
Abstract
Hydrological parameters can potentially have an overwhelming influence on sedimentary assemblages of Cladocera at certain sampling sites that can cause problems for palaeoenvironmental reconstructions. We applied a previously developed Cladocera-based inference model of water depth and a mean July air temperature transfer function developed in this study to a surface sediment dataset of fossil Cladocera from a lake in eastern Finland aiming to investigate the influence of stream flow and water depth on reconstruction results. The developed temperature-inference model, using the weighted averaging-partial least squares technique, had relatively favourable performance statistics suggesting that it is valid in means of performing temperature estimations. When the temperature model was applied to the intralake samples, the lotic samples had inferred values mostly within the model’s prediction error and only one lotic sample showed an underestimated temperature. Samples taken from depths over ~3 m inferred generally underestimated temperatures, although most of the values were within the model’s prediction error. The water depth reconstructions correlated significantly with the measured water depth, but the shallowest samples and most of the lotic samples yielded overestimated inferred values and the samples taken from depths >5 m showed underestimated values. In both reconstruction sets, the inferred values were underestimated in samples taken from deeper sites. Based on the present results, it may be recommendable that downcore sediment samples should be taken from intermediate depths, where also the diversity is higher, and deepest sites and inflows should be avoided. However, more research is needed to validate these results in a larger geographical context.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The parts of cladoceran (Crustacea: Cladocera) exoskeletons can be recovered from lake sediments. These fossil remains can be used in palaeolimnological studies to reconstruct past environmental conditions, such as lake level changes (Alhonen, 1970a, b; Hofmann, 1998; Bos et al., 1999; Korhola et al., 2005) and climate development (Lotter et al., 2000; Kattel et al., 2008), and also anthropogenic impacts (Brodersen et al., 1998; Jeppesen et al., 2001). It has been shown that the living cladoceran communities are well represented by the fossil assemblages (Frey, 1960; Nykänen et al., 2009). However, remains of other cladocerans besides Bosminidae and Chydoridae, such as Daphniidae, are selectively preserved in lake sediments (Frey, 1986; Hann, 1989; Korhola & Rautio, 2001). Therefore, Cladocera-based training sets should be developed from fossil remains from surface sediment samples instead of living samples to avoid this bias in representativeness. Furthermore, the surface sediment samples collect the species during a long time period, hence avoiding the bias caused by dynamic seasonal occurrence. Although the species composition of living communities matches that found in the fossil assemblages (Frey, 1960), there can be variation in the percentage abundances (Sarmaja-Korjonen, 2001). This variation creates a potential error source for quantitative Cladocera-based environmental reconstructions performed from single cores.
It has recently been shown that there is great within-lake variability in midge (Diptera: Nematocera) and diatom (Bacillariophyta) assemblages that can be used to develop quantitative reconstruction models for hydrological variables (Kurek & Cwynar, 2009; Laird et al., 2010; Luoto, 2010a). Therefore, it is more than likely that this approach can be applied to cladocerans as well. Although there is great potential in using intralake samples to develop calibration sets for hydrological variables, the high level of heterogeneity in lake-specific fossil assemblages can cause problems for selecting a representative sampling point for a downcore study. Because downcore studies that are based on a single sediment profile represent the same spot in the lake through time, the possible error in reconstruction would hence be systematic under- or overestimation of the examined environmental variable, e.g. temperature. However, if lake level changes would have occurred over time, this systematic trend would have disappeared and the inferred values would have potentially become seriously biased following the habitat change at the sampling point.
In this study, we aim to address the issue of sampling point selection in downcore palaeolimnological studies by investigating the influence of hydrological variables on quantitative Cladocera-based air temperature and water depth reconstructions. We use a new Finnish Cladocera-mean July air temperature (T Jul) transfer function presented in this study and apply a previously available Finnish Cladocera-based water depth reconstruction model to an intralake and -river dataset of fossil Cladocera. In so doing, we seek to find trends in the inferred temperatures and water depths along a depth gradient and assess the influence of stream flow on the quantitative estimates of these parameters. Although the major advantage of selecting the sampling point from the deepest location in a lake basin is that there is a reduced risk of running into hiatuses or sediment mixing (Walker et al., 1984), we hypothesise that the best coring location would correspond to the greatest cladoceran diversity, which most likely occurs closer to lake shores.
