
Abstract
To investigate the effects of nitrate enrichment, phosphate enrichment, and light availability on benthic algae, nutrient-diffusing clay flowerpots were colonized with algae at two sites in a Hawaiian stream during spring and autumn 2002 using a randomized factorial design. The algal assemblage that developed under the experimental conditions was investigated by determining biomass (ash-free dry mass and chlorophyll a concentrations) and composition of the diatom assemblage. In situ pulse amplitude-modulated fluorometry was also used to model photosynthetic rate of the algal assemblage. Algal biomass and maximum photosynthetic rate were significantly higher at the unshaded site than at the shaded site. These parameters were higher at the unshaded site with either nitrate, or to a lesser degree, nitrate plus phosphate enrichment. Analysis of similarity of diatom assemblages showed significant differences between shaded and unshaded sites, as well as between spring and autumn experiments, but not between nutrient treatments. However, several individual species of diatoms responded significantly to nitrate enrichment. These results demonstrate that light availability (shaded vs. unshaded) is the primary limiting factor to algal growth in this stream, with nitrogen as a secondary limiting factor.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Because of their short generation times, relative lack of mobility, and rapid growth, stream algae are highly useful indicators of environmental conditions, such as water quality (Lowe & Pan, 1996). Different taxa have different nutrient requirements (e.g., Tilman, 1981; Borchardt, 1996; Francouer, 2001), and assemblage composition may serve as a more sensitive indicator of habitat quality than bulk biomass does (Stelzer & Lamberti, 2001). Algal community composition has been used to indicate trophic statuses of rivers and lakes, as well as measure quality of water with regards to heavy metals and other pollutants (Kelly et al., 1998).
In Hawaiian streams, algal community composition has not been explored to date as an indicator of water quality. Benthic algae are the main sources of autochthonous primary production in Hawaiian streams, along with semi-aquatic mosses. To date, little research has been conducted on stream algae on Pacific islands, other than diatoms in Hawaii, and most information comes as a result of survey expeditions or studies of water quality (Resh & DeSzalay, 1995; Filkin et al., 2003). Algal surveys in Hawaii and other Pacific islands have provided information on stream floras; for example, Sherwood (2004a, b) lists 390 non-marine diatom taxa that have been found in Hawaii since 1876. Benthic algae are the primary food source for several native stream fish species (Kido, 1996a, b), and algal assemblage structure may affect the availability of food for native fish. Benthic algae also provide nutrients for native invertebrates (Williams, 1939; Hardy, 1960; Capps et al., 2009).
A few studies have been conducted on benthic Hawaiian stream algae communities. Based upon a field study, Laperriere (1995) suggested that algae in Hawaiian streams were nitrogen limited, while a nutrient addition experiment by Larned & Santos (2000) found phosphate limitation. Studies of algal dynamics in Hawaiian streams have primarily focused on bulk productivity measurements, rather than assemblage composition (Laperriere, 1995; Larned & Santos, 2000; Wolff, 2000). One study to date has examined algal dynamics using several environmental parameters simultaneously (Larned & Santos, 2000). Land use in Hawaiian watersheds plays an important role in stream nutrient levels (Anthony et al., 2004); for example, nitrate and ammonium concentrations are often elevated in agricultural areas (HCPSU, 1990). Under short-lived flood events, concentrations of nitrogen and phosphorus can also increase substantially (Ringuet & Mackenzie, 2005).
In well-shaded streams, light is expected to be the primary limiting factor on algal growth, after which nutrient levels and other factors are expected to influence the growth of individual species (Hill, 1996; Stevenson, 1997; Hill & Fanta, 2007). Given the shade created in closed-canopy reaches, such as those occurring in the upper portions of many Hawaiian streams, light may be sub-optimal. Consequently, nutrient enrichment may not affect gross algal productivity (Borchardt, 1996; Stevenson, 1997; Mosisch et al., 2001; but see Taulbee et al., 2005 for evidence of nutrient and light co-limitation). However, algal species' composition may be significantly affected by nutrient addition even if there are no substantial changes in gross productivity (Tilman, 1981). Thus, assemblage composition can be a sensitive indicator of water quality in both shaded and unshaded streams.
Nutrient amendment experiments are often used to determine which nutrient(s) may limit algal growth in a stream (e.g., Pringle & Bowers, 1984; Lowe et al., 1986; Tate, 1990; Mosisch et al., 2001; Stelzer & Lamberti, 2001; Tank & Dodds, 2003). Nitrogen and phosphorus are predicted to be the most likely macronutrients to limit algal growth, though other nutrients have been found to limit growth under certain conditions; for example, low levels of silicate may limit diatom growth (Borchardt, 1996). Nutrient-diffusing substrates (NDS) deployed in a stream provide a point-source method of nutrient enrichment, and allow the general detection of assemblage response to nutrient addition (Pringle & Triska, 1996). They also allow experimental replication over comparable substrates either within or across streams.
The present study used NDS to test the effects of nitrogen and phosphorus addition on benthic algae in a Hawaiian stream at two different light levels, shaded (with a closed forest canopy) and unshaded (with an open canopy). We specifically test four hypotheses: (1) algal biomass and productivity will increase with nutrient addition above ambient levels under non light-limited (unshaded) conditions; (2) under light-limited (shaded) conditions, nutrient limitation will be replaced by light limitation; (3) algal assemblages will differ between shaded and unshaded sites; and (4) algal assemblages will change in response to nutrient addition at both sites.
Materials and methods
Study site
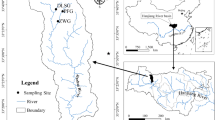
Experiments were conducted in Waihee Stream, Oahu Island, Hawaii, a perennial, groundwater fed first-order stream 2.5 km long, with a drainage area of approximately 1.6 km2. It originates in forest reserve land at an elevation of approximately 300 m, and runs through a rural residential area before terminating in a marsh and estuary. The stream substrate is variable, ranging from small pebbles to large boulders; study sites were located in segments with predominately cobble-sized substrates. A segment of the lower reach is channelized, and dense non-native riparian vegetations such as guava (Psidium guajava) are present along much of the length of the stream. Study sites were located just upstream of the forest reserve boundary (approximately 21°27′N, 157°51′W) at 52 m elevation (Fig. 1). The extent to which different algal species occur in different light environments in the study stream is unknown.

