
Abstract
The Upper River Paraná Floodplain System comprises the rivers Ivinheima, Baía and Paraná, which with their associated waterbodies form three subsystems, each showing individual characteristics. Hydrilla verticillata recently invaded the Upper Paraná Floodplain, while Egeria najas is the native most abundant submersed macrophyte. A large flood-pulse, during January–March 2007, abruptly reduced macrophyte stands in many areas to near-zero and dispersed propagules over the entire floodplain. From April 2007 to April 2008, we conducted three surveys sampling for the presence–absence of H. verticillata and E. najas and environmental variables aiming to answer: (1) How rapid is the colonization–regeneration process for both species? (2) Which habitats seem to be more susceptible to their colonization? (3) Which environmental factors can best predict their occurrence? Neither H. verticillata nor E. najas colonized the Ivinheima subsystem. In the Baía subsystem, E. najas had only two occurrences while H. verticillata was not present. In the Paraná subsystem, E. najas predominantly occurred in river channels, but it was also common in floodplain lakes. In April 2007, it was found in 13% of the sites in the Paraná subsystem, increasing to 30% in November 2007 and reaching 34% in April 2008. H. verticillata did not successfully colonize floodplain lakes of the Paraná subsystem. In channels, it had 34% occurrence in April 2007, increasing to 62% in November 2007 and remaining at 62% in April 2008. The role of environmental variables in predicting species occurrence changed depending upon the scale of the analysis. Considering the whole Upper Paraná floodplain, water transparency followed by electrical conductivity were the strongest predictors for both species. Colonization by submersed plants seems improbable in the Ivinheima subsystem owing to its low water transparency besides frequent localized floods; in the Baía subsystem, it seems inhibited by transparency and low alkalinity. Considering just the Paraná subsystem, the proportion of organic matter in sediment, ten times higher in floodplain lakes than in channels, was the best predictor for H. verticillata occurrence (also related to water pH and transparency), while E. najas was only significantly explained by transparency.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Hydrilla verticillata is regarded as one of the worst aquatic weeds in the world (Murphy, 1988; Mullin et al., 2000; Hershner & Havens, 2008). The weed shows a large ecological amplitude and thrives under environmental conditions with characteristics varying from lentic to lotic, from oligotrophic to eutrophic, from acid to alkaline, from low to high light availability and from tropical to temperate climate (Van et al., 1976; Barko & Smart, 1981; Cook & Lüönd, 1982; Spencer et al., 1994; White et al., 1996; Kahara & Vermaat, 2003). H. verticillata has specialized structures for vegetative reproduction (tubers and turions) besides a well-developed rhizome system capable of stocking carbohydrate reserves which enable the plant to persist and recover even after severe disturbances (Netherland, 1997; Owens & Madsen, 1998; Madsen & Smith, 1999). These features, together with aggressive growth rates and its morphological aspects, forming dense canopies near the water surface, make H. verticillata a highly competitive weed which can potentially disrupt native macrophyte communities (Haller & Sutton, 1975; Van et al., 1999; Mony et al., 2007; Wang et al., 2008).
Hydrilla verticillata recently invaded the Upper River Paraná Floodplain System (URPFS), a strategic ecosystem for biodiversity conservation in Brazil. Here, the weed was first recorded in June 2005. Since then it has rapidly spread along the River Paraná infesting large areas with high biomass production and causing problems related to navigation, fishing and recreation activities. The URPFS comprises a rich diversity of organisms such as fish and invertebrates, as well aquatic macrophytes (Agostinho et al., 2004) which can also be severely impacted by such an aggressive invader (Agostinho et al., 2005). Egeria najas is the dominant native submersed macrophyte in the URPFS (Thomaz et al., 2004a). E. najas and H. verticillata are both Hydrocharitaceae with closely similar morphology and ecological features which can result in intense competitive interactions between them. Although E. najas is regarded as a potential nuisance species that can cause some localized problems in artificial water bodies (Bini & Thomaz, 2005), it is not a nuisance weed outside its native range of southern Brazil, north east Argentina and Paraguay (Cook & Urmi-König, 1984; Chambers et al., 2008).
While H. verticillata is one of the most extensively studied aquatic macrophytes, there is only scarce information about the biology and ecology of E. najas (e.g. Tavechio & Thomaz, 2003; Pierini & Thomaz, 2004; Thomaz et al., 2007b), especially in its native habitats (Bini & Thomaz, 2005; Thomaz et al., 2006). Information about which factors can predict colonization of these native and exotic macrophytes can be an important tool for development of conservation and management efforts (Murphy & Pieterse, 1990; Wingfield et al., 2006).
From January to March 2007, a strong flood-pulse disturbed populations of H. verticillata and E. najas in the URPFS reducing stand biomass to almost zero in many areas (W. T. Z. Sousa, unpublished data). Indeed, only propagules of the plants (stem and root fragments) were found immediately after the big flood. The flood also acted to disperse large quantities of propagules between waterbodies in the floodplain as a result of the increased connectivity between them during the flood event (e.g. Neiff, 1990; Thomaz et al., 2007a). The flood provided an opportunity to study submersed macrophyte recolonization after a major disturbance event particularly aiming to address three questions: (1) How fast over time and how intense over space does the colonization–regeneration process occur in invasive H. verticillata and native E. najas post-disturbance in the URPFS?; (2) Which habitats seem to be more susceptible or protected from the occupation of these exotic and native Hydrocharitaceae?; (3) Which environmental factors are likely to hinder or propitiate the successful establishment of each species?