Materials and methods
Study site
The study lakes for the temperature calibration dataset are located in Finland along a latitudinal axis of 60–70°N (Fig. 1) covering the boreal mixed forests in the south and tundra vegetation in the north. The 78 lakes are small and shallow and were chosen for the study to represent different climatic conditions. The mean air T Jul of the study lakes varies from 17.1°C in the south to 11.3°C in the north (mean 14.9°C), and the mean annual temperature from 5.8 to −2.0°C (mean 1.8°C), respectively. The altitudinal range is from 11 to 404 m above sea level (mean 148 m) and the range in mean annual precipitation is from 400 to 660 mm. Water depth at the sampling sites varied between 0.5 and 7 m (mean 2.7 m). The study lakes are mostly the same as in Luoto (2009a), who also describes the limnological variation among the sites. Late-winter dissolved oxygen (~0.5 m from the bottom) varied in these lakes from 0.5 to 11.8 mg l−1 (mean 5.5 mg l−1), pH from 3.8 to 9.3 (mean 6.2) and conductivity from 8 to 312 μS cm−1 (mean 58 μS cm−1). However, the limnological variables were not measured from all the sites, with measurements lacking especially from the northern sites (consult Luoto, 2009a for details). The water depth calibration set consists of 55 lakes in Finland. These lakes are also included in the temperature calibration set, but include only lakes having a state close to pristine following the environmental screening by Luoto (2009b), where largest lakes and lakes located in urban or otherwise disturbed areas were deleted from the calibration set to better reflect the influence of water depth. There is no correlation between temperature and depth in the calibration sets (Luoto, 2009b) that could cause problems in separating these environmental signals (Korhola et al., 2000). The lakes are mostly dystrophic, which subjects them more easily to stratification even they are shallow. Detailed site descriptions are available in Nevalainen et al. (2011).

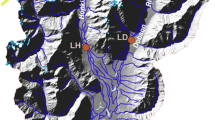
Location of the 31 intralake samples in Lake Pieni-Kauro and the Saavanjoki River, eastern Finland and the 78 multilake samples (temperature training set) along the latitudinal gradient in Finland
Lake Pieni-Kauro (64°17′ N, 30°07′ E), from where the intralake dataset was derived, is a small (~30 ha) dystrophic lake situated in Kuhmo, eastern Finland at an elevation of 188 m (Fig. 1). The mean air T Jul at the lake is 15.25°C and the annual precipitation varies between 600 and 650 mm (climate normals 1971–2000). The catchment is characterised by boreal coniferous forests and the bedrock consists of Archean granitoids and migmatites (Mikkola, 2008). The main inlet is the Saavanjoki River that enters from the north and the main outlet (Lake Kaurojärvi) is on the western side. The maximum water depth is 7.9 m and mean water depth ~3 m in Lake Pieni-Kauro and 2.3 m in the River Saavanjoki. The shoreline length is 3.8 km and the organic content of the sediment (measured as loss-on-ignition) varies from 11 (wind exposed shore) to 77% (next to an edge of peat) (see details in Luoto, 2010a). Because the lake is dystrophic, light does not penetrate very deep and thermal stratification probably occurs during the summers. The aquatic vegetation (Fig. 1) consists of Phragmites australis (Cav.) Trin ex steud, Nuphar lutea L. Sm. and Equisetum fluviatile L. and the lake has diverse and vital fish and macrobenthic communities (Luoto, 2010a).
Field, laboratory and statistical methods
Temperature training set
A Limnos gravity corer (Kansanen et al., 1991) was used to collect 78 surface sediment samples (topmost 0–1 cm representing the recent years of sedimentation) from lakes along a latitudinal gradient in Finland. The sampling was performed through ice in late winter 2005 from intermediate depths to better include littoral taxa. Fossil Cladocera analysis was performed using standard methods described by Szeroczyńska & Sarmaja-Korjonen (2007). The mean T Juls for the training set sites were estimated using a GIS-based method. The temperature data were interpolated from the data of all Finish meteorological stations to a 10 × 10 km grid, which takes into account, e.g. elevations. The temperature data based on the climate normals 1971–2000 was provided by the Finnish Meteorological Institute.
The gradient lengths of detrended correspondence analysis (DCA) were used to determine whether apply unimodal or linear methods in exploring the relationship between surface sediment Cladocera assemblages (relative abundances) and climate factors. Canonical correspondence analysis (CCA) was used to examine the relationship between the surface sediment Cladocera assemblages and environmental variables with Monte Carlo permutations (999 unrestricted permutations) to test the statistical significance. The CCAs were run one environmental variable at a time with square-root transformation of the species data and down-weighting of rare species. Because limnological data was not available from all the sites, it was not possible to use co-variables to test the strength of climatic variables against limnology. The DCA and CCA analyses were performed with the program CANOCO, version 4.52 (ter Braak & Šmilauer, 2002).
The method of weighted averaging-partial least squares (WA-PLS) was used to develop a Cladocera-mean air T Jul transfer function from the multilake surface sediment data. Also other model types were tested, such as WA using classical and inverse deshrinking regression and PLS. The species data was log10 transformed to stabilize the variance, but no rare taxa deletions were performed to better include all indicator taxa. The temperature gradient of the training set ranges from 17.0 to 11.3°C that makes it well suitable for this study lake (mean T Jul = 15.3°C). The optimal number of components in the Cladocera-mean air T Jul transfer function was assessed using leave-one-out cross-validation. The transfer function was developed and the cladoceran-based temperature reconstructions were run with the program C2 data analysis (Juggins, 2007).