Location of Waihee stream on Oahu, and location of study sites and USGS gage (inset). Gray contour lines represent approximately 61 m of elevation. Shaded area in inset shows the approximate extent of forest reserve land. Non-shaded area in inset shows extent of rural residential land use
Waihee Stream does not exhibit strong seasonal differences in flow (mean monthly discharge from 1975 to 2000 ranged from about 0.10 to 0.21 m3 s−1; Oki & Brasher, 2003), but high flow events are frequent and can occur in any month. Yearly peak stream flow recorded during flood events during 1975–1999 ranged from ~7 to 44 m3 s−1 (USGS, 2001). Mean monthly rainfall from June 2001–June 2003 (including the time of this study) was 16.9 cm [SD = 10.9 cm; data from NOAA rainfall gage #HI-30, located in Waihee valley (NOAA, 2009)]. Table 1 summarizes key environmental and chemical parameters during March 1999 to February 2001.
Field methods
Two field experiments were conducted: ”May 12–June 14 (“spring”), and September 14–October 5, 2002: (“autumn”). Algae were allowed to colonize the outer surfaces of inverted 8.9 cm clay flowerpots with the wide end sealed with Plexiglass™, and the drainage hole plugged with a rubber stopper. After filling with appropriate fertilizer, pots were attached to concrete cinder blocks (two pots per block, with nutrient treatments randomized). The cinder blocks were then partially buried in gravel in the streambed to a depth that allowed each pot to be submerged. Pots were allowed to fill with water by removing the plug when they were placed in the stream, then they were re-plugged. One-half of the 48 pots (in the spring experiment) and one-half of the 24 pots (in the autumn experiment) were placed at an unshaded site and half at a shaded site 50 m upstream (Fig. 2). No attempt was made to control herbivory at the study site; grazing insect larvae were noted in algae samples, and manually removed before sample processing. Because the rate of nutrient release from nutrient-diffusing pots may quickly decline over time (e.g., Pringle & Bowers, 1984; Smith et al., 2001; Stelzer & Lamberti, 2001), pots were re-supplied with fertilizer after 2 weeks during each experiment.

Schematic of study sites, showing cinder block placement (open squares) and overhead canopy (filled ovals)
One-quarter of pots were assigned to each nutrient treatment, which included: (1) 110 g of NaNO3 fertilizer (N-treatment), (2) 100 g of triple superphosphate fertilizer (P-treatment), (3) both NaNO3 and PO4 (nitrate + phosphate-treatment), and (4) a non-enriched control. One-hundred g of nitrate fertilizer and 70 g of phosphate fertilizer were added to refill appropriate pots after 2 weeks during each experiment (due to lower solubility of phosphate fertilizer, more phosphate fertilizer remained in the pots over time than nitrate fertilizer, hence the smaller volume added when refilling). To refill pots, each cinder block/pot assemblage was removed from the water, the rubber stopper in each pot was removed, and the appropriate fertilizer added for each nutrient treatment with the aid of a funnel. Pots were then replaced in the stream in the same positions. Control pots were handled the same way, but no fertilizer was added. Nutrient release rates (nitrate, ammonium, and phosphorus) were estimated six times during the spring experiment by removing 250 ml of stream water with a plastic 250-ml syringe just downstream of three pots.
Water temperature was logged every 20 min at each site over the course of each experiment using a single I-Button temperature logger (Dwyer Instruments). Water velocity was measured twice during each experiment at the upstream side of each pot using a Marsh McBirney flow meter. Depth of pots in the water (top of pot to surface) was also recorded on these dates. Stream flow was determined every 2–5 days from a permanent USGS stage gage (Gage #16284200; data from USGS, 2001).
Light levels at each site were measured in Lumens m−2 twice during each experiment with HOBO-LI data loggers (onset computer) placed in waterproof casings on the stream bank within 2 m of the pots. HOBO-LI data loggers were calibrated with a LI-COR quantum sensor (LI-COR, serial #Q23667), and light levels were converted into photosynthetically active radiation (μm m−2 s−1) using the linear regression calculated from the calibration data: (μm m−2 s−1) = −669.994 + 362.138 (log Lumens m−2) (df = 6, P < 0.01, R 2 = 0.77). Canopy cover was measured for each pair of pots using a spherical densiometer held at stream level. Relative monthly solar exposure at each site was measured using a Solar Pathfinder (Solar Pathfinder, Iron City, TN).
During the autumn experiment, a pulse amplitude-modulated (PAM) fluorometer (“Diving-PAM”, Heinz Walz GmbH) was used to determine electron transport rate (ETR) via rapid light response curves for the experimental algal communities. When ETR is graphed against photosynthetically active radiation (PAR), it approximates a photosynthesis versus irradiance (P vs. I) curve as measured by traditional laboratory methods such as oxygen electrodes; however, oxygen evolution is the more accurate measure (Nielsen & Nielsen, 2008). The utility of this method is that it is rapid and can be implemented in the field without removing the organism or assemblage from in situ habitats (e.g., Hiriart-Baer et al., 2008). Fluorescence was measured at one point on each pot, located midway from the top and between the upstream and downstream sides. Ambient light was excluded from the sensor by placing a tube of black foam pipe insulation around the sensor clip, and pressing the foam firmly to the pot surface. ETR was fitted to the nonlinear photosynthetic response curve of Platt and colleagues (in Hobson & Fallowfield, 2001). Two photosynthetic parameters were calculated to quantify physiological response to light: P max, the maximum rate of photosynthesis, and E k, the level of irradiance at which maximum photosynthetic rate is reached.
Sample processing
During the spring experiment, pots were sampled after 2 and 4 weeks of colonization. Three pots of each treatment type per site were randomly selected for removal on each sampling date (four nitrate + phosphate and two nitrate pots were removed at the unshaded site on the first sampling date in error). Because algae growth was sparse after 2 weeks, the 2-week data were used only for biomass calculations, not diatom species composition; consequently, all species composition data reported here come from the 4-week sampling date. During the autumn experiment, all pots were removed after 3 weeks. Because of an impending storm with forecasted potential to wash out the experiment, the autumn experiment was ended after 3 weeks. During the autumn experiment, periphyton from three rocks at each site was scrubbed off with a plastic brush, combined for each site, and transported to our laboratory for species identification.
Individual pots were retrieved from the stream, put into plastic bags, placed on ice, and transported to our laboratory. The entire surface of each pot, excluding the top (~144.7 cm2), was gently scrubbed with a brush into 300 ml distilled water until all visible periphyton was removed. Any filamentous cyanobacteria which were present on the pots readily disintegrated when touched; some biomass was unavoidably lost from the pot surfaces during transport to the lab for analysis. Samples were shaken to homogenize contents and divided into three 100-ml subsamples for (1) ash-free dry mass (AFDM), (2) chlorophyll a, and (3) diatom species identification. Algal species identification samples were preserved with about 4.5 ml of 4% formalin. Chlorophyll samples were kept on ice and in the dark until processed. The maximum holding time before processing was approximately 4 h.
