
Abstract
Sources of nitrogen and phosphorus were measured for a small pool, Brown Moss, which is a Site of Special Scientific Interest in the UK. The site is designated for its macrophyte and bird interest and comprises several pools flanked by heath and woodland. Currently the largest pool has high mean concentrations of total N and P (4 ± 2.4 mg N l−1 and 389 ± 254 μg P l−1), and also suffers from water table fluctuation and recreational impact. There are no point sources of nutrients but potential diffuse sources include direct rainfall, drainage from surrounding heathland and potentially from distal agricultural land, some septic tank drainage and bird excreta. Nutrients are also released from the sediments. Amongst external sources of phosphorus, birds contributed 234 mg m−2 yr−1 (73%), land drainage 63 mg m−2 yr−1 (20%), and rainfall 24 mg m−2 yr−1 (7%). Major bird species were dabbling ducks and geese. For external sources of nitrogen, the most important was land drainage (2,860 mg m−2 yr−1 and 64%), followed by rainfall (870 mg m−2 yr−1 and 19%) and birds (741 mg m−2 yr−1 and 11%) There was substantial internal loading of phosphorus (1670 mg m−2 yr−1) so that internal loading dominated the overall phosphorus budget (84%). The nitrogen budget was dominated by land drainage, though for short periods, geese made a major contribution. Restoration of the site to the status required by European legislation will be very difficult, if not impossible in the medium-term.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Birds are important components of the conservation psyche, familiar and affectionately regarded by both the general public and amateur ornithologists, and sometimes highly damaging to the sites where they are conserved. Several instances exist where guanotrophication, the avian version of eutrophication, is a problem (Leentvaar, 1967; Brandvold et al., 1976; Moss & Leah, 1982; Bales et al., 1993; Pettigrew et al., 1998; Don & Donovan, 2002; Vanni, 2002; Hahn et al., 2007), and the problems caused by exotic species such as Canada geese (Branta canadensis (Linnaeus)) and ruddy duck (Oxyura jamaicensis (Gmelin)) are widely acknowledged (Hussong et al., 1979; Conover & Chasko, 1985). Less well recognised but potentially of great significance are the problems of switching of shallow lakes from macrophyte-dominated communities to phytoplankton-dominated systems by bird flocks held well above natural carrying capacity by artificial feeding by the public (Moss et al., 1996). Almost every municipal park in the UK provides an example. Birds may nonetheless be crucial natural contributors of nutrients to lakes (Leentvaar, 1967; Portnoy, 1990 Manny et al., 1994). In that case there may be conflicts, sometimes not acknowledged or realised, between aspirations for conservation management of plants and birds. Brown Moss, in Shropshire, UK, may be one such case.
Brown Moss is a Site of Special Scientific Interest and part of a Ramsar site with several small and shallow pools. The largest pool has plenty of aquatic birds, some of which are resident and often fed by the public, for it is a popular and accessible site; some are exotic species, and others are migrants. The site, however, is susceptible to other pressures, including diffuse nutrient loading from surrounding land and septic tanks, water level fluctuation and atmospheric pollution. It no longer has the plant diversity that it formerly had (Whild, 2007). Management plans have depended more on casual information than systematic investigation. The present study attempted a comprehensive nutrient budget for the site as a contribution to an evidence-based management approach.
Materials and methods
Study site
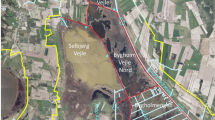
Brown Moss is small (33 ha) located to the south-east of Whitchurch, in northern Shropshire, UK (Latitude: 52o57′; Longitude: 2o39′; National Grid Reference SJ 562395) (Fig. 1). It was notified as a Site of Special Scientific Interest (SSSI) in 1953 and is owned and managed by Shropshire County Council. It is also a Local Nature Reserve, Special Area of Conservation, and part of the Midlands Meres and Mosses Ramsar Site. This study focussed on the largest pool on the site, known as Pool 6.