In addition to the temperature transfer function, a previously available Cladocera-based transfer function of water depth (Nevalainen et al., 2011) was applied to each of the intralake surface sediment samples. The multilake water depth calibration model was constructed from 55 samples across Finland. It has a water depth gradient of 0.5–7.0 m making it also suitable for this study lake, although the deepest sample in Lake Pieni-Kauro (7.3 m) was slightly deeper (0.3 m) than the deepest sample in the multilake training set. The water depth refers to the depth at the sampling sites (~sublittoral) rather than maximum lake depth. The modern analogue technique (MAT) model has a coefficient of determination (r 2) of 0.58, root mean squared error of prediction (RMSEP) of 1.1 m and a maximum bias of 2.9 m. Lake Pieni-Kauro is not included in the depth or temperature training set data, thus not providing a bias into the reconstruction results.
Intralake dataset
A Limnos gravity corer was used also to collect the 34 surface sediment samples along a water depth gradient between 0.5 and 7.3 m from Lake Pieni-Kauro and the Saavanjoki River. The sampling was performed from a small boat, M/S Unski, in October 2008. The samples were selected to characterise different habitat types, with a special focus on hydrological gradients (stream flow and water depth). Fossil Cladocera analysis was performed using standard methods described by Szeroczyńska & Sarmaja-Korjonen (2007) (see details in Nevalainen, 2011, this issues).
For the intralake samples, Pearson correlations (r) and coefficients of determination (r 2) were calculated and the levels of statistical significance (at P ≤ 0.05) were determined between measured water depth, Cladocera-inferred water depth and Cladocera-inferred mean air T Jul to detect the influence of sampling site selection (effects of stream flow and water depth) in downcore studies. Species diversity along the intralake water depth gradient was assessed using the Shannon index (Shannon, 1948), H′ = Σ(P i ln P i ). The null model software EcoSim (Gotelli & Entsminger, 2011) was used to apply the individual-based rarefaction approach on the incidence data for diversity calculations. In the rarefaction, the number of observations was set to 100 for each sample. The relationship between water depth and H′ was assessed using the Pearson correlation coefficient, coefficient of determination and level of statistical significance (P ≤ 0.05). A locally weighted scatterplot smooth (lowess) with a span of 0.7 was applied to the intralake data to illustrate general trends.
Results
Temperature training set
The DCA results for the temperature training set showed gradient lengths of 2.625 (axis 1) and 1.542 (axis 2), and eigenvalues λ1 = 0.267 and λ2 = 0.108, respectively, suggesting unimodal response of the taxa and further analysis based on CCA. The CCA showed that Cladocera assemblages were strongly correlated with mean air T Jul, because the relationship between the first and the second eigenvalue (λ1:λ2) was relatively high (0.670) and significant (P = 0.001). Compared to other climatic parameters, the mean air T Jul had stronger relationship with the distribution and abundance of Cladocera taxa (Table 1). The DCA indicated two outlier lakes (Rusutjärvi in Tuusula and Ylisjärvi in Muurla), which are hypereutrophic (total phosphorus > 100 μg l−1) lakes and were thus removed from the calibration model to reduce extraneous noise. The best Cladocera-mean air T Jul inference model (Fig. 2a) was developed with the WA-PLS technique using two regression calibration components and had favourable performance statistics: an r 2jack of 0.67, RMSEP of 0.86°C and mean and maximum biases of −0.02 and 1.73°C, respectively (Table 2). The addition of the second WA-PLS component decreased the model’s RMSEP by 15.6%, whereas a third component decreased the RMSEP by only 0.8%. There was a slight but statistically significant (P < 0.001) trend in the sample residuals, as cold lakes inferred generally overestimated temperatures and warm lakes underestimated temperatures (Fig. 2b).

Cladocera-based mean July air temperature transfer function using 76 lakes in Finland and its performance statistics, a the relationship between observed and inferred mean July air temperature values and b residuals along the temperature gradient
The Cladocera-water depth reconstruction model by Nevalainen et al. (2011) uses 55 lakes across Finland. The performance statistics of this MAT model are tabulated in Table 2.
Intralake dataset and temperature and water depth inferences
Of the 34 samples from Lake Pieni-Kauro and the Saavanjoki River, 31 contained enough remains for fossil Cladocera analysis. 41 Cladocera taxa were identified, mainly chydorids. Bosmina (Eubosmina) Seligo was the most abundant planktonic taxon and Alona affinis (Leydig) and Alonella nana Baird were the most abundant chydorids. The species assemblages along the intralake depth gradient are presented in detail elsewhere (Nevalainen, 2011).
The inferred mean air T Jul for the surface samples varied between 13.7 and 15.7°C. No statistically significant (P ≤ 0.05) systematic trend was found between the measured water depth and inferred air temperature (Table 3). However, the samples taken from depths >3 m showed consistently underestimated temperatures (Fig. 3a). Only one of the four lotic samples showed underestimation that was more than the calibration model’s RMSEP. None of the samples showed overestimation that was higher than the model’s RMSEP and 74% of the total samples had values within the model’s RMSEP and when only the lentic samples were included the share was 76.7%. The maximum bias in the inferred values of the samples was at 2.9 m, where the underestimation was 1.5°C.

