
Abstract
Mayflies (Insecta: Ephemeroptera) are common inhabitants of streams throughout the Appalachian Mountains. Headwater mayfly assemblages were evaluated with respect to regional landuse disturbances (coal mining and residential) in eastern Kentucky, USA. Estimates of mayfly taxa richness and relative abundance were compared at 92 sites represented by least-disturbed reference (REF; n = 44), residential only (RESID; n = 14), mixed residential and mining (MINED/RESID; n = 14), and mining only (MINED; n = 20) landuse categories. A total of 48 species from 27 genera and 9 families were identified; Ephemerella, Epeorus, Ameletus, Cinygmula, and Paraleptophlebia comprised the core 5 genera most frequently encountered at REF sites. These same genera (among others) were often reduced or extirpated from other landuse categories. Mean mayfly richness and relative abundance were significantly higher at REF sites compared to all other categories; MINED sites had significantly lower metric values compared to RESID and MINED/RESID sites. Relative mayfly abundance was most strongly correlated to specific conductance (r = 0.72) compared to total habitat score (r = 0.59), but relationships varied depending on landuse category. Non-metric multidimensional scaling (for mayfly taxa) and principal components analysis (for environmental variables) separated REF sites strongly from most other sites. The results indicate that expected mayfly communities are disappearing from streams where mining disturbance and residential development has occurred and because of the long-term impacts incurred by both landuses, recovery is uncertain.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Mayflies (Insecta: Ephemeroptera) have long been recognized as important indicators of stream health. Because of their sheer ubiquity and abundance in natural streams of the Appalachian Mountains, mayflies justifiably represent a component of the aquatic life that water quality regulations intend to protect. Analogous to the loss of other indigenous organisms from streams due to environmental stress (e.g., those receiving considerable public attention such as trout or bass), many mayflies are increasingly at risk from human disturbance in the Appalachians and show patterns of extirpation from streams disturbed by particular landuses.
Mayflies are found in a variety of aquatic systems ranging from vernal ponds to lakes and streams to estuaries (Barber-James et al., 2008). However, they reach their greatest diversity and abundance in rocky headwater streams. Morse et al. (1997) reported a total of 238 species from 63 genera and 16 families in the southeastern U.S. In the Appalachian Mountains of NC and SC, states that share many species with KY, Morse et al. (1997) reported 124 species of mayflies while Parker et al. (2007) recorded 124 species from the Great Smoky Mountain National Park (on the TN/NC border) alone. Kondratieff (2000) tentatively listed 109 species from KY but this state has not been thoroughly surveyed at the species level (Randolph & McCafferty, 1998). In general, lotic mayflies have been the subject of many freshwater biomonitoring studies ranging from empirical field observations (e.g., Brittain & Saltveit, 1989; Wallace & Gurtz, 1986; Moog et al., 1997) to controlled toxicological experiments (e.g., Goetsch & Palmer, 1996; Chadwick et al., 2002; Kennedy et al., 2003; Beketov, 2004; Hassell et al., 2006; Brinkman & Johnston, 2008). Ephemeroptera also fill important trophic roles in stream ecosystems, as displayed by their diverse functional feeding group designations. Their contribution to the overall structure and functioning of stream ecosystems has evoked calls for regional or global conservation (Morse et al., 1997; Barber-James et al., 2008)].
Bauernfeind & Moog (2000) established an approach to using Ephemeroptera in the assessment of ecological integrity in Central Europe while other researchers have used particular mayfly families (e.g., Baetidae; Buss & Salles, 2007) or assemblages (Moog et al., 1997; Courtney & Clements, 2000) as indicators of stream degradation. Previous studies from the Appalachian region revealed strong negative impacts to Ephemeroptera from surface coal mining in West Virginia (Chambers & Messer, 2000; Hartman et al., 2005; Merricks et al., 2007; Pond et al., 2008). Acidic deposition has also affected benthic communities in streams throughout the region (Herlihy et al., 1998; McClurg et al. 2007). Localized urbanization effects on benthos in headwater streams are commonly reported (Paul & Meyer, 2001; Cuffney et al., 2005; Blakely et al., 2006; Smith & Lamp, 2008), but few studies have focused on water quality and loss of taxa associated with rural residential landuse issues in the Appalachian coalfields (see Green et al., 2000; Soucek, 2001). I evaluated data from 92 headwater streams in the Appalachian Mountains of Kentucky to explore and describe regional patterns of diversity and distribution of lotic Ephemeroptera in relation to two common and pervasive stressors: coal mining and rural residential landuses.
