lake (Fig. 4) means that individuals now are likely to spend more of their time in the deep water relative to populations from the pre-trout era that were comprised of clones with more plasticity in their phototactic behavior. This behavioral shift in which individuals experience colder temperatures in deep water could cause individuals to have longer development times and population growth rates to be slower. Other studies have documented that the physical or chemical presence of a predator (but not actual consumption of prey) can induce changes in prey behavior (Schmitz et al., 1997) or development (Peckarsky et al., 2002; Pangle & Peacor, 2006) that alters the growth dynamics of prey populations and the structure of food webs. The evolutionary changes in the phototactic behavior of Daphnia in Long Lake may be an example of a nonconsumptive effect (Peckarsky et al., 2008) by a predator (rainbow trout) that could have altered the population dynamics of its prey.
By assaying Daphnia clones “resurrected” from resting eggs for an ecologically relevant trait (phototactic behavior), we have shown that the consistently high level of predation by rainbow trout on Daphnia in relatively shallow (metalimnetic) water (Hembre & Megard, 2006) has caused the Daphnia population to evolve a more conservative diel vertical migration strategy in which the standard phototactic response of clones is now significantly more negative and less plastic (Fig. 5) than it was before the advent of the trout stocking program (in 1961). These results imply that selection favoring deep-water clones must have been strong and consistent enough to enable this evolutionary change despite the emergence of individuals over the decades from pre-trout era sediments that would have promoted genetic constancy (Ellner & Hairston, 1994; Hairston et al., 1996). It is difficult to predict if the Daphnia population in Long Lake would rapidly return to an assemblage of clones with highly plastic phototactic behavior if trout were no longer stocked to Long Lake and levels of zooplanktivores were again regulated by natural processes. While resting eggs produced in the pre-trout era (in sediments deeper than 9 cm) are probably permanently buried (Cáceres & Hairston, 1998), some variation among clones derived from ephippia from recent sediments remains (Fig. 4). So recruitment and proliferation of phenotypically plastic clones could enable the population to revert to its pre-trout era behavior. An intriguing direction for future research in resurrection ecology would be to examine the rate of reversibility in evolutionary change for Daphnia populations in lakes of different trophic state. More productive lakes with higher sedimentation rates would more quickly bury ephippia and make immigration from the more distant past unlikely. Conversely, for oligotrophic lakes with lower sedimentation rates, surface sediments would be likely to contain a pool of genotypes that spans a greater time horizon. Emergence from those older ephippia should influence contemporary populations to a greater degree and allow evolutionary trajectories to be more rapidly reversed.
References
Appleby, P. G. & F. Oldfield, 1983. The assessment of 210Pb data from sites with varying sediment accumulation rates. Hydrobiologia 103: 29–35.
Cáceres, C. E., 1998. Interspecific variation in the abundance, production, and emergence of Daphnia diapausing eggs. Ecology 79: 1699–1710.
Cáceres, C. E. & N. G. Hairston Jr., 1998. Benthic-pelagic coupling in planktonic crustaceans: the role of the benthos. Ergebnisse der Limnologie 52: 163–174.
Cousyn, C., L. De Meester, J. K. Colbourne, L. Brendonck, D. Verschuren & F. Volckaert, 2001. Rapid, local adaptation of zooplankton behavior to changes in predation pressure in the absence of neutral genetic changes. Proceedings of the National Academy of Sciences of the United States of America 98: 6256–6260.
De Meester, L., 1991. An analysis of the phototactic behaviour of Daphnia magna clones and their sexual descendants. Hydrobiologia 225: 217–227.
De Meester, L., 1993. Genotype, fish-mediated chemical, and phototactic behavior in Daphnia magna. Ecology 74: 1467–1474.
De Meester, L., 1996. Evolutionary potential and local genetic differentiation in a phenotypically plastic trait of a cyclical parthenogen, Daphnia magna. Evolution 50: 1293–1298.
De Meester, L. & H. De Jager, 1993. Hatching of Daphnia sexual eggs. I. Intraspecific differences in the hatching responses of D. magna eggs. Freshwater Biology 30: 219–226.
Dean, W. E. Jr., 1974. Determination of carbonate and organic matter in calcareous sediments and sedimentary rocks by loss on ignition: comparison with other methods. Journal of Sedimentary Petrology 44: 242–248.
Decaestecker, E., L. De Meester & D. Ebert, 2002. In deep trouble: habitat selection constrained by multiples enemies in zooplankton. Proceedings of the National Academy of Sciences of the United States of America 99: 5481–5485.
Decaestecker, E., S. Gaba, J. A. M. Raeymaekers, R. Stoks, L. V. Kerckhoven, D. Ebert & L. De Meester, 2007. Host–parasite ‘Red Queen’ dynamics archived in pond sediment. Nature 450: 870–873.
Eakins, J. D. & R. T. Morrison, 1978. A new procedure for the determination of lead-210 in lake and marine sediments. International Journal of Applied Radiation and Isotopes 29: 531–536.
Ellner, S. P. & N. G. Hairston Jr., 1994. Role of overlapping generations in maintaining genetic variation in a fluctuating environment. American Naturalist 143: 403–417.
Hairston, N. G. Jr. & L. De Meester, 2008. Daphnia paleogenetics and environmental change: deconstructing the evolution of plasticity. International Review of Hydrobiology 93: 578–592.