For AFDM measurement, either 100 or 50 ml of sample (depending on quantity of algae present) was filtered onto pre-ashed and preweighed Whatman GF/F or Millipore 0.7 μm filters. Filters were individually loosely wrapped in aluminum foil and placed in a 105°C drying oven for 24 h, and weighed with a precision balance. Dry mass was then recorded. Filters were then ashed in a muffle furnace (model # M15A-1A, Blue M Co., IL) at 500°C for 1 h, after which ash-free mass was determined. This procedure follows Steinman & Lamberti (1996), who do not specify rehydrating samples to replace water of hydration, then drying until constant weight. Therefore, AFDM may have been overestimated.
For chlorophyll analysis, either 50 or 100 ml of sample was filtered (depending on quantity of algae present). Samples were kept on ice in the dark until filtration. Filters were placed into individual screw-top test tubes covered in aluminum foil. Ten milliliters of a 90% acetone (90 acetone: 10 buffered MgCO3 in dH2O) solution was added. Pigments were extracted for 24 h at 4°C without grinding filters, following Steinman & Lamberti (1996). Sample absorbance was measured using a HP 8452A Diode Array spectrophotometer following standard methods (APHA, AWWA & WEF, 1992).
For diatom identification, 50–75 ml of each algal subsample was oxidized by heating for 1 h with 20 ml of 1:1 50% hydrogen peroxide and 50% nitric acid under a fume hood. Samples were spun down in a Centra MP4R (IEC) centrifuge for 9 min at 1200 rpm, and then excess water was poured off, and distilled water added. This process was repeated until the samples tested neutral with a pH paper indicator, as modified from Patrick & Reimer (1966). Samples were then diluted to more or less equal concentrations and plated out onto coverslips to dry overnight. Two permanent slides of each sample were mounted using Naphrax mounting medium (Northern Biological Supply), with a refractive index of 1.74.
A total of 250 valves were identified on each slide using an Olympus BX-40 microscope at 100× magnification, to obtain a total of 500 valves per sample. Identifying a minimum of 200 valves per sample is recommended to provide enough information to evaluate water quality (Bare & Newall, 1998). Cell counts were conducted on samples of the spring sampling date of the first experiment and the autumn sampling date of the second experiment. References used to identify diatom species included Hustedt (1937), Patrick & Reimer (1966, 1975), Fungladda et al. (1983), Dodd (1987), Foged (1987), Simonsen (1987), Cassie (1989), Krammer & Lange-Bertalot (1991, 1999a, b, 2000) and Reichardt & Lange-Bertalot (1991).
Data analysis
Analyses of biomass (AFDM and chlorophyll a), fluorescence, physical factors, and diatom species distributions were carried out using the Minitab statistical package (Minitab Inc., 1998). Chlorophyll and AFDM data were log-transformed. Two-factor analyses of variance (ANOVA; general linear model procedure, with pairwise comparisons using a Tukey’s test where appropriate) with nutrient treatment and light were performed on chlorophyll a levels, AFDM, and fluorescence parameters for each experiment. Univariate ANOVA was performed on the physical factors of canopy cover, water velocity, and pot depth with response variables of AFDM and chlorophyll a. Diatom species distributions were measured using t tests using a Bonferroni correction to account for multiple comparisons.
Multivariate analysis of diatom species composition was carried out using Primer-E (Clarke & Gorley, 2001). Analysis of similarity (ANOSIM) and non-metric multidimensional scaling (NMDS) were performed on non-transformed cell counts. ANOSIM allows the calculation of statistical significance using a permutation test. NMDS provides a graphical representation of the distance between samples based on Bray–Curtis similarity. Diatom assemblage diversity was represented with the Shannon–Wiener heterogeneity statistic (H′), which is sensitive to rare species, and Simpson’s Index, which is sensitive to common species. Diversity was compared between light and nutrient treatments and dates, as well as between communities scrubbed from natural rocks in October and those on the experimental pots.
Results
Environmental factors
Water temperature remained between 19 and 21°C at both the unshaded and shaded sites during both experiments. Light levels between the unshaded and shaded sites were significantly different (t test, mean difference between sites 69.56 μm m−2 s−1, T = 17.64, P < 0.001). The shaded site received no direct sunlight, but did receive filtered light; in contrast, the unshaded site received direct sunlight. Canopy cover was significantly higher at the shaded site than the unshaded site (mean densiometer readings 14.9 and 6.0, respectively; ANOVA, F = 362.87, P < 0.001). Average day length in spring was 13 h, 20 min, and percent available solar radiation was 8.3% at the shaded site and 64.5% at the unshaded site. Average day length in autumn was 12 h, 5 min, and percent available solar radiation was 10.3% at the shaded site and 45.8% at the unshaded site (Table 1).
Current velocity was variable among pots (mean velocity = 0.46 m s−1, SD = 0.21 m s−1) and was not significantly different between pots in the shaded and unshaded sites or among nutrient treatments (ANOVA, F = 0.41, P = 0.523 and F = 0.59, P = 0.625, respectively). Depth of individual pots in the water varied less than water velocity (mean depth of water measured from top of pot = 11.7 cm, SD = 2.2 cm), and was not significantly different between sites or nutrient treatments (ANOVA, F = 0.81, P = 0.371 and F = 0.96, P = 0.416). Stream flow was higher during the spring experiment than in the autumn experiment (mean flow = 0.187 and 0.137 m3 s−1, respectively). By comparison, flow during an April 2002 flood event was greater than 4.7 m3 s−1 (USGS, 2001).
Neither average pot depth nor average water velocity were correlated with biomass (ANOVA, P = 0.405 and 0.203, respectively). However, the spring 2-week sample had greater biomass on pot surfaces that had lower water velocities (ANOVA, P = 0.030), while on the 4-week sampling date, biomass had attained a maximal level and did not change further with pot depth or water velocity.
There was no temporal pattern in nutrient release rates as measured less than 1 cm downstream from pot surfaces, and mean nutrient release levels did not correspond to treatment type (Table 2). Mean nutrient concentrations across all treatment types and sampling dates were: nitrate (NO3 −): 0.201 μg/l (mean SD = 0.056), phosphorus (P): 0.092 μg/l (mean SD = 0.120), and ammonium (NH4 +): 0.065 μg/l (mean SD = 0.015).
Biomass
Table 3 shows the results of AFDM determination. Levels of AFDM were significantly higher at the unshaded site than at the shaded site (ANOVA, F = 33.42, P < 0.001). AFDM also varied significantly with experiment (F = 17.57, P < 0.001). Tukey multiple comparisons by date indicated that the 4-week sampling date of the spring experiment had significantly higher AFDM than either the 2-week sampling date (T = 6.84, P < 0.001) or the autumn (3-week) experiment (T = 8.60, P < 0.001). Although neither the nutrient treatment nor nutrient treatment by shading interaction were statistically significant (F = 0.825, P = 0.486 and F = 1.91, P = 0.138, respectively), this was one of the experimental hypotheses, and Tukey multiple comparisons were calculated (significant results at the 95% level shown in Table 4).