Study area
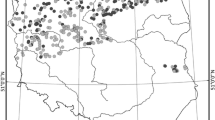
The study area comprises the three main rivers of the URPFS: Ivinheima, Baía and Paraná (Fig. 1). Each river with its associated waterbodies has its own individual characteristics (see Thomaz et al., 2004b) and can be regarded as a distinct subsystem of the URPFS: hereafter coded as the Ivinheima subsystem (IS), Baía subsystem (BS) and Paraná subsystem (PS).

Map of the sampling stations in the URPFS
The hydrology of the River Paraná is strongly influenced by an upstream cascade of reservoirs which retains solids and nutrients, resulting in clear oligotrophic water in the river channel by the time it reaches the URPFS. The water flow is influenced by the volume of precipitation in the catchment area and by operation of upstream dams (Thomaz et al., 2004b). Periods of high waters usually occur during the summer (November–March). When the water level surpasses 3.5 m above local datum (at Porto Rico: 22°46′ S; 53°15′ W) a majority of the URPFS, including waterbodies of the three subsystems, become highly connected (Souza-Filho et al., 2004). The flood enriches the river water with suspended solids and nutrients brought from the inundated floodplain resulting in reduction of water transparency. Supplied by groundwater, the River Baía is a semi-lotic distributary, rich in humic compounds (Souza-Filho et al., 2004; Thomaz et al., 2004b). Its hydrology, including direction of flow, is largely influenced by the water level regime of the River Paraná (Thomaz et al., 2004a). The River Ivinheima shows the lowest water transparency [depth of disappearance of the Secchi disc (ZDS) <1.4 m] and its hydrology is more independent, with local floods occurring with relative higher frequency and intensity, depending on precipitation in its (un-dammed) sub catchment (Souza-Filho et al., 2004; Thomaz et al., 2004a).
Methods
Data collection
The first survey was made in April 2007 at 148 sampling sites (c. 100 m2 each) distributed along 35 waterbodies (up to 38 sites per waterbody, dependent on size of the waterbody) located in the three subsystems of the URPFS (Fig. 1; Table 1). In each site, depths lower than 4 m were sampled with a rake for presence–absence data of H. verticillata and E. najas. Where these species were not colonizing, the presence of vegetative propagules was also checked in each site. Site coordinates were marked with a GPS which enabled precisely the same sites to be sampled again in November 2007 and April 2008. However, in April 2008, one channel in the BS (with 10 sampling stations) could not be assessed by boat due to vegetation blockage.
In April and November 2007, physico-chemical variables were measured at 38 sampling stations (Table 1). In April 2007, we analysed water pH, electrical conductivity, alkalinity and depth of disappearance of the Secchi disc (ZDS). In November 2007, we measured these variables plus underwater light attenuation coefficient (k), concentrations of phosphorus (TP) and nitrogen (TN), and sediment proportion of organic matter (O.M.sed). In April 2008, ZDS and electrical conductivity were sampled in all the sites sampled for macrophytes (n = 138).
Water electrical conductivity and pH were measured with a portable meter (Digimed, São Paulo, Brazil). In order to calculate the light attenuation coefficient (k; m−1), we measured the photosynthetically active radiation (PAR; μmol s−1 m−2) immediately subsurface and at a known depth using a LiCor underwater quantum sensor connected to a hand-held meter. k was estimated following the equation k = [ln (I 0) − ln (I d )]/d (where, I 0 = irradiance at the subsurface, I d = irradiance at the depth d m). Water samples were taken for determination of alkalinity using Gran titration (Carmouze, 1994) and for laboratory analysis of concentrations of total phosphorus (TP) and total nitrogen (TN) according to Golterman et al. (1978) and Zagatto et al. (1981), respectively. Sediment samples were collected for analysis of proportion of organic matter per dry mass (O.M.sed; % DM−1). This was obtained by gravimetry after burning ca. 0.3 g of sediment in a furnace at 450°C for 4 h.
Data analyses
Analysis of variance (ANOVA) followed by Tukey post-hoc mean-separation testing was applied to search for statistical differences in environmental conditions between subsystems. Pair-wise t-tests were used to find differences between lentic and lotic habitats within a subsystem. Dependent variables were checked for assumptions of normality and homogeneity of variances.
Relationships between macrophytes and environmental data were assessed by logistic regression analysis. The procedure was as described by Bini & Thomaz (2005) plus references therein. For each species, the best combination of predictor variables was chosen using the following procedure: (1) all the predictors were tested separately; (2) the one that resulted in the largest change of deviance given the degrees of freedom (assessed by the chi-square (χ2) statistic) was added to the model; (3) the procedure was repeated with all the remaining predictors until addition of another variable did not result in a significant (at the P < 0.05 level) reduction in deviance (difference between χ2 statistics calculated for models with different number of predictors). Model predictive capability was evaluated by the percentage of cases in which observed presence, and absence data were misclassified. An arbitrary logit value (0.5) was used as a threshold to categorize misclassification. A significance level of P < 0.05 was considered significant for all the analyses. The software Statistica 7 for Windows (StatSoft Inc.) was used.