Relationship between measured water depth and a Cladocera-inferred mean air T Jul, b Cladocera-inferred water depth and c rarefied Shannon diversity in the 31 surface sediment samples in Lake Pieni-Kauro and the Saavanjoki River, eastern Finland. Stream samples are marked as grey dots, lake samples as black dots and the lake-specific average sample as a sum (Σ). A locally weighted scatterplot smooth (lowess) (0.7) is applied to the data to illustrate general trends. The error bars in Fig. 3a and b represent the models’ root mean squared errors of prediction (RMSEP) and the 95% confidence intervals in Fig. 3c
The inferred water depth for the surface samples varied between 1.7 and 4.4 m. In overall, the inferred values were consistent with the measured values having a strong relationship (r = 0.67, r 2 = 0.45, P < 0.001) (Table 3). However, the lentic samples taken from the shallowest depths (~1 m) and three of the four lotic samples inferred overestimated depths (Fig. 3b). Furthermore, samples taken from depths >5 m inferred homogenously underestimated values (ca., 4 m). In all, 61.3% of the samples had inferred values within the model’s RMSEP and when only the lentic samples were included it increased to 66.7%. The maximum bias (−3.2 m) was found from a lotic sample, which was also the shallowest sample in the dataset.
A significant relationship was found between the measured water depth and rarefied H′ (r = 0.56, r 2 = 0.32, P < 0.001) (Table 3), the diversity indices varying between 1.5 and 2.5. The highest diversity was found from samples of shallow water and lowest diversity from samples taken from depths >4 m (Fig. 3c). All the stream samples had relatively high species diversity.
Discussion
Temperature training set
The CCA results showed that mean air T Jul was a major factor influencing Cladocera distribution in shallow lakes along the latitudinal transect in Finland (Table 1). However, limnological data was available only for some of the sites and, therefore, the examination of the influence of other parameters than those related to climate, was not possible for the present dataset. If it would have been possible to include limnological variables as co-variables in the analyses, the variance explained by the mean air T Jul could have potentially decreased. Temperature influences cladocerans directly through metabolism and reproduction (Frey, 1982; Gillooly & Dodson, 2000; de Eyto & Irvine, 2001), but also indirectly as temperature dictates major part of limnological variability. Therefore, at a regional scale climate may have profound influence on their distribution, whereas at local and site-specific scales other factors can have more important influence on the community composition. For example, limnology, hydrology, wind exposure, presence of macrophytes, and fish predation (Sweetman & Smol, 2006; DeSellas et al., 2008; Chen et al., 2010; Davidson et al., 2010; Nevalainen & Luoto, 2010) have a major contribution in structuring cladoceran communities. This implies that cladocerans can be used as ecological indicators of several environmental variables at different spatial scales. However, in palaeolimnology, it may be difficult or even impossible to separate many of the environmental signals from downcore data as these variables tend to simultaneously change through time. Furthermore, many environmental variables have interconnections; lake trophic status and macrophyte abundance have often a strong correlation with latitude/temperature and substrate type and fish predation and presence of macrophytes correlate with water depth, providing problems for palaeoenvironmental reconstructions (e.g. Bjerring et al., 2009; Nevalainen, 2011). Although it is evidently important to carefully examine the influence of all these parameters on Cladocera-based reconstructions, in this study, we focus on the influence of site-specific hydrology (stream flow and water depth) on quantitative temperature and water depth reconstructions.
The developed Cladocera-based mean air T Jul inference model showed relatively favourable performance statistics with an r 2jack of 0.67, an RMSEP of 0.86°C (Fig. 2a) and a maximum bias of 1.73°C (Fig. 2b). These statistics are well competitive with the previous Cladocera-based temperature inference models from the Alps (Lotter et al., 1997), northwestern Finnish Lapland (Korhola, 1999) and Scotland (Kattel et al., 2008), although they are not directly comparable due to their differences in the number of training set lakes and temperature gradient. The trend in the sample residuals where cold lakes inferred slightly overestimated temperatures and warm lakes underestimated temperatures (Fig. 2b), i.e. the linear distortions at the ends of environmental gradients, are a common and inherent feature in most WA-based reconstruction models (Luoto, 2010b). In the current temperature model, the problem is focused on the cold end of the temperature gradient, where the samples’ temperature distances are more uneven. Thus, the problem could be reduced in the future by adding more samples to the cold end of the gradient.