Chlorophyll a levels were significantly higher at the unshaded site than at the shaded site (ANOVA, F = 22.23, P < 0.001). The effects of nutrient treatment on chlorophyll a content were also significant (F = 3.01, P = 0.038), as were sampling dates (F = 34.81, P < 0.001). Table 5 shows the results of chlorophyll a determination.
Tukey multiple comparisons showed that chlorophyll a levels were significantly higher with nitrate treatment than phosphate treatment (F = 2.801, P = 0.035). Further, chlorophyll a levels on the 4th week of the spring experiment were significantly higher than those of the 2nd week of the spring experiment F = 8.244, P < 0.001). Although not directly comparable, chlorophyll a levels for the 3rd week of the autumn experiment were higher than those of the 2nd week of the spring experiment (F = 3.039, P < 0.001), and lower than those of the 4th week of the spring experiment (F = 5.266, P < 0.001). Tukey multiple comparisons were calculated (significant results at the 95% level shown in Table 6).
Assemblage composition
A total of 66 diatom taxa were identified on pots and natural substrates. Of the dominant taxa, two were centric diatoms: Melosira varians Ag. and Hydrosera whampoensis (Schwartz) Deby, and 13 taxa were motile: Bacillaria paradoxa Gmelin, 11 Navicula taxa, and Sellaphora pupula (Kutz.) Meresch. The remaining 51 taxa were either solitary or colonial adnate species. Cell volumes of taxa observed during cell counts ranged from 32 to 13,119 μm3 (mean size 588 μm3; SD = 9,290 μm3). The dominant non-diatom taxa included filamentous Chlorophytes (Cloniophora spp.) and Cyanophytes (Oscillatoria spp.).
Proportional cell numbers of several taxa were significantly different with either nitrate enrichment or shading (Table 7). Three diatom taxa responded significantly to nitrate addition (Gomphonema minutum (Ag.) Ag., Navicula gregaria Donkin, and Nitzschia amphibia Grun.), and thirteen taxa to shading (Achnanthes biasolettiana (Kutz.) Grun., Achnanthes lanceolata (Breb.) Grun., Achnanthes linearis (W. Smith) Grun., Achnanthes rupestoides Hohn, Fragilaria capucina var. perminuta (Grunow) Lang.-Bert., Gomphonema parvulum (Kutz.) Kutz., Gomphonema sp. “A”, Navicula bryophila Peters. & Ostr., Navicula menisculus var. upsaliensis Grun., Navicula spp. 2, Rhoicosphenia curvata (Kutz.) Grun. ex Rabh., Sellaphora pupula (Kutz.) Meresch., Synedra ulna Ehr.). Eight of these taxa were common (occurred in more than 75% of samples), and all taxa occurred in more than 30% of samples (Table 7). No taxon responded significantly to phosphate enrichment.
Two analyses of similarity (ANOSIM) were performed. In the first ANOSIM, data were split by experiment and tested across light treatment and nutrient treatment. The main effect of light treatment was significant for both experiments (spring: R = 0.505, P < 0.01; autumn: R = 0.500, P < 0.01). The main effect of nutrient treatment was not statistically significant, but significant comparisons of nitrate and control nutrient treatments were significant for both experiments (spring: R = 0.426, P < 0.05; autumn: R = 0.370, P < 0.05). No other nutrient comparisons were significant.
In the second ANOSIM, data were split by light treatment and tested across nutrient treatment and experiment. The main effect of experiment was significant for both light treatments (Unshaded: R = 0.667, P < 0.001; Shaded: R = 0.769, P < 0.001). The main effect of nutrient was not statistically significant, but significant comparisons of nitrate and control nutrient treatments were significant for both experiments (Unshaded: R = 0.389, P < 0.05; Shaded: R = 0.769, P < 0.001). No other nutrient comparisons were significant.
Diatom species composition on pots was significantly different between experiments (Fig. 3). When assemblage data from the spring (4-week sampling date) and autumn experiments were separated, there was considerable overlap in diatom species composition among nutrient treatments (compare Fig. 4a, c). However, data showed more consistent differences between light treatments (compare Fig. 4b, d).

Non-metric multidimensional scaling ordination of diatom species after different experiments. The stress value of 0.14 for the analysis indicates a fair representation of assemblage composition

Non-metric multidimensional scaling ordination of diatom species, separated by experiment. Assemblage from the spring experiment a under different nutrient treatments, and b under different light treatments. Assemblage from the autumn experiment c under different nutrient treatments, and b under different light treatments. The stress values of 0.11 for the spring analysis and 0.15 from the autumn indicate fair representations of assemblage composition
Analysis of similarity was also performed to examine whether diatom assemblages on experimental pots and stream rocks differed. Two-way ANOSIM of cell counts between substrate and light treatment had similar results, and showed significant differences between light treatment (R = 0.247, P < 0.001) as well as between substrates (R = 0.367, P < 0.05).
As measured by the Shannon–Wiener H′ statistic, algal assemblage diversity of samples from the shaded site (from both experiments and stream rocks) was significantly greater than that of the assemblage on natural rocks sampled at the unshaded site. There were also significant differences in algal assemblage diversity at the unshaded site between the autumn experiment and all other samples (Fig. 5). There were no significant differences in algal assemblage diversity among nutrient treatments using either the Shannon–Wiener H′ statistic or Simpson’s Index (data not shown).

Diatom assemblage diversity as measured by Shannon–Wiener H′ at unshaded and shaded sites. Bars represent standard deviation
Patterns of diversity as measured by Simpson’s Index were similar to those measured by H′. However, the only significant difference between any site or substrate was that between the natural rock assemblage at the unshaded site and all other samples except the spring assemblage at the unshaded site.
PAM fluorescence measurements
Values of E k for algal communities were significantly higher at the unshaded site than at the shaded site (F = 21.25, P < 0.001). Elevated E k values were also correlated with nitrate enrichment (ANOVA, F = 3.04, P = 0.059) and the shading and nutrient treatment interaction (F = 2.75, P = 0.077). Tukey multiple comparisons of effects of nutrient treatment indicated that E k was significantly higher with nitrate treatment than the control (T = 2.868, adjusted P < 0.05). P max was significantly higher at the unshaded site than the shaded site (F = 17.41, P < 0.001). P max did not differ significantly with nutrient treatment (F = 1.84, P = 0.180); however, P max differed significantly with the interaction of shading and nutrient treatment (F = 3.50, P < 0.05).