Results
Macrophyte occurrences
Hydrilla verticillata was not found at any site in the IS during any survey. In the BS, only propagules (plant fragments) were found just after the flood disturbance in April 2007 at only one sampling site. The macrophyte successfully established only in the river channels of the PS. Although in this subsystem propagules of H. verticillata were found in one floodplain lake in April 2007 and in five floodplain lakes in November 2007, H. verticillata was found colonizing only one floodplain lake station of the PS in each survey, and it was not found twice over time in the same lake. Considering only sampling sites located in river channels of the PS (n = 47 sites), H. verticillata occurred in 34.0% of the sites in April 2007 (just after the major flood disturbance) and increased to 61.7% of occurrence in November 2007. In April 2008, it had not enlarged its occupancy and still colonized 61.7% of the sites sampled in river channels of the PS.
Egeria najas also was not found in any site of the IS. In November 2007, this species occurred in one channel in the BS, which could not be sampled again in April 2008 because the boat access was blocked by vegetation development. However, in April 2008, E. najas was found once again in the BS, colonizing just one site in a floodplain lake. In the PS, E. najas occurred in 12.8% of the sites sampled in April 2007. Its occurrence increased to 29.8% in November 2007 and reached 34.0% of the sites in the PS in April 2008. In the PS, E. najas was found more frequently in river channel habitats (61% of the occurrences) and it was also common in floodplain lakes (39% of the occurrences).
Environmental conditions
Ivinheima (IS), Baía (BS) and Paraná (PS) subsystems
Environmental variables measured in the three subsystems of the URPFS during the period of study are summarized in Table 2. As expected, the major flood of January–March 2007 temporarily affected the limnological characteristics in the whole URPFS. A notable example of change was the homogenization of the water transparency among subsystems. Habitats of IS, BS and PS, which previously differed in water clarity, showed statistically similar values of ZDS in April 2007.
In November 2007 the limnological characteristics of each subsystem had returned to their normal range. Habitats of the PS had significantly higher water transparency (measured as ZDS and k) than both IS and BS which did not differ between each other (Table 2). Habitats of BS had significant lower electrical conductivity and alkalinity compared to IS and PS which did not differ between each other. The PS had significant lower concentrations of TN and TP while IS and BS did not differ in nutrients. In November 2007 the three subsystems of the URPFS did not differ statistically in regard to water pH and O.M.sed.
Analysis of the dataset from April 2008 for electrical conductivity and ZDS largely confirmed the results observed with data from November 2007 (Table 2). However, with the April 2008 dataset the analysis showed that the IS had significantly lower values of ZDS than the BS and PS, with the PS still sustaining the higher values.
River channels and floodplain lakes of the PS
In order to search for evidence that could explain why H. verticillata did not successfully establish in floodplain lakes but only in the river channels of the PS we compared the environmental conditions between these two types of habitat. The most pronounced (and significant; P < 0.001) difference was found for O.M.sed, 10 times higher in floodplain lakes (13.0 ± 3.1% DM−1) than in river channels (1.3 ± 1.1% DM−1). Other significant differences (P < 0.05) were detected for water pH, in April (7.3 ± 0.4 in the river and 6.8 ± 0.3 in lakes) and November 2007 (7.2 ± 0.7 in the river and 6.6 ± 0.2 in lakes), and for ZDS, in April 2008 (1.7 ± 0.4 m in the river and 1.1 ± 0.5 in lakes), both showing significantly lower values in floodplain lakes than in river channels. No other significant difference (P > 0.05) was found between channels and floodplain lakes of the PS for any environmental variable for any survey.
Predictors of species occurrence in the URPFS
Preliminary analyses (November 2007 dataset)
Given the atypical environmental and biotic conditions in the URPFS in April 2007 (associated with the major flood-pulse), we used data from November 2007 to explore preliminary relationships between macrophytes and environmental variables. This snapshot period was considered the most appropriate to choose because habitats were likely to have recovered from the unusual conditions caused by the major flood. In addition, by November 2007 propagules dispersed by the flood were considered likely to have had sufficient time to regenerate in habitats suitable for this to occur.
Light attenuation coefficient (k), ZDS and concentrations of TP and TN were the best predictors for both species (Table 3; Fig. 2). These four explanatory variables were strongly inter-correlated (r 2 > 0.31; P < 0.001). Together with the former ANOVA results, this suggests that habitats with lower water transparency (e.g. sites of IS and BS) had higher nutrient levels. H. verticillata and E. najas were positively related to water transparency and negatively related to nutrient levels. Since nutrients in excess were no longer likely to be directly limiting macrophyte development, water transparency was regarded as the main factor explaining species distribution over the URPFS. Thus, to avoid multicollinearity problems involving k, ZDS, TP and TN, only ZDS was included in multiple regression analysis. ZDS was preferred because of the ease in sampling it and its robustness (e.g. no need for equipment calibration).

Significant (P < 0.05) logistic regression models, yielded with the November 2007 dataset (n = 38), predicting probability of H. verticillata and E. najas occurrence in the URPFS as a function of k, ZDS, TP and TN. Statistics of each model are displayed in Table 3
Water electrical conductivity, alkalinity, pH and O.M.sed yielded significant relationships with H. verticillata occurrence, while pH was also a significant explanatory predictor of E. najas colonization (Table 3; Fig. 3). Electrical conductivity and alkalinity were strongly correlated (r = 0.79; P < 0.001), thus we also avoided coupling these variables in regression analysis. Between the two, electrical conductivity was preferred for its ease of measurement in field.