Map of Brown Moss SSSI. The studied pool (6) is indicated by an arrow
The pool (Fig. 1) has an area of 2.9 ha and mean depth <1 m. It is isolated, supplied by shallow ground water and rain and there are no surface inflow streams. The water is highly eutrophic, with mean concentrations of TP, TN and chlorophyll a (n = 49) of 389 ± 254 μg l−1, 4.0 ± 2.4 mg l−1 and 99 ± 96 μg l−1, respectively, in 2005–2007. Large numbers of birds congregate on the pool in preference to other smaller ones nearby, which do not provide adequate flight paths for water birds owing to surrounding trees.
Methods
Samples of rain water were collected on a regular basis by using a plastic rainfall collector (40 × 57 × 19 cm), situated above-ground level, outside the Bioscience Building, School of Biological Science, the University of Liverpool from January to December 2006. Rain collected in Liverpool has a broadly similar nutrient concentration to that elsewhere in northern England and the west midlands. Estimation of nutrient loading from rain was based on average annual rainfall, average nutrient concentrations and area of the pool (Migon & Sandroni, 1999).
Shallow soil water and groundwater flowing from the land was collected by ten tubes ((lysimetres) of 10 cm diameter × 1.50 m deep) installed in two lines N/S and E/W across the area of Brown Moss SSSI in January 2005. Water chemistry of groundwater was analysed every two weeks from February 2005 to October 2006. Values of conservative ions (alkalinity, conductivity and winter TN) of groundwater from boreholes were statistically compared (similarity analysis) with those of the pool to determine those closest to the pool chemistry and to generate the likely catchment area. Estimation of nutrient loading by shallow ground water to the pool was then calculated based on volume of run-off water entering the pool from that area, mean concentration of plant nutrients from the specific boreholes concerned and area of catchment estimated. The average annual rainfall and evaporation at Brown Moss were 0.70 and 0.57 m yr−1, respectively (Environmental Consultancy, 2001). The amount of water entering the pool was estimated from the difference between average annual precipitation and evaporation, which is 0.13 m yr−1.
Nutrient release from the sediment was measured once a month from November 2004 to October 2006. Fifteen coring tubes with three replicates (length 50 cm diameter 6.9 cm) were inserted into the pool bed to a depth of approximately 15 cm. Cores composed of pool water (70%) above the sediment (30%) were then sealed bottom and top with rubber bungs and kept in darkness and brought to the laboratory. Approximately 200 ml of pool water from each tube was immediately pumped after being brought to the laboratory from about 5 cm above the sediment surface through a plastic tube without stirring to avoid re-suspension of the sediment and the water filtered and analysed. Afterwards, core samples without further treatment were kept in darkness and incubated for 24 h in a water bath located in an open area outside the building at ambient temperature, which mimicked the pool environment. Redox conditions are thus likely to reflect those found in the pool. After 24 h, 200 ml of overlying water was again pumped from the same cores, filtered and analysed for nutrients. The volume of water left in the samples was finally measured and net nutrient release rates were estimated based on the difference between concentrations of nutrients measured at the start compared with the concentrations after 24 h.
To estimate nutrient loading from waterfowl, birds present near and in the pool were counted around mid-day by species every two weeks using binoculars (10 × 50 wide angle) from November 2004 to October 2006. Diurnal estimates carried out every two months showed that the mid-day values were representative of the 24-h period. The number of bird-days was calculated for each species. Bird excretion rates were based on literature estimates and a log–log relationship with estimated weight (Gould and Fletcher, 1978; Manny et al., 1994). For instance, the daily nutrient load by migrant Canada geese was on average 1,570 mg TN and 490 mg TP per individual per day. Monthly nutrient load from birds to the pool was estimated using nutrient loading rates (TP and TN), monthly numbers of specific waterfowl and area of the pool. Most of the bird biomass fed on the land rather than in the water, especially in winter and spring. Supplementary addition of food by people also tended to encourage land feeding. Birds were thus assumed to be bringing largely external nutrients into the pool rather than recycling existing nutrients.