Discussion
Effects of light and nutrients on algal biomass and fluorescence
One unexpected finding of our study after our initial spring experiment was that biomass (AFDM and chlorophyll a) measurements did not behave as predicted for our control nutrient treatments during the 4-week sampling date. During the 2-week sampling date of the spring experiment, biomass was lower on pots of all nutrient treatments at the shaded site than on the pots at the unshaded site, as would be expected if light were limiting algal growth. However, during the 4-week sampling date of the spring experiment, biomass was higher on the shaded control pots than the unshaded control pots (for all other nutrient treatments, biomass was lower on the shaded pots than the unshaded pots). This finding is difficult to explain. There was no correlation among water velocity, canopy cover, and pot depth at the control pots at the two study sites that might explain this difference. Because of this unexpected result, we added an additional experiment to our study in the autumn. We did not detect this anomaly in our second experiment. A possible mechanism to explain this pattern is discussed later in “study limitations” part of this section.
In this study, algal biomass (AFDM and chlorophyll a) and at least some diatom species responded positively to nitrate enrichment when light levels were at 45% or higher, as found at the unshaded site. This suggests that the periphytic assemblage in Waihee Stream is primarily light-limited, and secondarily nitrogen-limited. This conclusion supports our first two hypotheses (that biomass will increase with nutrient addition, unless limited by light).
Significant differences in biomass (AFDM and chlorophyll a) at high light levels were also observed for the nitrate plus phosphate treatment in this study. However, there was less evidence overall for the positive effect of this treatment than that of the nitrate treatment alone. One possible explanation for this is that phosphate fertilizers commonly contain toxic metals (Charter et al., 1995), which could have negatively affected algal growth; our fertilizer samples were not tested for metals. Another possibility is that the phosphorus was adsorbed to minerals in the clay pots, thus less was released (Pringle, 1987).
When PAM fluorescence was performed on the periphytic assemblage in the autumn experiment, E k and P max were significantly higher at the unshaded site than the shaded site. P max is the maximum rate of photosynthesis, and is expected to increase in plants that live at higher light levels. T, the irradiance level at which maximum photosynthesis is reached (E k), will increase in the same way in response to light exposure (Hill, 1996). For these parameters, the results of this experiment behaved as predicted. PAM fluorescence results support the suggestion from the biomass data that the benthic algal assemblage is more light-limited than nitrogen-limited in Waihee Stream. Interactions of nutrient enrichment and light showed higher E k and P max levels at the unshaded site, when either nitrate, or to a lesser degree, nitrate plus phosphate enrichment took place. These results suggest that photosynthetic capacity was enhanced under a non-light-limited situation with the addition of a limiting nutrient—in this case nitrogen. Similar interactive effects between light and nutrient limitation have been observed in other studies for nitrogen (Lapointe & Tenore, 1981) and phosphorus (Hiriart-Baer et al., 2008).
Effects of light and nutrients on diatom assemblages
Diatom assemblage compositions were significantly different between shaded and unshaded sites, supporting our third hypothesis: that assemblages will differ between shaded and unshaded sites. Differences in assemblage composition between experiments also were detected. Differences in assemblage composition between experiments could have risen from the experiment lengths (4 weeks in spring vs. 3 weeks in autumn), or from random variation in colonizing species. Combining these observations with the PAM fluorescence data at the shaded and unshaded sites, we suggest that moderate shaded and unshaded assemblages of diatoms and other benthic algae occupy discrete stream habitats, changing dynamically with abiotic factors.
In this study, large differences in diatom assemblage composition were seen for light, and nutrient effects were secondary. Our fourth hypothesis predicted that algal assemblages would change in response to nutrient addition at both the shaded and unshaded sites. Comparisons between nitrate treatment and at least one other nutrient treatment were significantly different in all but one analysis, suggesting partial support for this hypothesis. Three diatom taxa responded significantly to nitrate addition, and 13 taxa to shading. These taxa have the potential to be used as indicator species elsewhere in Hawaii, although additional research should be carried out in different streams to confirm the results obtained here.
As measured by Shannon–Wiener H′, diversity of the diatom assemblage was significantly lower at the unshaded site on the 4-week sampling date of the spring experiment than at either the shaded site on the same date or at either site on the 3-week sampling date in autumn. This result suggests that certain species within the algal assemblage were beginning to dominate the substrate and outcompete other species at the unshaded site. This process may be expected as an algal assemblage matures and resources become scarcer (McCormick, 1996). A similar pattern of reduced diversity at the unshaded site was observed on natural rocks sampled during October. This suggests that, by the 4th week, assemblage dynamics on clay pots was similar to that on natural substrates.
Study limitations
We recognize several limitations in this study. First, because both study sites were located in a single stream, results cannot be generalized to other Hawaiian streams (Hurlbert, 1984). In order to avoid cross-treatment contamination, the relative position of treatments should be considered carefully in the design of NDS studies (Tate, 1990). In this study, there were two potential sources of cross-contamination: among NDS within each site, and between the upstream (shaded) and downstream (unshaded) sites. We did not measure significantly higher nutrient concentrations in the water collected at the unshaded site than at the shaded site, suggesting that between-site contaminations had minimal effect. In a similar study, Tate (1990) did not detect increased nutrient concentrations downstream from nutrient-diffusing substrata. Within each site in our study, NDS treatments were randomized, and water samples were not fine-grained enough to allow us to detect possible cross-treatment contamination. Additional replication of study sites within the stream could have strengthened our conclusion that upstream–downstream cross-contamination was likely minimal. Cross-contamination between nutrient treatments might have explained the unexpected result that higher biomass accrued on shaded versus unshaded control treatments during the 4-week sample date of the spring experiment, while control treatments on all other sampling dates accrued more biomass at the unshaded site than the shaded site.
Second, algal assemblages may differ depending on substrate texture, leading to differences between assemblages on smooth experimental surfaces and rough natural substrates (Dudley & D’Antonio, 1991; Bergey, 2005). We saw lower assemblage diversity on rocks at the unshaded site than on pots at the unshaded site. However, diversity on the pot surfaces did not differ significantly overall from that on rocks at the shaded site. One possible explanation for this is that the periphytic assemblage on unshaded rocks had become dominated by fewer species than either the shaded communities or the assemblage on the unshaded pots, given both non-light-limited conditions and a longer time for colonization and succession to occur.