Logistic models, yielded with the November 2007 dataset (n = 38), predicting probability of macrophyte occurrence in the URPFS as a function of water electrical conductivity, alkalinity, pH and O.M.sed. For E. najas there were no significant effects of electrical conductivity (P = 0.079), alkalinity (P = 0.082) and O.M.sed (P = 0.995). All other relationships are significant (P < 0.05). Statistics of each model are displayed in Table 3
The relationship between ZDS and H. verticillata occurrence was significantly improved by including electrical conductivity (P = 0.03) in the model. Thus, with the November 2007 dataset, the best model (minimal adequate model) to predict probability of H. verticillata occurrence (P H. verticillata ) was obtained by combining ZDS and electrical conductivity (Table 3). The equation is as follows:
No other combination of explanatory variables significantly improved (P > 0.05) k or ZDS relationships with H. verticillata occurrence. With the November 2007 dataset, no variable significantly improved (P > 0.05) the relationship between E. najas occurrence and ZDS.
Main predictive models (all surveys dataset)
In April 2008, ZDS and water electrical conductivity were measured at all the sites sampled for macrophytes (n = 138). This substantially larger dataset allowed a refined evaluation of the role of these two variables in predicting species occurrence. With the use of the dataset from all the surveys (n = 214), ZDS and electrical conductivity each yielded highly significant models predicting probability of H. verticillata and E. najas colonization (Table 3). However, for both species, ZDS yielded models which achieved higher significance and predictability of the observed data than electrical conductivity. For both predictors H. verticillata had better models than E. najas.
Both species displayed quite similar positive relationships with their predictors (Fig. 4). However, probability of H. verticillata occurrence increases more steeply in relation to ZDS and electrical conductivity than is the case for E. najas. In fact, sites with H. verticillata did not differ significantly from sites with E. najas with regard to ZDS and electrical conductivity (Table 4), although sites with E. najas had lower minimum values (ZDS: 0.60 m; electrical conductivity: 28.0 μS cm−1) than the minimum observed for sites with H. verticillata (ZDS: 0.70 m; electrical conductivity: 46.1 μS cm−1).

Significant (P < 0.05) logistic models, yielded with all surveys dataset (n = 214), predicting probability of H. verticillata and E. najas occurrence in the URPFS as a function of ZDS and electrical conductivity. Statistics of each model are displayed in Table 3
Adding electrical conductivity significantly improved (P < 0.001) ZDS relationships with H. verticillata and E. najas yielding two best models with data from all the surveys (Table 3), as given below:
where P H. verticillata and P E. najas are probabilities of H. verticillata and E. najas occurrence, respectively.
Hydrilla verticillata had a better model than E. najas. The observed absence of both species was well predicted (more than 91% correct) by the models. However, sites with E. najas achieved low correct predictions (8.0%) while 64.4% of the sites with H. verticillata were correctly predicted by its model (Table 3).
Predictors of species occurrence in the PS
Since both species were commonest in the PS, we restricted our analysis to a search for environmental variables which could predict species occurrence considering only sites of the PS. With the November 2007 dataset (n = 16 sites, eight with H. verticillata and four with E. najas), no environmental variable showed any significant relationship with E. najas occurrence. However, for H. verticillata significant relationships were found with water pH, O.M.sed and k (Table 5; Fig. 5). None of these relationships could be significantly improved (P > 0.05) by the addition of any other variable. Water pH was strongly correlated with O.M.sed (r = −0.50; P < 0.05) and both were the best predictors for H. verticillata in the PS. Water pH and O.M.sed differed very little in correctly predicting sites without H. verticillata (seven and six sites, respectively). For sites with H. verticillata pH correctly predicted occurrence at five sites, while O.M.sed correctly predicted seven sites.

Probability of macrophyte occurrence in sites of the PS as a function of k, ZDS, pH and O.M.sed. For E. najas there were no significant effects of k (P = 0.505), pH (P = 0.100) and O.M.sed (P = 0.682). All other relationships were significant (P < 0.05). Models with k, pH and O.M.sed were yielded with data from November 2007 (n = 16), while models with ZDS were yielded with data from all the surveys (n = 94). Statistics of each model are displayed in Table 5
Despite the fact that ZDS did not show a significant relationship with H. verticillata (P = 0.147) or E. najas (P = 0.072) with the PS dataset from November 2007, the effect of water transparency upon the occurrence of both species in the PS was suggested by a significant relationship obtained with ZDS data from all the surveys (n = 94; Table 5; Fig. 5). Again, H. verticillata was much better predicted than E. najas. While correct predictions exceeded 67% for sites with and without H. verticillata, ZDS only correctly predicted 4.2% of the sites occupied by E. najas in the PS. With the PS dataset, electrical conductivity did not yield a significant relationship with either species (P > 0.13), nor did it significantly improve ZDS relationships with them (P > 0.41).
Discussion
Our results suggest that the absence of H. verticillata or E. najas is very predictable, whereas the presence of each taxon is less predictable. In particular, the presence of E. najas was only poorly predicted, at least in part due to the low number of sites with this species. Some environmental conditions (e.g. water transparency, electrical conductivity, sediment organic matter) could be related to the occurrence of these exotic and native submersed macrophytes, but the role of these variables in predicting species occurrence changed dependent upon the scale of the analysis.