Materials and methods
Study area
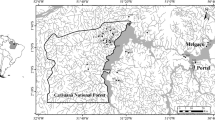
The Appalachian Mountains of eastern Kentucky encompass three distinct Level III ecoregions (Fig. 1; Southwestern Appalachian, Central Appalachian, and Allegheny Plateau; Woods et al., 2002) and are characterized by highly dissected terrain with diverse forest types, geology, and climate. Elevation of the region ranges from ~150 m in river valleys to >1,000 m on ridges. Within the study area, bedrock geology is mostly sedimentary and consists of interbedded sandstones, siltstones, shale, and coal (Woods et al., 2002), and the dominant vegetation is part of the temperate, mixed-mesophytic forest classification (Braun, 1950). Common tree species found along least-disturbed streams include eastern hemlock, beech, maples, oaks, hickories, basswood, buckeye, and yellow polar. Common shrubs include spicebush, witch hazel, pawpaw, rhododendron, hydrangea, and ironwood. Headwater streams in this region are cool and dilute (i.e., low dissolved solids) and typically flow through constrained valleys with relatively high gradients and have boulder–cobble substrates. Precipitation patterns are generally uniform throughout the study region (Woods et al., 2002). With regard to headwater macroinvertebrates, community structure is relatively uniform in this bioregion (Pond and McMurray, 2002) but distinct from other ecoregions in Kentucky (see Pond et al., 2003).

Generalized map of study area showing level 3 ecoregion designations (after Woods et al., 2002) in eastern Kentucky. Boxes represent sampling areas and number of sites per area. West Allegh Western Allegheny Plateau, Cen App Central Appalachians, SW App Southwestern Appalachians
Data for this study were filtered from the Kentucky Department for Environmental Protection (KYDEP) ecological database; sites were compiled from moderately high gradient (~2–5%), cobble–boulder dominated 1st–3rd order streams that were sampled throughout the Eastern KY coalfield region. Regional landuse stressors include coal mining, residential development, logging, oil and gas drilling, and light agriculture (e.g., pasture and row crops). Surface coal mining and residential landuses are currently the most common and long-term stressors to headwater streams in the Appalachian region of KY. Surface mining occurs on steep slopes and narrow ridges where headwater valleys are used for overburden and spoil disposal. In contrast, residential landuse occurs within the riparian corridor because of topographic limitations in these narrow valleys.
Site selection
As part of Kentucky’s routine monitoring program, 92 Appalachian headwater streams (defined herein as sites draining <10 km2) were sampled once for macroinvertebrates in the spring (late-Feb–late May), spanning a 5 year period (1999–2004). Spring-time sampling often yields the most taxa and abundance in Appalachian headwater streams (KDEP unpub. data). A 100-m sampling reach was established in “typical” or “representative” segments of each stream. This meant that sites were situated in relatively natural channels free from direct influences of bridges, culverts, dredging, or artificial substrates. Although the majority of “reference” sites (REF; total n = 44) were located within state research forests, state parks, and the Daniel Boone National Forest, many of the watersheds had some mild disturbance associated with past logging (i.e., most REF sites drained second and third growth forest), roads, and gas wells and thus were not considered “pristine”. In fact, REF watersheds were ravaged by clearcut logging and steep slope agriculture in the early twentieth century.
Efforts were also made to sample a gradient of more recent disturbances and to categorize sites into different landuse or stressor types in the presumed absence of other stressors. Since digitized landcover in the watershed was not sufficiently accurate for quantification in these small watersheds (i.e., outdated or misinterpreted), site classification was accomplished by examining aerial photos, topographic maps, mining permit information, number of houses upstream of sample reaches, and actual conditions in the field. Sites with coal mining only (MINED; n = 20), rural residential only (RESID; n = 14), or mixed mining and residential landuse (MINED/RESID; n = 14) were used as grouping variables to document patterns of mayfly distribution. This generalized classification scheme assured a sufficient number of sites for group-type analysis. Sites were excluded from the dataset if they explicitly failed to meet these individual landuse scenarios (e.g., other commercial/industrial, agriculture). Overall, streams were small but perennial, with catchment areas that averaged 3.1 km2 at REF sites, 2.6 km2 at MINED sites, 3.9 km2 at RESID, and 5.9 km2 at MINED/RESID.