Water samples from the main pool were taken fortnightly from November 2004 to October 2006. Water collected from the pool, rain water, soil and shallow ground water and sediment release cores was filtered through Whatman GFC filters. The filtered water was subsequently analysed for soluble reactive phosphorus (SRP, μg l−1), nitrate nitrogen (NO3 −-N, mg l−1) and ammonium nitrogen (NH4 +-N, μg l−1), based on Mackereth et al. (1989). Total nitrogen (TN, mg l−1) and total phosphorus (TP, μg l−1) were measured following Johnes & Heathwaite (1992) on unfiltered water. pH and conductivity (μS cm−1) were measured with a Hanna HI 9812 meter and alkalinity (mEq l−1) by titration against standard acid to an end point of pH 4.5 (Mackereth et al., 1989) within a few hours after collection.
Values given are means ± standard deviation and numbers of samples measured. Regression was used to establish relationships amongst variables in the pool. Tukey honestly significant difference (HSD) tests were used to investigate differences amongst seasons. Bray–Curtis analysis, (PRIMER 6.0) was used to analyse similarities amongst alkalinity, conductivity and total nitrogen data for groundwater and the pool.
Results
In rain water, phosphorus was less abundant than nitrogen. Average concentrations of SRP, TP, NO3 −-N, NH4 +-N and TN (n = 19) in rain water were 18.9 ± 12 μg l−1, 33.8 ± 22.8 μg l−1, 0.33 ± 0.2 mg l−1, 768 ± 475 μg l−1 and 1.2 ± 0.7 mg l−1, respectively. The amounts of both TP and TN were highest in summer and lowest in winter (Fig. 2). Overall, no statistical differences between average concentrations of TP and TN in each season were observed (F = 1.69, P = 0.211 and F = 0.53, P = 0.669, respectively). Estimated TP and TN contributed by rain water was calculated using average annual rainfall (0.7 m yr−1) multiplied by average TP and TN of rain water. Annual TP and TN loadings to the main pool by rainfall were 24 and 870 mg m−2 yr−1, respectively.

Average concentrations of TP and TN delivered from rain water to Pool 6 at Brown Moss in each season
Soil and shallow ground water levels fluctuated dramatically in the boreholes. The lowest and highest levels recorded were 0 and 72 cm (measured from the bottom of the borehole to the groundwater surface) in September 2005 and in May 2006, respectively. Ground water levels in the boreholes were positively correlated with that in the main pool. (r = 0.529, P = 0.016).
Concentrations of nutrients in boreholes increased simultaneously with those in the main pool. Nitrate-N in particular increased mostly in winter and autumn and was low in spring and summer (Fig. 3). In contrast, lack of washout in spring and summer was associated with lower concentrations of nitrate. Higher concentrations of total nitrogen and phosphorus were detected in boreholes situated along the edge of Brown Moss close to farmland and cottages (Fig. 4). In contrast, groundwater in boreholes in the middle of the site had lower concentrations of nutrients.

Average seasonal concentrations of TP and TN in boreholes

Average concentrations of TP and TN in boreholes around the site (black and white columns represent boreholes near the edge of the site and in inner areas, respectively)
Bray–Curtis analysis using values of alkalinity and conductivity, and concentrations of winter TN (Fig. 5) which are assumed to be conservative, suggested that the main pool receives most water from areas where boreholes 1, 2, 3, 4 and 8 were situated (Fig. 6). The areas where boreholes 5, 6, 7, 9 and 10 were located were not included because the results showed that they were less similar in conservative ions than those mentioned. The likely catchment area of the pool around boreholes 1, 2, 3, 4 and 8 was calculated as 12 ha.

Cluster analysis of alkalinity and conductivity (above), and winter TN (below) amongst boreholes and the pool

The estimated catchment area of the main pool (dot line) and alkalinity (A) and conductivity (C) of groundwater in the boreholes measured around Brown Moss
Total phosphorus and TP input from groundwater was estimated from average concentrations of TP (117 mg m−3) and TN (5,320 mg m−3) from the appropriate boreholes, multiplied by runoff (0.13 m yr−1) and estimated catchment area of the pool (120,000 m2), then divided by area of the pool. Estimated TP and TN loadings from ground and soil water were 63 and 2,860 mg m−2 yr−1, respectively.