Considering the URPFS as a whole, colonization by H. verticillata and E. najas was mainly explained by water transparency. H. verticillata did not colonize any habitat in the sub-catchments of River Ivinheima (IS) and River Baía (BS). E. najas also did not occur in the IS and only had rare appearances in the BS. Apart from the clear water state, the colonization of submersed macrophytes in the River Paraná is largely favoured by constant inputs of propagules from upstream reservoirs (which are densely colonized by submersed plants; Martins et al., 2008) and downstream dispersal via river water flow. However, major flood-pulses in the River Paraná tend to homogenize habitats of the URPFS, greatly increasing the horizontal connectivity (exchange of matter and energy) among waterbodies parallelling the main river channel (e.g. Neiff, 1990; Thomaz et al., 2007a). In other words, the flood acts to disperse macrophyte propagules over the floodplain system. Although in this study we did not quantify the flux of propagules to the sites sampled, H. verticillata propagules (plant fragments with adventitious roots) were seen in one station of the BS just after the major flood in April 2007, but this species was not found rooted in this habitat. Thus, propagules of both species were available at least in one site of the BS. Propagules of submersed plants are much more difficult to find during surveys than established stands of macrophytes. H. verticillata and E. najas both have a high dispersal capability via vegetative structures such as stem fragments (Cook & Lüönd, 1982; Cook & Urmi-König, 1984; Rybicki et al., 2001; Bini & Thomaz, 2005). Although macrophyte colonization in the River Paraná is favoured by longitudinal dispersal, it is very probable that propagules eventually become available in the sub-catchments of the River Ivinheima, and especially in the River Baía (S. M. Thomaz, unpublished). The proximity and hydrological connectivity between sub-catchments (especially during flood events) propitiate propagule movement between subsystems via processes such as flux of water, migration of animals such as waterfowl (Figuerola & Gree, 2002) and transit of fishing boats and fishing equipment. Thus, the availability of plant propagules alone does not seem to be the main cause of the unsuccessful establishment of species, especially in the BS.
Habitats of the BS had intermediate water transparency (ZDS: 0.91 ± 0.35 m, min: 0.30 m, max: 1.90 m) compared with the subsystems PS (ZDS: 1.57 ± 0.73 m, min: 0.50 m, max: 5.50 m) and IS (ZDS: 0.64 ± 0.19 m, min: 0.45 m, max: 1.30 m). Although the BS did not support dense stands of submersed macrophytes, as seen in the PS, E. najas and several other submersed species were found to colonize sites in the BS, such as Najas sp., Bulbostylis sp., Utricularia sp., Potamogeton sp., Bacopa sp. and Myriophyllum sp. (confirming the results of earlier studies which also found a rich submersed macrophyte flora at sites in this distributary, e.g. Murphy et al., 2003). Since H. verticillata invasion to the UPRFS is very recent (substantial infestations in the River Paraná were first detected in 2006) and this species is well adapted to cope with lower-transparency conditions (Van et al., 1976; Bowes et al., 1977; Barko & Smart, 1981; Spencer et al., 1994), it is possible that propagules of H. verticillata might settle in some shallow littoral areas of the BS with enough light availability to permit macrophyte development. For example, analysing the response curve yielded in this study (Fig. 4) the probability of H. verticillata occurrence increases abruptly when ZDS varies from 1.0 to 2.0 m. Also, while the minimum ZDS recorded in sites colonized by H. verticillata was 0.70 m, the maximum ZDS recorded in the BS was much higher, 1.9 m. Thus, in relation to light availability, our results suggest some possibility of eventual H. verticillata establishment in shallow sites of the BS. This suggestion is reinforced by studies conducted in the tidal Potomac River in the USA (Rybicki et al., 2001). There H. verticillata developed well in sites with values of ZDS fluctuating around 0.60 m, but not in localities with lower transparency (ZDS around 0.50 m) even though propagules were highly available. Rooted submersed species were never found colonizing any site in the IS (e.g. Thomaz et al., 2004a) and, owing to its low water transparency together with the high instability caused by frequent and intense localized flood disturbances, it is unlikely that submersed species will successfully colonize sites in the River Ivinheima sub-catchment if the current environmental conditions are maintained.
Water electrical conductivity helped ZDS in predicting submersed macrophyte colonization in the URPFS. This tendency was clearly influenced by the low electrical conductivity levels in the BS. The semi-lotic characteristics of the River Baía seems to propitiate the accumulation of organic matter from the surrounding wetlands and riparian vegetation (although not statistically significant, O.M.sed in the BS was higher than in PS and IS). Vegetation debris (e.g. rich in lignin and cellulose) tends to enrich the BS waters with dissolved organic compounds (e.g. humic substances) which tend to coagulate, adsorbing ions such as bicarbonates, and sink in the sediment (Wetzel, 1983). Indeed, water electrical conductivity and alkalinity were strongly correlated (r = 0.79; P < 0.001). H. verticillata and E. najas are both reportedly capable of utilizing bicarbonate as a source of dissolved inorganic carbon (Van et al., 1976; Kahara & Vermaat, 2003; Pierini & Thomaz, 2004). Thus, increasing water bicarbonate concentrations also increases the availability of dissolved inorganic carbon and tends to stimulate the photosynthesis and growth rates of some Hydrocharitaceae, including H. verticillata and E. najas (Van et al., 1976; Madsen & Sand-Jensen, 1987; Kahara & Vermaat, 2003; Pierini & Thomaz, 2004). Hence, the development of H. verticillata and E. najas may be inhibited in waterbodies of the BS, compared to other environments in the UPRFS, because of inorganic carbon scarcity. This agrees with the findings of Bini & Thomaz (2005) who found water electrical conductivity to be the main predictor of E. najas colonization in the Itaipu Reservoir, located downstream from the URPFS: they suggested that growth of E. najas may be favoured in waters with higher bicarbonate concentrations. Thus, the gradient of significant differences between the three subsystems of the URPFS for water electrical conductivity (BS: 43 ± 11 μS cm−1; IS: 58 ± 4 μS cm−1; PS: 64 ± 7 μS cm−1) can be regarded as a reasonable explanation which helps the differences in water transparency to predict the colonization–absence of these two Hydrocharitaceae at a regional scale (between the three sub-catchments), as explained by inorganic carbon availability to plants. From our results, it is reasonable to suggest that plants, especially H. verticillata, have a lower probability of occurrence in the IS and BS, compared to the PS, because of the lower water transparency and light availability, especially in the IS, coupled with lower inorganic carbon availability in the BS.