A third limitation comes from our nutrient enrichment method. Typically, NDS are constructed using nutrient-enriched agar (e.g., Pringle, 1987; Wellnitz et al., 1996), which provides controlled delivery of targeted nutrients at rates that decrease in a known, exponential fashion (Tate, 1990; Rugenski et al., 2008). Since benthic diatom community succession has been observed to take approximately 4 weeks (Tuji, 2000; Julius, 2007), nutrient enrichment from the NDS does not take place in a constant manner throughout community formation. In this study, fertilizer was used as a nutrient source to maintain a higher level of enrichment over the course of the study, following the method of Smith et al. (2001). Fertilizer in the pots did dissolve over time, although the nitrate fertilizer solubilized more quickly than the phosphate fertilizer; therefore, enrichment was taking place to some degree over the course of each experiment. We were not able to characterize nutrient release rates accurately over the course of this study; for example, measured nutrient levels did not correspond with nutrient treatment type. This may have been a consequence of the sampling method used, since the turbulent water flow around the pots may have rapidly mixed or diluted the nutrient plume even less than 1 cm downstream. We also did not characterize the rate of nutrient leaching from pots nor the attenuation of any released nutrients downstream. A sampling method like the one used by Tate (1990) might have better characterized nutrient release.
A final limitation is the result of herbivory during our study. We did not attempt to exclude grazers from our experimental pots, and grazing insect larvae were noted in our algae samples. Larvae were removed before sample processing; however, we did not attempt to quantify whether their presence was correlated with treatment type, site, season, or other environmental factors. Their presence may have affected algal accrual on pots. In addition, differences in grazer density between the shaded and unshaded sites might account for the finding that algal biomass was higher at the unshaded control pots.
Conclusions
The results of this study in a Hawaiian stream support the prediction that nutrient enrichment will enhance algal productivity when adequate light is available (Lowe et al., 1986; Borchardt, 1996). This finding has implications for riparian management in Hawaii. Most lower-elevation Hawaiian streams have been altered by both canopy removal and increased nutrient input (Timbol & Maciolek, 1978; Archer, 1982; HCPSU, 1990), making algal blooms more likely (Mosisch et al., 2001). Native Hawaiian stream fish primarily consume benthic algae, and different species are adapted to graze algae with different growth habits (Julius, 2007); the formation of large filamentous algal blooms might be detrimental to these fish species.
This study also suggests that assemblage composition of diatoms in Waihee Stream may change with nutrient enrichment and canopy removal, although further study is needed in other Hawaiian streams to determine broader patterns and detailed relationships. Diatoms are routinely sampled in indices of biotic integrity elsewhere (e.g., Kelly et al., 1998; Stoermer & Smol, 2000). In addition, in Hawaii, high silicate levels in stream water derived from the volcanic soils (Smith et al., 1981) and dense shade in natural settings may favor diatoms over other algal groups, such as green or blue-green algae. Therefore, diatom assemblages may be appropriate indicators in Hawaiian streams, and potentially other Pacific island streams.
Based upon this study, the use of algae in Hawaii for water quality monitoring is likely to be a fruitful area for further research. Algae are ideally suited for use as water quality indicators because of their lack of motility, large number of species, and rapid response to changes in stream quality as primary producers (Lowe & Pan, 1996), and they can serve as a useful complement to surveys of vertebrate or invertebrate biotic surveys of water quality. In Hawaii, a diatom index could be used to supplement the state’s two stream assessment protocols: the Hawaii Stream Visual Assessment Protocol (NRCS, 2001) and the Hawaii Stream Bioassessment Protocol (Kido, 2002), neither of which makes use of algae as biotic indicators. One important consideration if this approach is to be considered is that reference materials for diatom identification for Hawaii are somewhat scattered (e.g., Sherwood, 2004a, b). A lack of a comprehensive guide to species identification in Hawaii might make the task of identification more challenging than is the case in other areas (Round, 1991) and require a greater level of expertise with these taxa than might be the case for assessments with invertebrate taxa. Future study should investigate which species in particular have most utility as water quality indicators, as well as which species are likely to be found in specific stream habitats.
References
APHA, AWWA & WEF, 1992. Chlorophyll. Standard Methods for the Examination of Water and Wastewater, 18th edn. American Public Health Association, Washington, DC: 10.18–10.19.
Anthony, S. S., C. D. Hunt Jr., A. M. Brasher, L. D. Miller & M. S. Tomlinson, 2004. Water Quality on the Island of Oahu, Hawaii. U.S. Geological Survey Circular 1239, Reston.
Archer, K. M., 1982. Leaf Litter Decomposition in Hawaiian Streams. M.S. Thesis, University of Hawaii, Zoology Department.
Bare, N. & P. Newall, 1998. Techniques for the use of diatoms in water quality assessment: How many valves? Proceedings of the 15th International Diatom Symposium 15: 153–160.
Bergey, E. A., 2005. How protective are refuges? Quantifying algal protection in rock crevices. Freshwater Biology 50: 1163–1177.
Borchardt, M. A., 1996. Nutrients. In Stevenson, R. J., M. I. Bothwell & R. L. Lowe (eds), Algal Ecology: Freshwater Benthic Ecosystems. Academic Press, San Diego: 183–227.
Capps, K. A., C. B. Turner, M. T. Booth, D. L. Lombardozzi, S. H. McArt, D. Chai & N. G. Hariston Jr, 2009. Behavioral responses of the endemic shrimp Halocaridina rubra (Malostraca: Atyidae) to an introduced fish, Gambusia affinis (Actinopterygii: Poeciliidae) and implications for the trophic structure of Hawaiian anchialine pools. Pacific Science 63: 27–37.
Cassie, A., 1989. A contribution to the study of New Zealand diatoms. Bibliotheca Diatomologica 17: 1–266.
Charter, R. A., M. A. Tabatabai & J. W. Schafer, 1995. Arsenic, molybdenum, selenium, and tungsten contents of fertilizers and phosphate rocks. Communications in Soil Science and Plant Analysis 26: 3051–3062.
Clarke, K. R. & R. N. Gorley, 2001. PRIMERv5: User Manual/Tutorial. PRIMER-E, Plymouth.
Dodd, J. J., 1987. Diatoms: the Illustrated Flora of Illinois. Southern Illinois University Press, Carbondale.
Dudley, T. & C. D’Antonio, 1991. The effects of substrate texture, grazing, and disturbance on macroalgal establishment in streams. Ecology 72: 297–309.
Filkin, N. R., A. R. Sherwood & M. L. Vis, 2003. Macroalgae from 23 stream segments in the Hawaiian Islands. Pacific Science 57: 421–431.
Foged, N., 1987. Diatoms from Viti Levu, Fiji Islands. Bibliotheca Diatomologica 14: 1–195.
Francouer, S. N., 2001. Meta-analysis of lotic nutrient amendment experiments: detecting and quantifying subtle responses. Journal of the North American Benthological Society 20: 358–368.
Fungladda, N., I. Kaczmarska & S. R. Rusforth, 1983. A contribution to the freshwater diatom flora of the Hawaiian Islands. Bibliotheca Diatomologica 2: 1–103.
Hardy, D. E., 1960. Insects of Hawaii: Diptera-Brachycera, Vol. 10. University of Hawaii Press, Honolulu.