Hairston, N. G. Jr., S. Ellner & C. M. Kearns, 1996. Overlapping generations: the storage effect and the maintenance of biotic diversity. In Rhodes, O. E. J., R. K. Chesser & M. H. Smith (eds), Population Dynamics in Ecological Space and Time. University of Chicago Press, Chicago: 109–145.
Hairston, N. G. Jr., W. Lampert, C. E. Cáceres, C. L. Holtmeier, L. J. Weider, U. Gaedke, J. M. Fischer, J. A. Fox & D. M. Post, 1999. Rapid evolution revealed by dormant eggs. Nature 401: 446.
Hebert, P. D. N., 1995. The Daphnia of North America: an illustrated fauna (computer program). AuthorWare.
Hembre, L. K. & R. O. Megard, 2005. Timing of predation by rainbow trout controls Daphnia demography and the trophic status of a Minnesota lake. Freshwater Biology 50: 1064–1080.
Hembre, L. K. & R. O. Megard, 2006. Direct and indirect effects of predation on the genetic structure of a Daphnia population. Journal of Plankton Research 28: 1129–1141.
Kerfoot, W. C. & L. J. Weider, 2004. Experimental paleoecology (resurrection ecology): chasing Van Valen’s Red Queen hypothesis. Limnology and Oceanography 49: 1300–1316.
Kerfoot, W. C., J. A. Robbins & L. J. Weider, 1999. A new approach to historical reconstruction: combining descriptive and experimental paleolimnology. Limnology and Oceanography 44: 1232–1247.
Lampert, W., W. Fleckner, H. Rai & B. E. Taylor, 1986. Phytoplankton control by grazing zooplankton: a study on the spring clear-water phase. Limnology and Oceanography 31: 478–490.
Latta, L. C., J. W. Bakelar, R. A. Knapp & M. E. Pfrender, 2007. Rapid evolution in response to introduced predators II: the contribution of adaptive plasticity. BMC Evolutionary Biology 7: 21.
Loose, C. J. & P. Dawidowicz, 1994. Trade-offs in diel vertical migration by zooplankton: the costs of predator avoidance. Ecology 75: 2255–2263.
Pangle, K. L. & S. D. Peacor, 2006. Non-lethal effect of the invasive predator Bythotrephes longimanus on Daphnia. Freshwater Biology 51: 1070–1078.
Pauwels, K., L. De Meester, S. Put, E. Decaestecker & R. Stoks, 2010. Rapid evolution of phenoloxidase expression, a component of innate immune function, in a natural population of Daphnia magna. Limnology and Oceanography 55: 1408–1415.
Peckarsky, B. L., A. R. Mcintosh, B. W. Taylor & J. Dahl, 2002. Predator chemicals induce changes in mayfly life history traits: a whole-stream manipulation. Ecology 83: 612–618.
Peckarsky, B. L., P. A. Abrams, D. I. Bolnick, L. M. Dill, J. H. Grabowski, B. Luttbeg, J. L. Orrock, S. D. Peacor, E. L. Preisser, O. J. Schmitz & G. C. Trussell, 2008. Revisiting the classics: considering nonconsumptive effects in textbook examples of predator–prey interactions. Ecology 89: 2416–2425.
R Development Core Team, 2011. R: a language and environment for statistical computing. R-Foundation for Statistical Computing. Vienna, Austria. http://www.R-project.org.
Rudstam, L. G., R. C. Lathrop & S. R. Carpenter, 1993. The rise and fall of a dominant planktivore: direct and indirect effects on zooplankton. Ecology 74: 303–319.
Schmitz, O. J., A. P. Beckerman & K. M. O’Brien, 1997. Behaviorally mediated trophic cascades: effects of predation risk on food web interactions. Ecology 78: 1388–1399.
Schwartz, S. S. & P. D. N. Hebert, 1987. Methods for the activation of resting eggs of Daphnia. Freshwater Biology 17: 373–379.
Steiner, C. F., C. E. Caceres & S. D. Smith, 2007. Resurrecting the ghost of competition past with dormant zooplankton eggs. American Naturalist 169: 416–422.
Wang, L., K. Zimmer, P. Dietrich & S. Williams, 1996. The two-story rainbow trout fishery and its effect on the zooplankton community in a Minnesota lake. Journal of Freshwater Biology 11: 67–80.
Acknowledgments
Financial support for this research was provided by the Hamline University Lund Fund and the National Science Foundation (DIGG Grant #0105009). We are very grateful to Dan Engstrom (Science Museum of Minnesota St. Croix Watershed Research Station) for performing the 210Pb dating of the sediment core, to Brian Johnson and Doug Schnurrenburger for their assistance in coring Long Lake, to Emily Walsh and Rebecca Forman for help with the hatching experiments, to Frank Shaw for advice on statistical analyses, and to Robert Sterner for providing access to his laboratory at the University of Minnesota. Lastly, we thank Robert Megard for his consultation at various stages of this research.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Jasmine Saros
Rights and permissions
About this article
Cite this article
Hembre, L.K., Peterson, L.A. Evolution of predator avoidance in a Daphnia population: evidence from the egg bank. Hydrobiologia 700, 245–255 (2013). https://doi.org/10.1007/s10750-012-1234-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-012-1234-6