Inferred temperature and water depth for the intralake samples
The application of the temperature inference model to the site-specific samples from Lake Pieni-Kauro revealed no significant relationship between the inferred mean air T Jul and measured water depth (Table 3). However, a clear trend where the inferred temperatures were slightly underestimated was apparent, but this underestimation occurred mostly within the model’s RMSEP (77% of the lacustrine samples) (Fig. 3a). A more serious underestimation when taking core samples from the deepest point of a lake can occur when using midges (Nematocera, mostly Chironomidae) as a quantitative temperature proxy, because most of the cold-indicating taxa live in the profundal whilst the warm-indicators are often found from the shallow areas (Luoto, 2010a). Furthermore, the profundal environment and macrobenthic community is protected from direct influence of air temperature changes as the profundal water temperatures in stratified/dystrophic lakes remain persistently at ~4°C (the Hertta database, Finnish Environment Institute). Of the stream samples, only one was outside the model’s RMSEP showing underestimation of >1°C, whereas all the other samples showed inferred temperatures, which were very close to the measured value (Fig. 3a). Similar results have been found using fossil midges: a stream contribution does not have significant influence on midge-based temperature reconstructions (Engels et al., 2008; Luoto, 2010a). The inferred temperature in the summary sample, combining all the cladoceran data from Lake Pieni-Kauro, was also very close to the measured temperature, hence indicating that the model works well in the study site. Interestingly, the largest biases were at some of the samples taken from depths between 2 and 3 m. Nevertheless, also at this depth range most of the samples showed reliable temperature estimates.
In addition to temperature (e.g. Lotter et al., 1997; Korhola, 1999; Bigler et al., 2006; Kamenik et al., 2007), Cladocera are also strongly influenced by water depth enabling their use as a quantitative proxy for lake level changes (Korhola et al., 2005; Nevalainen et al., 2011). The influence of depth on Cladocera is mostly indirect and mediated through, e.g. chemical and temperature stratifications, presence of macrophytes, substrate type and food availability (Brodersen et al., 1998; Korhola et al., 2000; Jeppesen et al., 2003). Nevertheless, despite the indirect nature of the influence, water depth reconstructions using Cladocera assemblages are well established and validated (Korhola et al., 2005; Luoto, 2009b; Nevalainen et al., 2011). When the multilake water depth calibration model was applied on the Pieni-Kauro data, a strong and significant relationship between the inferred and measured values was found (Table 3). However, the samples taken from depths >5 m inferred underestimated values, which all bended to a level of 4 m (Fig. 3b). This is most likely because majority of the taxa found from the surface sediment samples from lake Pieni-Kauro were littoral chydorids (Chydoridae), whose fossil remains apparently accumulate close to their original shallow habitats (Nevalainen, 2011) and the planktonic species, such as Bosmina (Eubosmina) and Daphnia can occur in open-water over both the deeper and shallower parts of the lake. However, it should be noted that the deepest site in Lake Pieni-Kauro is probably the most underestimated because it is 0.3 m deeper than the deepest lake in the multilake dataset and therefore, the model cannot reliably estimate its water depth. Furthermore, the distinct trend in the residuals of the water depth inference model (Nevalainen et al., 2011), where shallowest sites were overestimated and deepest sites underestimated, are potential causal factors for the similar trend found in Fig. 3b. The intermediate depths and sites close to the training set’s mean depth of 2.6 m hence provide the most reliable water depth inferences.
Three of the four riverine samples showed inferred values that were overestimated by several meters and only one sample had the estimation close to the measured water depth (Fig. 3b). This implies that a stream contribution can have an effect on Cladocera-based water depth reconstructions, especially because the fossil remains originated from streams (usually shallow sites) can more easily re-accumulate into the end basin in lakes with a significant inlet, following events of elevated turbulence. The summary sample of all intralake species data had inferred water depth very close to the measured depth indicating that the overall assemblage represents well the average water depth of the lake.
Intralake variability in diversity and implications for optimal sampling point
The diversity of the cladocerans, measured as H’, in Lake Pieni-Kauro had significant correlation with the measured water depth. The diversity was higher in the shallow and intermediate samples and lower in samples taken from depths >4 m. This is due to the fact that there were only few planktonic taxa in Lake Pieni-Kauro and also the number of benthic chydorids found from the deeper waters was low, while most of the chydorid taxa prefer littoral environments (Nevalainen, 2011). All the lotic samples had relatively high cladoceran diversity, corresponding to the level found from the lentic samples taken from the approximately same depth.
The results from the examinations of the influence of water depth on cladocera-based reconstructions of air temperature and water depth and on species diversity indicate that samples taken from the deepest part of the lake show underestimated inferred values and lower cladoceran diversity. The most reliable quantitative estimates were found from intermediate depths of ca., 2–4 m, where also the diversity was high. However, the tendency in the model performance of shallow sites being overestimated and deep water sites being underestimated can cause the present results favour the intermediate depths. Although the present results suggest that it could be recommendable to take downcore sediment samples from intermediate depths, as they seem to provide the most reliable quantitative Cladocera-based estimations, the results should be considered carefully because of the problems in the water depth inference model’s performance (trends in residuals). In addition, the deepest sample in Lake Pieni-Kauro was probably underestimated because it was slightly outside the calibration model’s depth gradient. However, regardless of the problems in the water depth model, intermediate samples seem to have higher diversity compared to deep water sites providing also a higher number of indicator taxa and thus, increasing the reliability of reconstructions.