HCPSU (Hawaii Cooperative Park Services Unit), 1990. Hawaii Stream Assessment: A Preliminary Appraisal of Hawaii’s Stream Resources. Report R84. Prepared for Commission on Water Resource Management, Honolulu.
Hill, W., 1996. Effects of light. In Stevenson, R. J., M. I. Bothwell & R. L. Lowe (eds), Algal Ecology: Freshwater Benthic Ecosystems. Academic Press, San Diego: 121–148.
Hill, W. R. & S. E. Fanta, 2007. Phosphorus and light colimit periphyton growth at subsaturating irradiances. Freshwater Biology 53: 215–225.
Hiriart-Baer, V. P., T. J. Arciszewski, S. Y. Malkin, S. J. Guildford & R. E. Hecky, 2008. Use of pulse-amplitude-modulated fluorescence to assess the physiological status of Cladophora sp. along a water quality gradient. Journal of Phycology 44: 1604–1613.
Hobson, P. & H. Fallowfield, 2001. Effect of salinity on photosynthetic activity of Nodularia spumigena. Journal of Applied Phycology 13: 493–499.
Hurlbert, S. H., 1984. Pseudoreplication and the design of ecological field experiments. Ecological Monographs 54: 187–211.
Hustedt, F., 1937. Systematische und oekologische untersuchungen uber die diatomeen-flora von Java, Bali und Sumatra. Archiv Für Hydrobiologie 15: 131–177.
Julius, M. L., 2007. Why sweat the small stuff: the importance of microalgae in Hawaiian stream ecosystems. In Evenhuis, N. L. & J. M. Fitzsimons (eds), Biology of Hawaiian Streams and Estuaries, Vol. 3. Bishop Museum Bulletin in Cultural and Environmental Studies, Honolulu, 183–193.
Kelly, M. G., A. Cazaubon, E. Coring, A. Dell’Uomo, L. Ector, B. Goldsmith, H. Guasch, J. Hurlimann, A. Jarlman, B. Kawecka, J. Kwandrans, R. Laugaste, E. A. Lindstrom, M. Leitao, P. Marvan, J. Padisak, E. Pipp, J. Prygiel, E. Rott, S. Sabater, H. van Dam & J. Vizinet, 1998. Recommendations for the routine sampling of diatoms for water quality assessments in Europe. Journal of Applied Phycology 10: 215–224.
Kido, M. H., 1996a. Diet and food selection in the endemic Hawaiian amphidromous goby, Sicyopterus stimpsoni (Pisces: Gobiidae). Environmental Biology of Fishes 45: 199–209.
Kido, M. H., 1996b. Morphological variation in feeding traits of native Hawaiian stream fishes. Pacific Science 50: 184–193.
Kido, M. H., 2002. Hawaii Stream Bioassessment Protocol, Ver. 3.10. The Hawaii Stream Research Center, Center for Conservation Research and Training, University of Hawaii, Honolulu.
Krammer, K. & H. Lange-Bertalot, 1991. Susswasserflora von Mitteleuropa: Bacillariophyceae, Teil 4: Achnanthaceae, kritische erganzungen zu Navicula (Lineolatae) und Gomphonema. Gustav Fischer, Stuttgart.
Krammer, K. & H. Lange-Bertalot, 1999a. Susswasserflora von Mitteleuropa: Bacillariophyceae, Teil 1: Naviculaceae. Spektrum Akademischer, Heidelberg.
Krammer, K. & H. Lange-Bertalot, 1999b. Susswasserflora von Mitteleuropa: Bacillariophyceae, Teil 2: Bacillariaceae, Epithemiaceae, Surirellaceae. Spektrum Akademischer, Heidelberg.
Krammer, K. & H. Lange-Bertalot, 2000. Susswasserflora von Mitteleuropa: Bacillariophyceae, Teil 3: Centrales, Fragilaceae, Eunotiaceae. Spektrum Akademischer, Heidelberg.
Laperriere, J. D., 1995. Riffle algal ecology of small streams during the rainy season: Islands of Hawaii, Maui and Oahu. Tropical Ecology 36: 59–72.
Lapointe, B. E. & K. R. Tenore, 1981. Experimental outdoor studies with Ulva fasciata Delile. I. Interaction of light and nitrogen on nutrient uptake, growth, and biochemical composition. Journal of Experimental Marine Biology and Ecology 53: 135–152.
Larned, S. T. & S. R. Santos, 2000. Light- and nutrient-limited periphyton in low order streams of Oahu, Hawaii. Hydrobiologia 432: 101–111.
Lowe, R. L., S. W. Golladay & J. R. Webster, 1986. Periphyton response to nutrient manipulation in streams draining clearcut and forested watersheds. Journal of the North American Benthological Society 5: 221–229.
Lowe, R. L. & Y. Pan, 1996. Benthic algal communities as biological monitors. In Stevenson, R. J., M. I. Bothwell & R. L. Lowe (eds), Algal Ecology: Freshwater Benthic Ecosystems. Academic Press, San Diego: 705–739.
McCormick, P. V., 1996. Resource competition and species coexistence in freshwater benthic algal assemblages. In Stevenson, R. J., M. I. Bothwell & R. L. Lowe (eds), Algal Ecology: Freshwater Benthic Ecosystems. Academic Press, San Diego: 229–252.
Minitab, Inc, 1998. MINITAB Ver. 12.2. State College.
Mosisch, T. D., S. E. Bunn & P. M. Davies, 2001. The relative importance of shading and nutrients on algal production in subtropical streams. Freshwater Biology 46: 1269–1278.
NOAA (National Oceanic and Atmospheric Administration), 2009. Hawai`i Archived Hydronet Data [Accessed 9 June 2009 from http://www.prh.noaa.gov/hnl/hydro/hydronet/hydronet-data.php].
NRCS (Natural Resources Conservation Service), 2001. Hawaii Stream Visual Assessment Protocol, Ver. 1.0. Natural Resources Conservation Service, Honolulu.
Oki, D. S. & A. M. D. Brasher, 2003. Environmental Setting and the Effects of Natural and Human-Related Factors on Water Quality and Aquatic Biota, Oahu, Hawaii. United States Geological Survey Water-Resources Investigations Report 03-4156.
Patrick, R. & C. Reimer, 1966. Diatoms of North America, exclusive of Alaska and Hawaii: Fragilariaceae, Eunotiaceae, Achnanthaceae, Naviculaceae, Vol. 1. Monographs of the Academy of Natural Sciences of Philadelphia, Philadelphia.
Patrick, R. & C. Reimer, 1975. Diatoms of North America, Exclusive of Alaska and Hawaii: Entomoneidaceae, Cymbellaceae, Gomphonemaceae, Epithemiaceae, Vol. 2, Part 1. Monographs of the Academy of Natural Sciences of Philadelphia, Philadelphia.