Nutrient release rates varied throughout the season owing to climatic conditions and water fluctuations (Fig. 7). SRP, TP and NH4 +-N were mostly released from the sediments in summer. Winter release of SRP, TP and NH4 +-N was much lower. Release of TN from the sediment began to increase in late winter and early spring but release of NO3 −-N was negligible or transformed to other N forms in all seasons. Average release rates of TP and TN (n = 3) were 4.57 ± 8.5 and 0.96 ± 30.8 mg m−2 day−1, respectively. Annual TP and TN release rates were 1,670 and 350 mg m−2 yr−1, respectively.

Nutrient release (TP and TN (mg m−2 month−1)) from the sediment over 24 h measured from 2004 to 2006 (values above the zero line represent mean release to the water and values below the zero line represent mean uptake by the sediment)
Thirteen species of waterfowl were observed in the pool during the study period and categorised into six groups, based on body weight. Most birds were dabbling ducks whilest scarcest were grebes (Fig. 8). Proportions of coot, small ducks, geese and gulls were similar. A large flock of Canada geese was observed in winter 2004. Numbers were relatively lower in 2005 and 2006.

Seasonal numbers of birds present on the main pool at Brown Moss from November 2004 to October 2006
More species and higher numbers of wildfowl were recorded in winter and numbers decreased gradually towards summer (Fig. 9). Densities of waterfowl reached a peak (108 ha−1 day−1) in November 2004 when large numbers of Canada geese were roosting on the pool and fell to 6 ha−1 day−1 in September 2005 when water in the main pool nearly dried out. Daily loading rates from birds are shown in Table 1.

Species of waterfowl recorded in each season
Monthly mean TP and TN loadings by aquatic birds were 0.57 and 1.8 kg per pool, respectively. Higher TN and TP loads were found in winter and autumn and lower values in spring and summer (Fig. 10). Annual TP and TN loadings by waterfowl to the pool were 234 and 741 mg m−2 yr−1, respectively. The impact of birds was clearly seen in winter when a gathering of Canada geese in 2004/2005 immediately increased nitrogen compounds in the pool as indicated by strong correlation between bird abundance and ammonium concentrations in the water (r = 0.871, P = 0.01).

Seasonal loading of nutrients by waterfowl to the main pool, Brown Moss
Birds contributed the most external phosphorus (73%) to the pool (Table 2) whereas groundwater and rainwater added 20 and 7% of TP, respectively. Groundwater was the major external major source of nitrogen (64%) whilst rain water and birds supplied 19 and 17% of nitrogen to the pool, respectively.
Of total external plus internal loading, sediment was the major supplier of phosphorus and ground water contributed the most nitrogen to the pool. Phosphorus release from the sediment contributed up to 84% and ground water supplied about 60% of nitrogen (Table 2). Birds, groundwater and rain added 12, 3 and 1% of phosphorus. Rain, birds and sediment contributed 18, 15 and 7% of nitrogen.
Discussion
Construction of a reasonably accurate and precise nutrient budget is conceptually easy but is rarely done because it is an expensive task for even moderate sized catchments. The amount of data needed is considerable. Usually budgets are desk-based using models. These can be precise if there are already data collected in the field available to calibrate and validate them (Johnes et al., 1996) but increasingly that is not the case and the models may be becoming less precise and even less accurate as some sources of nutrients may be overlooked. Small catchments provide insights, because they can be relatively exhaustively studied, that may prove helpful in delineating the scope of models for larger ones. The relative importance of wild birds may be one such aspect.