Although the temperature reconstructions were apparently not influenced by stream flow, much higher water depths were inferred from the stream samples compared to the measured values. This gives a reason to avoid major inlets when taking sediment cores to study lake level changes, and it is also likely that lake level fluctuations can be detected more easily from enclosed basins rather than open basins (Mason et al., 1994). The present results are in good agreement with previous Cladocera-based results from Scotland (Kattel et al., 2007) and midge-based results from Finland (Luoto, 2010a) that also indicated the optimal sampling point to locate at intermediate depths. Therefore, most reliable multiproxy results from invertebrate-based quantitative palaeoenvironmental reconstructions are probably achieved when taking the samples from the sublittoral parts of the study lakes and the deepest points are avoided. However, when taking core samples from the sublittoral, lake morphometric features, such as steepness, should be taken into consideration to avoid areas of potential sediment mixing or hiatuses. It is also important to recognise that the sedimentary patterns vary among lakes, as e.g. steep lakes can have more homogenous mixture of fossil remains (van Hardenbroeck et al., 2011) and in multiproxy studies light weighted microfossils, such as pollen and diatoms, can more easily be transported into deeper locations (Mieszczankin, 1997; Kauppila, 2002).
Conclusions
The performance statistics of the Cladocera-based inference model of summer air temperature were relatively favourable suggesting that it is valid in means of performing temperature inferences from fossil material. The results from the intralake and -river dataset from Lake Pieni-Kauro and the Saavanjoki River provide evidence for the influence of hydrological variables of water depth and stream flow on Cladocera-based palaeolimnological reconstructions of air temperature and water depth. The samples taken from shallow and intermediate water depths showed good correspondence between the inferred and measured temperature and depth, but the samples taken from depths >4 m had inferred values lower than the measured temperature and depth. However, this trend may have been driven by the apparent problems in the water depth inference model’s residuals. Nevertheless, the cladoceran diversity was lower in the deep samples, thus decreasing the number of many potential indicator taxa. The stream samples had high diversity and generally correct quantitative estimates for summer air temperature, but the Cladocera-inferred water depths were greatly overestimated. These results imply that the optimal sampling point for quantitative Cladocera-based temperature and lake level reconstructions could locate at intermediate water depths and stream contribution should be avoided when performing lake level reconstructions. Although it is clear that more research is needed to validate these results in a geographically larger context, the present results imply that when reconstructing temperatures using fossil Cladocera, a change in lake level can cause a serious bias to the temperature reconstruction. Our results also suggest that intralake datasets of fossil remains may be useful in validating multilake water depth calibration sets through inferring the natural within-lake depth gradient.
References
Alhonen, P., 1970a. On the significance of the planktonic/littoral ratio in the cladoceran stratigraphy of lake sediments. Commentationes Biologicae 35: 1–9.
Alhonen, P., 1970b. The paleolimnology of four lakes in south-western Finland. Academiae Scientiarum Fennicae Series A III Geologica-Geographica 105: 1–39.
Bigler, C., O. Heiri, R. Krskova, A. F. Lotter & M. Sturm, 2006. Distribution of diatoms, chironomids and cladocera in surface sediments of thirty mountain lakes in south-eastern Switzerland. Aquatic Sciences 68: 154–171.
Bjerring, R., E. Becares, S. Declerck, E. M. Gross, L. A. Hansson, T. Kairesalo, M. Nykänen, A. Halkiewicz, R. Kornijów, J. M. Conde-Porcuna, M. Seferlis, T. Nõges, B. Moss, S. L. Amsinck, B. V. Odgaard & E. Jeppesen, 2009. Subfossil Cladocera in relation to contemporary environmental variables in 54 Pan-European lakes. Freshwater Biology 54: 2401–2417.
Bos, D. G., B. F. Cumming & J. P. Smol, 1999. Cladocera and Anostraca from the interior plateau of British Columbia, Canada, as paleolimnological indicators of salinity and lake level. Hydrobiologia 392: 129–141.
Brodersen, K. P., M. C. Whiteside & C. Lindegaard, 1998. Reconstruction of trophic state in Danish lakes using subfossil chydorid (Cladocera) assemblages. Canadian Journal of Fisheries and Aquatic Sciences 55: 1093–1103.
Chen, G., C. Dalton & D. Taylor, 2010. Cladocera as indicators of trophic state in Irish lakes. Journal of Paleolimnology 44: 465–481.
Davidson, T. A., C. D. Sayer, M. Perrow, M. Bramm & E. Jeppesen, 2010. The simultaneous inference of zooplanktivorous fish and macrophyte density from sub-fossil cladoceran assemblages: a multivariate regression tree approach. Freshwater Biology 55: 546–564.
de Eyto, E. & K. Irvine, 2001. The response of three chydorid species to temperature, pH and food. Hydrobiologia 459: 165–172.
DeSellas, A. M., A. M. Paterson, J. N. Sweetman & J. P. Smol, 2008. Cladocera assemblages from the surface sediments of south-central Ontario (Canada) lakes and their relationships to measured environmental variables. Hydrobiologia 600: 105–119.