Pringle, C. M., 1987. Effects of water and substratum nutrient supplies on lotic periphyton growth: an integrated bioassay. Canadian Journal of Fisheries & Aquatic Science 44: 619–629.
Pringle, C. M. & J. A. Bowers, 1984. An in situ substratum fertilization technique: diatom colonization on nutrient-enriched, sand substrata. Canadian Journal of Fisheries & Aquatic Science 41: 1247–1251.
Pringle, C. M. & F. J. Triska, 1996. Effects of nutrient enrichment on periphyton. In Hauer, F. R. & G. A. Lamberti (eds), Methods in Stream Ecology. Academic Press, San Diego: 607–623.
Nielsen, H. D. & S. L. Nielsen, 2008. Evaluation of imaging and conventional PAM as a measure of photosynthesis in thin- and thick-leaved marine macroalgae. Aquatic Biology 3: 121–131.
Reichardt, E. & H. Lange-Bertalot, 1991. Taxonomische revision des artenkomplexes um Gomphonema angustum – G. dichotomum – G. intricatum – G. vibrio und ahnliche Taxa (Bacillariophyta). Nova Hedwigia 53: 519–544.
Resh, V. H. & F. A. DeSzalay, 1995. Streams and rivers of Oceania. In Cushing, C. E., K. W. Cummins & G. W. Minshall (eds), Ecosystems of the World 22: River and Stream Ecosystems, Chap 23. Elsevier Press, New York.
Ringuet, S. & F. T. Mackenzie, 2005. Controls on nutrient and phytoplankton dynamics during normal flow and storm runoff conditions, southern Kaneohe Bay, Hawaii. Estuaries 28: 327–337.
Rugenski, A. T., A. M. Marcarelli, H. A. Bechtold & R. S. Inouye, 2008. Effects of temperature and concentration on nutrient release rates from nutrient diffusing substrata. Journal of the North American Benthological Society 27: 52–57.
Round, F. E., 1991. Diatoms in river water-monitoring studies. Journal of Applied Phycology 3: 129–145.
Sherwood, A. R., 2004a. Bibliographic checklist of the non-marine algae of the Hawaiian Islands. Bishop Museum Occasional Papers Series 80: 1–23.
Sherwood, A. R., 2004b. New records of freshwater macroalgae and diatoms from the Hawaiian Islands. Records of the Hawaii Biological Survey 79: 1–8.
Simonsen, R., 1987. Atlas and Catalogue of the Diatom Types of Friedrich Hustedt. J. Cramer, Berlin.
Smith, J. E., C. M. Smith & C. L. Hunter, 2001. An experimental analysis of the effects of herbivory and nutrient enrichment on benthic community dynamics on a Hawaiian reef. Coral Reefs 19: 332–342.
Smith, S. V., W. J. Kimmerer, E. A. Laws, R. E. Brock & T. W. Walsh, 1981. Kaneohe Bay sewage diversion experiment. Perspectives on ecosystem responses to nutritional perturbation. Pacific Science 35: 279–407.
Steinman, A. D. & G. A. Lamberti, 1996. Biomass and pigments of benthic algae. In Hauer, F. R. & G. A. Lamberti (eds), Methods in Stream Ecology. Academic Press, San Diego: 295–313.
Stelzer, R. S. & G. A. Lamberti, 2001. Effects of N: P ratio and total nutrient concentration on stream periphyton community structure, biomass, and elemental composition. Limnology and Oceanography 46: 356–367.
Stevenson, R. J., 1997. Scale-dependent determinants and consequences of benthic algal heterogeneity. Journal of the North American Benthological Society 16: 248–262.
Stoermer, E. F. & J. Smol, 2000. The Diatoms: Applications for Earth and Environmental Sciences. Cambridge University Press, London.
Tank, J. L. & W. K. Dodds, 2003. Nutrient limitation of epilithic and epixylic biofilms in 10 North American streams. Freshwater Biology 48: 1031–1049.
Tate, C. M., 1990. Patterns and controls of nitrogen in tallgrass prairie streams. Ecology 71: 2007–2018.
Taulbee, W. K., S. D. Cooper & J. M. Melack, 2005. Effects of nutrient enrichment on algal biomass across a natural light gradient. Archiv Für Hydrobiologie 164: 449–464.
Tilman, D., 1981. Tests of resource competition theory using four species of Lake Michigan algae. Ecology 62: 802–815.
Timbol, A. & J. A. Maciolek, 1978. Stream Channel Modification in Hawaii. Part A: Statewide Inventory of Streams; Habitat Factors and Associated Biota. U. S. Fish & Wildlife Service, FWS/OBS-78/16.
Tuji, A., 2000. Observation of developmental processes in loosely attached diatom (Bacillariophyceae) communities. Phycological Research 48: 75–84.
USGS (United States Geological Survey), Pacific Islands Water Science Center, 2001. Site Description and Available Data: Waihee Stream Near Kahaluu [Accessed 9 June 2009 from http://hi.water.usgs.gov/studies/nawqa/site_waihee.html].
USNO (United States Naval Observatory), 2009. Data Services [Accessed 5 September 2009 from http://aa.usno.navy.mil/data/].
Wellnitz, T. A., R. B. Rader & J. V. Ward, 1996. Importance of light and nutrients in structuring an algal community in a Rocky Mountain stream. Journal of Freshwater Ecology 11: 399–413.
Williams, F. X., 1939. Biological studies of Hawaiian water-loving insects, Part III Diptera or flies. Proceedings of the Hawaiian Entomological Society 10: 281–315.
Wolff, R. H., 2000. Seasonal Recovery Patterns of Hawaiian Stream Flora and Fauna, Wainiha River, Kauai, Hawaii. M.S. Thesis, University of Hawaii, Zoology Department.
Acknowledgements
The authors would like to thank Isabella Abbott, Sarah Spaulding, Steve Main, Jennifer Smith, Mike Kido, Chris Bird, Maya Iriondo, Wendy McDowell, James Mazza, and Dan Hoover for their assistance with this project. Funding was provided by the University of Hawaii (UH) Ecology, Evolution and Conservation Biology Program, UH Arts and Sciences Advisory Council, and the UH Botany Department. The authors also thank Larry Brown, Kurt Carpenter, and Suzanne Femmer, who all provided comments on an earlier version of this article.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: David Philip Hamilton
Rights and permissions
About this article
Cite this article
Stephens, S.H., Brasher, A.M.D. & Smith, C.M. Response of an algal assemblage to nutrient enrichment and shading in a Hawaiian stream. Hydrobiologia 683, 135–150 (2012). https://doi.org/10.1007/s10750-011-0947-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-011-0947-2