Bird flocks can be major contributors of nutrients and clearly so where their numbers are spectacular. The hundreds of thousands of gulls roosting in the late 1970s at Hickling Broad in Norfolk (Moss & Leah, 1982) whitened the surface of the water as they flew in at dusk (B. Moss & R. T. Leah, personal communication); the excretal contribution made by cormorants to the wetland fringes of the Nardermeer in the Netherlands can be smelled as well as seen and the nutrient export from the reserves of the Wildfowl and Wetlands Trust in the UK, where geese, swans and ducks are attracted by suitable management and some artificial feeding is great enough to merit treatment systems. What is not so obvious is the contribution made by less spectacular communities of birds and Brown Moss is a case in point.
Apart from a very occasional visit of a view days by a small flock of Canada geese, when immediate effects on the ammonia concentration in the water were recorded, the bird community of Brown Moss is not particularly notable. It does not include particularly unusual birds and the common ones are not unusually abundant. Nonetheless, birds dominated the external phosphorus budget, even though there are cottages around the site discharging septic tank effluent into it, as manifested by the high phosphorus concentrations in some of the soil waters. The total load of 321 mg m−2 yr−1 from all external sources would create a theoretical concentration of (321 × 29,000)/(0.13 × 120,000) or about 597 μg P l−1, compared with the mean concentration recorded of 389 μg P l−1 and so theoretically could create the recorded concentration without any contribution from sediment release. Sediment release, however, as in many shallow lakes (Jensen & Andersen, 1992; Petterson, 1998; Søndergaard et al., 1999, 2003) was very considerable, amounting to 1,670 mg m−2 yr−1. Of itself this could create a far higher concentration ((1,670 × 29,000)/(0.13 × 120,000) or 3,104 μg P l−1) than recorded. However, if there are processes of seepage into the pool, there are likely to be seepage losses from the pool to the deeper groundwater. The total theoretical concentration of TP is 597 + 3,104 = 3,711 μg P l−1 implying that only about 389/3,711 or 10% of the load is retained within the system. The phosphorus in the sediment must originally come from external sources so there must be an element of double-counting, but the concentration recorded is nonetheless very high, possibly due to anthropogenic impacts, compared with the approximately 15 μg P l−1 to be expected from a pristine catchment under these climatic and geological conditions. Brown Moss thus appears to have a very severe eutrophication problem if the evaluation is made from phosphorus alone and birds must be accounted major contributors to this, perhaps because their numbers are boosted by artificial feeding by the public. In many other instances birds may be simply part of the nutrient budget but not problematic (Kitchell et al., 1999).
Some of the lakes in the north-west midland plain of the UK, however, are frequently very phosphorus-rich (of the order of 1 mg TP l−1). This is attributed to concentration mechanisms leading to phosphorus-saturated sediments in groundwater-fed lakes with long retention times (Kilinc & Moss, 2002) or to the presence of rich apatite deposits in the glacial drift that covers the plain (Reynolds, 1979). There are instances also of eutrophication by intensive stock farming and sewage effluent in the area (Moss et al., 1994) but these sources may be absent yet very high TP concentrations (of the order of mg P l−1) found (Moss et al., 1997). In such lakes, bioassays suggest that the phytoplankton is nitrogen limited (Hameed et al., 1999; James et al., 2003). Brown Moss is not such a lake and might be expected to have much more conventional phosphorus concentrations. Nonetheless, the ratio of nitrogen to phosphorus in the external load to Brown Moss was 2.47, whereas the mean ratio in the water was 10.3. The former value is well below the conventional Redfield ratio and implies a severe relative shortage of nitrogen to the pool at Brown Moss. The ratio in the water still implies this but the nitrogen has apparently been to some extent enriched, perhaps through nitrogen fixation by prokaryotes in the sediment because planktonic cyanobacterial blooms have not been recorded.