Hydrilla verticillata was strongly predominant in the River Paraná channels in relation to E. najas. Apart from its ability to produce adventitious roots which can grow from stem nodes, the exotic species has a well developed root system where stolons grow over the water–sediment interface as well as horizontally within the sediment, thus enabling the plant to expand rapidly over an area (Cook & Lüönd, 1982; Madsen & Smith, 1999). Moreover, carbohydrate allocation in the root system propitiates shoot regrowth through tubers, stolons and root crowns, after the photosynthetic tissues have been removed by disturbances (Netherland, 1997; Owens & Madsen, 1998). In contrast, E. najas has only simple slender adventitious roots (Cook & Urmi-König, 1984) and recovers after disturbance mainly from stem fragments. These advantages might have favoured the rapid recovery and expansion of H. verticillata in the River Paraná after the flood disturbance. Moreover, studies have shown that the rates of biomass increase of H. verticillata in the River Paraná are much higher than those shown by E. najas, which seems to suffer high competitive pressure from the alien species (W. T. Z. Sousa, unpublished data).
However, H. verticillata did not thrive in floodplain lakes connected to the River Paraná, where E. najas dominated. Owing to daily water level fluctuation of the River Paraná, due to upstream operation of dams for hydroelectric power generation, the exchange of water, and consequently of plant propagules, between the River Paraná and its connected floodplain lakes was intense. Propagules of H. verticillata (stem fragments) were very frequently seen inside the floodplain lakes, and thus lack of propagules was unlikely to be the cause for non-establishment of the weed in these habitats. Our results showed that in the PS H. verticillata colonization was related to water pH, sediment organic content and water transparency. Despite the fact that floodplain lakes had lower water transparency (ZDS: 1.20 ± 0.60 m, min: 0.50 m, max: 2.70 m) than sites in the River Paraná (ZDS: 1.73 ± 0.73 m, min: 0.50 m, max: 5.50 m), these floodplain lakes in the PS are generally shallow (max. depth: c. 2.5 m, outwith the flood period) with developed littoral zones which usually enables sufficient light for above-compensation point photosynthesis to reach the substrate. In consequence, development of abundant stands of submersed macrophytes such as E. najas and Cabomba furcata is common in the PS floodplain lakes. Both H. verticillata and E. najas are able to photosynthesise at low light availabilities, overcoming light scarcity by forming canopies near the water surface (Van et al., 1976; Bowes et al., 1977; Barko & Smart, 1981; Spencer et al., 1994; Tavechio & Thomaz, 2003). Thus, despite some significant relationships being found between water transparency variables (ZDS and k) and occurrence of both species in sites of the PS, there is little evidence that light alone could be acting as the main factor hindering H. verticillata establishment in floodplain lakes.
Floodplain lakes connected to the River Paraná had water pH values around 6.7 while the River Paraná sustained pH near 7.3. The strong positive relation found between pH and H. verticillata colonization is more likely related to an environmental effect upon the plant than related to a plant effect upon the water column physico-chemical characteristics. Although H. verticillata develops very dense stands in the littoral zone of the River Paraná, the plant occupies a small proportion of the river area (rarely more than some 10 m into the channel from the margin: in a river channel which has a width of c. 2 km in the URPFS). This means that, due to the large quantity of water flowing through the river channels and constant turbulence and mixing pattern of the water column, submersed macrophytes are unlikely to exert any great influence on the water column physico-chemical characteristics of the main river channel. Thus, the significantly higher pH values found in sites of the river channels compared with floodplain lakes probably reflect the environmental characteristics of each habitat (e.g. respiration–production balance), with the river water being more oligotrophic and better aerated by the high water flow and mixing.
Cook & Lüönd (1982) state that H. verticillata develops well in acid oligotrophic waters. Moreover, laboratory studies have shown that the species can photosynthesize even at pH levels below 4 (Van et al., 1976). Thus, it is also unlikely that water pH per se limited H. verticillata development inside floodplain lakes of the PS. Not surprisingly, pH was negatively correlated with O.M.sed (r = −0.50; P < 0.05). Organic matter decomposition in the bottom of lakes tends to be associated with several microbial processes, such as generation of carbon dioxide, nitrification of ammonia or methane fermentation, all of which lead to decreased levels of pH in the water column. Moreover, anaerobic decomposition of organic matter tends to generate substances, such as methane and sulphide, which can cause toxic effects to living organisms (Wetzel, 1983). Indeed, the occurrence of H. verticillata in sites of the PS was better predicted in our study by proportion of organic matter in the sediment. Owing to the lentic characteristics of floodplain lakes, riparian vegetation debris tends to accumulate at the bottom. Thus, levels of sediment organic matter in the floodplain lakes were typically 10 times higher than in the river channel sites. The negative impact of high levels of organic matter in sediment upon H. verticillata has already been shown experimentally by Barko & Smart (1983, 1986) who cited generation of toxic compounds and mechanisms related to nutrient limitations as possible explanations.