Engels, S., S. J. P. Bohncke, O. Heiri & M. Nyman, 2008. Intraregional variability in chironomid-inferred temperature estimates and the influence of river inundations on lacustrine chironomid assemblages. Journal of Paleolimnology 40: 129–142.
Frey, D. G., 1960. The ecological significance of cladoceran remains in lake sediments. Ecology 41: 684–699.
Frey, D. G., 1982. Contrasting strategies of gamogenesis in northern and southern populations of Cladocera. Ecology 63: 223–241.
Frey, D. G., 1986. Cladocera analysis. In Berglund, B. E. (ed.), Handbook of Holocene Palaeoecology and Palaeohydrology. John Wiley and Sons, New York: 667–692.
Gillooly, J. F. & S. I. Dodson, 2000. Latitudinal patterns in the size distribution and seasonal dynamics of new world, freshwater cladocerans. Limnology and Oceanography 45: 22–30.
Gotelli, N. J. & G. L. Entsminger, 2011. EcoSim: Null Models Software for Ecology. Version 7. Acquired Intelligence Inc. & Kesey-Bear, Jericho.
Hann, B. J., 1989. Methods in quaternary ecology. Cladocera. Geoscience Canada 16: 17–26.
Hofmann, W., 1998. Cladocerans and chironomids as indicators of lake level changes in north temperate lakes. Journal of Paleolimnology 19: 55–62.
Jeppesen, E., P. Leavitt, L. De Meester & J. P. Jensen, 2001. Functional ecology and palaeolimnology: using cladoceran remains to reconstruct anthropogenic impact. Trends in Ecology & Evolution 16: 191–198.
Jeppesen, E., J. P. Jensen, T. L. Lauridsen, S. L. Amsinck, K. Christoffersen, M. Søndergaard & S. Mitchell, 2003. Sub-fossils of cladocerans in the surface sediment of 135 lakes as proxies for community structure of zooplankton, fish abundance and lake temperature. Hydrobiologia 491: 321–330.
Juggins, S., 2007. Program C2 Data Analysis. Version 1.5.0. University of Newcastle, Newcastle Upon Tyne.
Kamenik, C., K. Szeroczyńska & R. Schmidt, 2007. Relationships among recent Alpine Cladocera remains and their environment: implications for climate-change studies. Hydrobiologia 594: 33–46.
Kansanen, P., T. Jaakkola, S. Kulmala & R. Suutarinen, 1991. Sedimentation and distribution of gamma-emitting radionuclides in bottom sediments of southern Lake Päijänne, Finland, after the Chernobyl accident. Hydrobiologia 222: 121–140.
Kattel, G. R., R. W. Battarbee, A. W. Mackay & H. J. B. Birks, 2007. Are cladoceran fossils in lake sediment samples a biased reflection of the communities from which they are derived? Journal of Paleolimnology 38: 157–181.
Kattel, G. R., R. W. Battarbee, A. W. Mackay & H. J. B. Birks, 2008. Recent ecological change in a remote Scottish mountain loch: an evaluation of a Cladocera-based temperature transfer-function. Palaeogeography, Palaeoclimatology, Palaeoecology 259: 51–76.
Kauppila, T., 2002. Variability of surface sediment diatom assemblages in Lake Salkolanjärvi, Finland. In Proceedings of the 15th International Diatom Symposium, Perth, 1998: 263–274.
Korhola, A., 1999. Distribution patterns of Cladocera in subarctic Fennoscandian lakes and their potential in environmental reconstruction. Ecography 22: 357–373.
Korhola, A. & M. Rautio, 2001. Cladocera and other Branchiopod crustaceans. In Smol, J. P., H. J. B. Birks & W. M. Last (eds), Tracking Environmental Change Using Lake Sediments, Volume 4: Zoological Indicators. Kluwer Academic Publishers, Dordrecht: 5–41.
Korhola, A., H. Olander & T. Blom, 2000. Cladoceran and chironomid assemblages as quantitative indicators of water depth in subarctic Fennoscandian lakes. Journal of Paleolimnology 24: 43–53.
Korhola, A., M. Tikkanen & J. Weckström, 2005. Quantification of Holocene lake-level changes in Finnish Lapland using a cladocera-lake depth transfer model. Journal of Paleolimnology 34: 175–190.
Kurek, J. & L. C. Cwynar, 2009. The potential of site-specific and local chironomid-based inference models for reconstructing past lake levels. Journal of Paleolimnology 42: 37–50.
Laird, K. R., M. V. Kingsbury & B. F. Cumming, 2010. Diatom habitats, species diversity and water-depth inference models across surface-sediment transects in Worth Lake, northwest Ontario, Canada. Journal of Paleolimnology 44: 1009–1024.
Lotter, A. F., H. J. B. Birks, W. Hofmann & A. Marchetto, 1997. Modern diatom, cladocera, chironomid, and chrysophyte cyst assemblages as quantitative indicators for the reconstruction of past environmental conditions in the Alps. I. Climate. Journal of Paleolimnology 18: 395–420.