Two-thirds of the nitrogen comes to the pool from soil seepage, the remainder from direct rainfall and birds. Sediment release is a very minor contributor. The catchment is not agricultural but comprises birch woodland and heathland, though there is the possibility of some contribution from septic tank drainage. Run-off from agricultural land normally has very high nitrogen to phosphorus ratios as nitrogen compounds are much more soluble than phosphorus compounds and nitrogen fertiliser is widely used (Young & Briggs, 2005). The marginal boreholes receiving water from agricultural land at the edge of Brown Moss had N to P ratios around 400 and this is likely due to stores of nitrogen in the cultivated soils. Run-off from pristine forest vegetation under similar climatic and geological conditions to those at Brown Moss has a TN to TP ratio of about 10:1 (Smith et al., 2003) and meta-analyses (Elser et al., 2007) of nutrient bioassay experiments have shown that limitation by either nitrogen alone or nitrogen and phosphorus together is much more common than previously believed (Schindler, 1977, 1978).
Nutrient budgets give valuable information for management of nature reserves. The pool water at Brown Moss is clearly artificially eutrophicated and changes in the flora according to a study of Whild (2007) suggest that even the terrestrial vegetation has changed through increased nitrogen loading since the 19th century as atmospheric deposition of nitrogen has increased and agricultural nitrogen has entered at the edges. The pool, even without a fish community, due to drying out in summer 2005, to predate on zooplankton and increase algal biomass through a trophic cascade sometimes has high chlorophyll a concentrations. The TP concentrations are well above those proposed under European legislation for restoration of water bodies to good ecological status (UKTAG, 2007) and this means that a restoration plan must be put into place. The problem is a very difficult one, however. It is essential to reduce external loads but reduction in bird numbers would be illegal under UK wildlife legislation. Discouragement of artificial feeding by the public has been attempted but has not solved the problem and in any case it may be that occasional visits by larger groups of Canada geese are greater contributors than the smaller ducks and gulls that are the main recipients of bread thrown into the water. Canada geese are introduced species that are expanding their numbers in the UK. They can be legally shot, but the more effective control comes from pricking of eggs at nests, a procedure made difficult by the widespread and covert nesting sites. Little can be done about nutrient stores in the soil bar waiting for the passage of time to leach out the stores. The effluents from septic tanks are small contributors to the problem and it would be costly to install mains sewerage.
Because there is substantial release of phosphorus from the sediment an obvious procedure might be sediment sealing or removal. There are several objections to this, however. It is disruptive and expensive and may have little effect unless external loads are first reduced. In any case, nitrogen, for which sediment is a minor contributor, may be the more important nutrient in controlling algal growth in the pool. Symptoms such as toxic blooms are not present in any case.
The value of the site is as a bird reserve and as a site for the rare macrophyte, Luronium natans (L.) Raf., a submerged plant that is an annual and easily displaced by more vigorous vegetation. There is a strong relationship between nitrogen loading and submerged plant diversity (James et al., 2003; Barker et al., 2008) and the increased nitrogen loading at Brown Moss has undoubtedly contributed to Luronium reduction (Whild, 2007) in recent decades. The fluctuating water levels, exposing bare sediment undoubtedly favour such a species but at Brown Moss they also favour two introduced competitors, Crassula helmsii (Kirk) Cockayne and Epilobium brunescens (Cockayne) Raven & Engelhorn that might be much more important in suppressing Luronium than the increased nitrogen loadings. The nitrogen that comes from land seepage probably has not increased greatly during the period that Luronium has declined because the site was formerly grazed and dunged. Most of the increase has thus been from the atmosphere and attributable to agricultural volatilisation of ammonia and nitrate from oil burning in vehicle engines. This is not a problem that can be solved on a site-by-site basis. We conclude that the situation is currently intractable. The ultimate problem is partly of eutrophication, but eutrophication exacerbated by the problems of introduced bird and plant species that cannot be easily controlled.
References
Bales, M., B. Moss, G. Phillips, K. Irvine & J. Stansfield, 1993. The changing ecosystem of a shallow, brackish lake, Hickling Broad, Norfolk, UK. II Long-term trends in water chemistry and ecology and their implications for restoration of the lake. Freshwater Biology 29: 141–165.