References
Agostinho, A. A., S. M. Thomaz & L. C. Gomes, 2004. Threats for biodiversity in the floodplain of the Upper Paraná River. Ecohydrology & Hydrobiology 4: 267–280.
Agostinho, A. A., S. M. Thomaz & L. C. Gomes, 2005. Conservation of the biodiversity of Brazil’s inland waters. Conservation Biology 19: 646–652.
Barko, J. W. & R. M. Smart, 1981. Comparative influences of light and temperature on the growth and metabolism of selected submersed freshwater macrophytes. Ecological Monographs 51: 219–235.
Barko, J. W. & R. M. Smart, 1983. Effects of organic matter additions to sediment of the growth of aquatic plants. Journal of Ecology 71: 161–175.
Barko, J. W. & R. M. Smart, 1986. Sediment-related mechanisms of growth limitation in submersed macrophytes. Ecology 67: 1328–1340.
Bini, L. M. & S. M. Thomaz, 2005. Prediction of Egeria najas and Egeria densa occurrence in a large subtropical reservoir (Itaipu Reservoir, Brazil-Paraguay). Aquatic Botany 83: 227–238.
Bowes, G., T. K. Van, L. A. Garrard & W. T. Haller, 1977. Adaptation to low light levels by Hydrilla. Journal of Aquatic Plant Management 15: 32–35.
Carmouze, J. P., 1994. O metabolismo dos ecossistemas aquáticos. Edgard Blücher/SBL/FABESP, São Paulo.
Chambers, P., P. Lacoul, K. J. Murphy & S. M. Thomaz, 2008. Aquatic macrophytes: world diversity and distribution. Hydrobiologia 595: 9–26.
Cook, C. D. K. & R. Lüönd, 1982. A revision of the genus Hydrilla (Hydrocharitaceae). Aquatic Botany 13: 485–504.
Cook, C. D. K. & K. Urmi-König, 1984. A revision of the genus Egeria (Hydrocharitaceae). Aquatic Botany 19: 73–96.
Figuerola, J. & A. J. Gree, 2002. Dispersal of aquatic organisms by waterbirds: a review of past research and priorities for future studies. Freshwater Biology 47: 483–494.
Golterman, H. L., R. S. Clymo & M. M. Ohmstad, 1978. Methods for physical and chemical analysis of fresh waters. Blackwell Scientific Publications, Oxford.
Haller, W. T. & D. L. Sutton, 1975. Community structure and competition between Hydrilla and Vallisneria. Journal of Aquatic Plant Management 13: 48–50.
Hershner, C. & K. J. Havens, 2008. Managing invasive aquatic plants in a changing system: strategic consideration of ecosystem services. Conservation Biology 22: 544–550.
Kahara, S. N. & J. E. Vermaat, 2003. The effect of alkalinity on photosynthesis-light curves and inorganic carbon extraction capacity of freshwater macrophytes. Aquatic Botany 75: 217–227.
Madsen, T. V. & K. Sand-Jensen, 1987. Photosynthetic capacity, bicarbonate affinity and growth of Elodea canadensis exposed to different concentrations of inorganic carbon. Oikos 50: 176–182.
Madsen, J. D. & D. H. Smith, 1999. Vegetative spread of dioecious Hydrilla colonies in experimental pounds. Journal of Aquatic Plant Management 37: 25–29.
Martins, D., N. V. Costa, M. A. Terra & S. R. Marchi, 2008. Characterization of the aquatic plant communities of 18 reservoirs of five watersheds in Sãol Paulo, Brazil. Planta Daninha 26: 17–32.
Mony, C., T. J. Koschnick, W. T. Haller & S. Muller, 2007. Competition between two invasive Hydrocharitaceae (Hydrilla verticillata L. f. and Egeria densa (Planch)) as influenced by sediment fertility and season. Aquatic Botany 86: 236–242.
Mullin, B. H., L. W. J. Anderson, J. M. DiTomaso, R. E. Eplee & K. D. Getsinger, 2000. Invasive plant species. CAST 13: 1–18.
Murphy, K. J., 1988. Aquatic weed problems and their management: a review – I. The worldwide scale of the aquatic weed problem. Crop Protection 7: 232–248.
Murphy, K. J. & A. H. Pieterse, 1990. Present status and prospects of integrated control of aquatic weeds. In Pieterse, A. H. & K. J. Murphy (eds), The Ecology and Management of Nuisance Aquatic Vegetation. Oxford Publications, New York: 222–227.
Murphy, K. J., G. Dickinson, S. M. Thomaz, L. M. Bini, K. Dick, K. Greaves, M. Kennedy, S. Livingstone, H. McFerran, J. Milne, J. Oldroyd & R. Wingfield, 2003. Aquatic plant communities and predictors of diversity in a sub-tropical river floodplain: the Upper Rio Paraná, Brazil. Aquatic Botany 77: 257–276.
Neiff, J. J., 1990. Ideas para la interpretacion ecologica del Paraná. Interciencia 15: 424–440.
Netherland, M. D., 1997. Turion ecology of Hydrilla. Journal of Aquatic Plant Management 35: 1–10.