Lotter, A. F., H. J. B. Birks, U. Eicher, W. Hofmann, J. Schwander & L. Wick, 2000. Younger Dryas and Allerød summer temperatures at Gerzensee (Switzerland) inferred from fossil pollen and cladoceran assemblages. Palaeogeography, Palaeoclimatology, Palaeoecology 159: 349–361.
Luoto, T. P., 2009a. Subfossil Chironomidae (Insecta: Diptera) along a latitudinal gradient in Finland: development of a new temperature inference model. Journal of Quaternary Science 24: 150–158.
Luoto, T. P., 2009b. A Finnish chironomid- and chaoborid-based inference model for reconstructing past lake levels. Quaternary Science Reviews 28: 1481–1489.
Luoto, T. P., 2010a. Hydrological change in lakes inferred from midge assemblages through use of an intralake calibration set. Ecological Monographs 80: 303–329.
Luoto, T. P., 2010b. Spatial and Temporal Variability in Midge (Nematocera) Assemblages in Shallow Finnish Lakes (60–70°N): community-based modelling of past environmental changePublications of the Department of Geosciences and Geography, University of Helsinki A4: 1–62.
Mason, I. M., M. A. J. Guzkowska, C. G. Rapley & F. A. Street-Perrot, 1994. The response of lake levels and areas to climatic change. Climatic Change 27: 161–197.
Mieszczankin, T., 1997. A spatio-temporal pattern of pollen sedimentation in a dimictic lake with laminated sediments. Water, Air, & Soil Pollution 99: 587–592.
Mikkola, P., 2008. Koillis-Kainuun kallioperä. Summary: Pre-Quaternary Rocks of Northeast Kainuu. Geological Survey of Finland, Report of Investigation 175.
Nevalainen, L., 2011. Intralake heterogeneity of sedimentary cladoceran (Crustacea) assemblages forced by local hydrology. Hydrobiologia. doi:10.1007/s10750-011-0707-3.
Nevalainen, L. & T. P. Luoto, 2010. Implications for the use of sedimentary invertebrate communities to infer past presence of fish. Knowledge and Management of Aquatic Ecosystems 396(5): 1–13.
Nevalainen, L., K. Sarmaja-Korjonen & T. P. Luoto, 2011. Sedimentary Cladocera as indicators of past water level changes in shallow northern lakes. Quaternary Research 75: 430–437.
Nykänen, M., K. Vakkilainen, M. Liukkonen & T. Kairesalo, 2009. Cladoceran remains in lake sediments: a comparison between plankton counts and sediment records. Journal of Paleolimnology 42: 551–570.
Sarmaja-Korjonen, K., 2001. Correlation of fluctuations in cladoceran planktonic:littoral ratio between three cores from a small lake in southern Finland: Holocene water-level changes. The Holocene 11: 53–63.
Shannon, C. E., 1948. A mathematical theory of communication. Bell System Technical Journal 27(379–423): 623–656.
Sweetman, J. & J. P. Smol, 2006. Patterns in the distribution of cladocerans (Crustacea, Branchiopoda) in lakes across a north-south transect in Alaska, USA. Hydrobiologia 553: 277–291.
Szeroczyńska, K. & K. Sarmaja-Korjonen, 2007. Atlas of Subfossil Cladocera from Central and Northern Europe. Friends of the Lower Vistula Society, Świecie.
ter Braak, C. J. F. & P. Šmilauer, 2002. CANOCO Reference Manual and Canodraw for Windows User’s Guide: Software for Canonical Community Ordination (version 4.5). Microcomputer Power, Ithaca.
van Hardenbroeck, M., O. Heiri, M. F. Wilhelm & A. F. Lotter, 2011. How representative are subfossil assemblages of Chironomidae and common benthic invertebrates for the living fauna of Lake De Waay, The Netherlands? Aquatic Sciences 73: 247–259.
Walker, I. R., C. H. Fernando & C. G. Paterson, 1984. The chironomid fauna of four shallow lakes and their representation by subfossil assemblages in the surficial sediments. Hydrobiologia 112: 61–67.
Acknowledgments
This study is a contribution of the EPHIPPIUM (Academy of Finland, grant no. 1107062), INFER (INtralake Fossil invERtebrate Assemblages) and LOSER (LOng-term climate impactS on lakE tRophic status) projects. Personal financial support was provided by the Kone Foundation for T.P. Luoto and Waldemar von Frenckell Foundation for L. Nevalainen. We thank the two anonymous reviewers for their helpful comments that improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: H. Eggermont & K. Martens / Cladocera as indicators of environmental change
Rights and permissions
About this article
Cite this article
Luoto, T.P., Nevalainen, L., Kultti, S. et al. An evaluation of the influence of water depth and river inflow on quantitative Cladocera-based temperature and lake level inferences in a shallow boreal lake. Hydrobiologia 676, 143–154 (2011). https://doi.org/10.1007/s10750-011-0801-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-011-0801-6