Barker, T., K. Hatton, M. O’Connor, L. Connor & B. Moss, 2008. Effects of nitrate load on submerged plant biomass and species richness: results of a mesocosm experiment. Fundamental and Applied Limnology 173: 89–100.
Brandvold, D. K., C. J. Popp & J. A. Brierley, 1976. Waterfowl refuge effect on water quality: chemical and physical parameters. Journal of Water Pollution Control Federation 48: 685–687.
Conover, M. R. & G. G. Chasko, 1985. Nuisance of Canada geese problems in the eastern United States. Wildlife Society Bulletin 13: 228–233.
Don, G. L. & W. F. Donovan, 2002. First Order Estimation of the Nutrient and Bacterial Input from Aquatic Birds to Twelve Rotorua Lakes. Bioresearches, Auckland: 58.
Elser, J. J., M. E. S. Bracken, E. E. Cleland, D. S. Gruner, W. S. Harpole, H. Hillebrand, J. T. Ngai, E. W. Seabloom, J. B. Shurin & J. E. Smith, 2007. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters 10: 1135–1142.
Environmental Consultancy, 2001. Meres and Mosses Conservation Plans, Brown Moss. University of Sheffield, Sheffield.
Fredrickson, L. H., 1969. Mortality of coots during severe spring weather. Wilson Bulletin 81(4): 450–458.
Gould, D. J. & M. R. Fletcher, 1978. Gull dropping and their effects on water quality. Water Research 12: 665–672.
Hahn, S., S. Bauer & M. Klaassen, 2007. Estimating the contribution of carnivorous waterbirds to nutrient loading in freshwater habitats. Freshwater Biology 52(12): 2421–2433.
Hameed, H. A., S. Kilinc, S. McGowan & B. Moss, 1999. Physiological tests and bioassays–aids or superfluities to the diagnosis of phytoplankton nutrient limitation? A comparative study in the Broads and Meres of England. European Phycological Journal 34: 253–270.
Hussong, D., J. M. Damaré, R. J. Limpert, W. J. Sladen, R. M. Weiner & R. R. Colwell, 1979. Microbial impact of Canada geese (Branta canadensis) and whistling swans (Cygnus columbianus columbianus) on aquatic ecosystems. Applied Environmental Microbiology 37(1): 14–20.
James, C., J. Fisher & B. Moss, 2003. Nitrogen driven lakes: the Shropshire and Cheshire Meres? Archiv fur Hydrobiologie 2003: 249–266.
Jensen, H. S. & F. O. Andersen, 1992. Importance of temperature, nitrate and pH for phosphorus release from aerobic sediments of four shallow eutrophic lakes. Limnology and Oceanography 73(3): 557–589.
Johnes, P., B. Moss & G. Phillips, 1996. The determination of total nitrogen and total phosphorus concentrations in freshwaters from land use, stock headage and population data: testing of a model for use in conservation and water quality management. Freshwater Biology 36: 451–473.
Johnes, P. J. & A. L. Heathwaite, 1992. A procedure for the simultaneous determination of total nitrogen and total phosphorus in freshwater samples using persulphate microwave digestion. Water Research 26: 1281–1287.
Kilinc, S. & B. Moss, 2002. Whitemere, a lake that defies some conventions about nutrients. Freshwater Biology 47: 207–218.
Kitchell, J. F., D. E. Schindler, B. R. Herwig, D. M. Post, M. H. Olson & M. Oldham, 1999. Nutrient cycling at the landscape scale: the role of diel foraging migrations by geese at the Bosque del Apache National Wildlife Refuge, New Mexico. Limnology and Oceanography 44: 828–836.
Leentvaar, P., 1967. Observations in guanotrophic environments. Hydrobiologia 29: 441–489.
Mackereth, F. J. H., J. Heron & J. F. Talling, 1989. Water Analysis: Some Revised Methods for Limnologists. Freshwater Biological Association, Scientific Publication. No. 36: 120.