Owens, C. S. & J. D. Madsen, 1998. Phenological studies of carbohydrate allocation in Hydrilla. Journal of Aquatic Plant Management 36: 40–44.
Pierini, S. A. & S. M. Thomaz, 2004. Effects of inorganic carbon source on photosynthetic rates of Egeria najas Planchon and Egeria densa Planchon (Hydrocharitaceae). Aquatic Botany 78: 135–146.
Rybicki, N. B., D. G. McFarland, H. A. Ruhl, J. T. Reel & J. W. Barko, 2001. Investigations of the availability and survival of submersed aquatic vegetation propagules in the Tidal Potamic River. Estuaries 24: 407–424.
Souza-Filho, E. E., E. Comunello & P. C. Rocha, 2004. Flood extension in the Baia-Curutuba-Ivinheima complex of the Paraná River Floodplain. In Agostinho, A. A., L. Rodrigues, L. C. Gomes, S. M. Thomaz & L. A. Miranda (eds), Structure and functioning of the Paraná River and its floodplain: LTER – Site 6 – (PELD – Sítio 6). EDUEM, Maringá: 19–24.
Spencer, W. E., J. Teeri & R. G. Wetzel, 1994. Acclimation of photosynthetic phenotype to environmental heterogeneity. Ecology 75: 301–314.
Tavechio, W. L. G. & S. M. Thomaz, 2003. Effects of light on the growth of Egeria najas Planchon. Brazilian Archives of Biology and Technology 46: 203–209.
Thomaz, S. M., L. M. Bini, T. A. Pagioro, K. J. Murphy, A. M. dos Santos & D. C. de Souza, 2004a. Aquatic macrophytes: diversity, biomass and decomposition. In Thomaz, S. M., A. A. Agostinho & N. S. Hahn (eds), The Upper Paraná River and its floodplain: physical aspects ecology and conservation. Backhuys Publishers, Leiden: 331–352.
Thomaz, S. M., T. A. Pagioro, L. M. Bini, M. C. Roberto & R. R. A. Rocha, 2004b. Limnology of the Upper Paraná Floodplain habitats: patterns of spatio-temporal variations and influence of the water levels. In Agostinho, A. A., L. Rodrigues, L. C. Gomes, S. M. Thomaz & L. A. Miranda (eds), Structure and functioning of the Paraná River and its floodplain: LTER – Site 6 – (PELD – Sítio 6). EDUEM, Maringá: 37–42.
Thomaz, S. M., T. A. Pagioro, L. M. Bini & K. J. Murphy, 2006. Effect of reservoir drawdown on biomass of three species of aquatic macrophytes in a large sub-tropical reservoir (Itaipu, Brazil). Hydrobiologia 570: 53–59.
Thomaz, S. M., L. M. Bini & R. L. Bozelli, 2007a. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 579: 1–13.
Thomaz, S. M., P. A. Chambers, S. A. Pierini & G. Pereira, 2007b. Effects of phosphorus and nitrogen amendments on the growth of Egeria najas. Aquatic Botany 86: 191–196.
Van, T. K., W. T. Halle & G. Bowes, 1976. Comparison of the photosynthetic characteristics of three submersed aquatic plants. Plant Physiology 58: 761–768.
Van, T. K., G. S. Wheeler & T. D. Center, 1999. Competition between Hydrilla verticillata and Vallisneria americana as influenced by soil fertility. Aquatic Botany 62: 225–233.
Wang, J., D. Yu, W. Xiong & Y. Han, 2008. Above- and belowground competition between two submersed macrophytes. Hydrobiologia 607: 113–122.
Wetzel, R. G., 1983. Limnology. 2nd ed. Saunders College Publishing, Orlando: 767 pp.
White, A., J. B. Reiskind & G. Bowes, 1996. Dissolved inorganic carbon influences the photosynthetic responses of Hydrilla to photoinhibitory conditions. Aquatic Botany 53: 3–13.
Wingfield, R., K. J. Murphy & M. Gaywood, 2006. Assessing and predicting the success of Najas flexilis (Willd.) Rostk. & Schimidt, a rare European aquatic macrophyte in relation to lake environmental conditions. Hydrobiologia 570: 79–86.
Zagatto, E. A. G., A. O. Jacintho, B. F. Reis, F. J. Krug, H. Bergamin, L. C. Pessenda, R. J. Mortatti & M. F. Giné, 1981. Manual de análises de plantas empregando sistemas de injeção em fluxo. Universidade de São Paulo, Piracicaba. 45.
Acknowledgements
The first author thanks the Brazilian Council of Research (CNPq) and the Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for funding his PhD course. We thank Agaito Barnabé and Sebastião Rodrigues for helping with the field work. Partial funds were provided by CNPq/MCT, through the Long Term Ecological Research Program (PELD, site number 6). S. M. Thomaz thanks the Brazilian Council of Research (CNPq) for ongoing funding through a Research Productivity Grant.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Luis Mauricio Bini
Rights and permissions
About this article
Cite this article
Sousa, W.T.Z., Thomaz, S.M., Murphy, K.J. et al. Environmental predictors of the occurrence of exotic Hydrilla verticillata (L.f.) Royle and native Egeria najas Planch. in a sub-tropical river floodplain: the Upper River Paraná, Brazil. Hydrobiologia 632, 65–78 (2009). https://doi.org/10.1007/s10750-009-9828-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-009-9828-3