Manny, B. A., W. C. Johnson & R. G. Wetzel, 1994. Nutrient additions by waterfowl to lakes and reservoirs: predicting their effects on productivity and water quality. Hydrobiologia 279/280: 133–147.
Migon, C. & V. Sandroni, 1999. Phosphorus in rainwater: partitioning inputs and impact on the surface coastal ocean. Limnology and Oceanography 44(4): 1160–1165.
Moss, B. & R. T. Leah, 1982. Changes in the ecosystem of a guanotrophic and brackish shallow lake in eastern England: potential problems in its restoration. International Review Gesamten Hydrobiology 67: 625–659.
Moss, B., S. McGowan & L. Carvalho, 1994. Determination of phytoplankton crops by top-down and bottom-up mechanisms in a group of English lakes, the West Midland meres. Limnology and Oceanography 39: 1020–1029.
Moss, B., J. Madgwick & G. Phillips, 1996. A guide to the restoration of nutrient- enriched shallow lakes. Environment Agency & Broads Authority, Norwich.
Moss, B., M. Beklioglu, L. Carvalho, S. Kilinc, S. McGowan & D. Stephen, 1997. Vertically-challenged limnology; contrasts between deep and shallow lakes. Hydrobiologia 342/343: 257–267.
Petterson, K., 1998. Mechanisms for internal loading of phosphorus in lakes. Hydrobiologia 373(374): 21–25.
Pettigrew, C. T., B. J. Hann & L. G. Goldsborough, 1998. Waterfowl feces as a source of nutrients to a prairie wetland: responses of microinvertebrates to experimental additions. Hydrobiologia 362: 55–66.
Portnoy, J. W., 1990. Gull contribution of phosphorus and nitrogen to a Cape Cod kettle pond. Hydrobiologia 202: 61–69.
Reynolds, C. S., 1979. The Limnology of the Eutrophic Meres of the Shropshire – Cheshire Plain – A Review. Field Studies Council, Shropshire.
Schindler, D. W., 1977. Evolution of phosphorus limitation in lakes. Science 195: 260–262.
Schindler, D. W., 1978. Factors regulating phytoplankton production and standing crop in the world’s freshwaters. Limnology and Oceanography 23: 478–486.
Smith, R. A., R. B. Alexander & G. E. Schwarz, 2003. Natural background concentrations of nutrients in streams and rivers of the conterminous United States. Environmental Science and Technology 37: 3039–3047.
Søndergaard, M., J. P. Jensen & E. Jeppesen, 1999. Internal phosphorus loading in shallow Danish lakes. Hydrobiologia 408/409: 145–152.
Søndergaard, M., J. P. Jensen & E. Jeppesen, 2003. Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 506–509: 135–145.
UKTAG, 2007. Recommendations on surface water classification schemes for the purposes of the water framework directive. Environment Agency, Bristol.
Vanni, M. J., 2002. Nutrient cycling by animals in freshwater ecosystems. Annual Review of Ecology and Systematics 33: 341–370.
Young, E. O. & R. D. Briggs, 2005. Shallow ground water nitrate-N and ammonium-N in cropland and riparian buffers. Agriculture Ecosystems and Environment 109: 297–309.
Whild, S., 2007. Detecting Changes in the Aquatic Vegetation of the West Midlands Meres of Great Britain. PhD thesis, The University of Birmingham, Birmingham.
Acknowledgement
RC thanks the Royal Thai Government for a scholarship to work at the School of Biological Sciences, University of Liverpool.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: M. Meerhoff, M. Beklioglu, R. Burks, F. García-Rodríguez, N. Mazzeo & B. Moss / Structure and Function of World Shallow Lakes: Proceedings from the 6th Shallow Lakes Congress, held in Punta del Este, Uruguay, 23–28 November, 2008
Rights and permissions
About this article
Cite this article
Chaichana, R., Leah, R. & Moss, B. Birds as eutrophicating agents: a nutrient budget for a small lake in a protected area. Hydrobiologia 646, 111–121 (2010). https://doi.org/10.1007/s10750-010-0166-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-010-0166-2