Despite natural variation in precipitation across sampling years, seasonal flows were ample and considered normal at the time of all spring collections. Nearly ½ of the sites were sampled in 2000 (all landuse categories), the spring following both 1998 and 1999 summer and fall droughts (NOAA, 2009). It was thought that some REF sites could have dried during the summer of 1998 and 1999. However, spring 1999 and 2000 samples were collected during typical spring flows and showed abundant macroinvertebrate populations at sites. Green et al. (2000) sampled benthos in headwater streams in WV and noted that this drought affected their summer and fall samples, but not their spring samples. Spring 2001 precipitation was slightly below average but stream discharge was good at the time of sampling and sites yielded typical abundance values of macroinvertebrates. All other sample events occurred when proceeding months experienced normal amounts of rainfall (NOAA, 2009). For these reasons, hydrological condition at the time of sampling was considered to be similar across all sites and years.
Although acid mine drainage (AMD; pH < 5) is also present throughout much of this region, I selected non-AMD, alkaline streams because AMD often eliminates all benthic organisms and these data would, therefore, be less useful for making mayfly assemblage comparisons. All MINED sites were relatively alkaline (pH > 7) and most located several hundred meters downstream of mountaintop mining areas with headwater valleyfills (mining methods detailed in Slonecker & Benger, 2002); a few MINED sites contained only contour mining operations >20 years old. RESID sites had >5 houses upstream of the 100 m sample reach and paved or gravel roads often paralleled or crossed streams. Some RESID and MINED/RESID sites had relatively high housing density (>60 houses/linear km) within the upstream corridor. These same streams’ uplands were >90% forested. In eastern KY, steep terrain and narrow valleys limit upland residential development and most roads and houses are located in valleys and within the riparian corridor. In contrast, many MINED sites had mostly forested stream corridors near the sampling reach but very little or no forest in the adjacent uplands and extreme headwaters where mountaintop mining and valley filling occurred. Sites downstream of MINED/RESID had a mix of upland and riparian disturbance with surface mining and >5 houses upstream of the sampling reach. In terms of streamside vegetation, RESID and MINED/RESID sites commonly had exotic multiflora rose, Japanese and tartarian honeysuckle, and Japanese knotweed along their banks.
Sample collection
Mayflies were collected through riffle kicknet sampling with a 0.5-m wide, 595 μm mesh net supplemented by multihabitat samples following KYDEP standard operating procedures (KYDEP, 2008). These techniques are used by KYDEP for statewide monitoring and assessment of macroinvertebrate communities in streams and rivers. Briefly, four 0.25 m2 samples were collected in cobble riffles from a 100-m sampling reach and composited into a 1-m2 sample. Care was taken to position individual kicknets in similar substrate and current velocities across all sites to minimize these important factors’ influence on benthos distribution. The standardized-effort multihabitat samples were composited from leafpacks, root and bank D-net sweeps, and hand-picking large boulders, and woody debris. In the laboratory, entire picks of the 1-m2 sample, and multihabitat collections were done, but for analytical purposes, only the riffle sample was quantified for this study; new taxa collected in the multihabitat sample were simply added for richness and presence/absence comparisons. All mayflies were identified to the species level but this proved difficult for some samples that had early instar or damaged specimens. For comparisons across landuse categories, mayflies were collapsed to the genus level in the database.
Co-occurring habitat and chemical data collection included the Rapid Bioassessment Protocol Habitat Assessment (RBP habitat scores after Barbour et al., 1999), and in situ physicochemical measurements (pH, specific conductance, dissolved oxygen, and temperature). The RBP habitat assessment evaluates important habitat components such as epifaunal substrate quantity and quality, embeddedness, velocity/depth regimes, sediment deposition, channel flow status and channel alteration, stream bank stability, bank vegetation protection, and riparian zone width. Each component was scored on a 20-point scale with a total possible summed score of 200. At few sites, a nearly full suite of chemical analytes (e.g., metals and nutrients) were measured from grab samples taken at the time of benthic sampling but were only used for site-specific interpretations.
Data analysis
A combination of multivariate (ordination) and bivariate (Spearman correlation, scatterplots, boxplots) techniques were used to explore patterns of mayfly distribution among the disturbance categories (grouped as REF, RESID, MINED/RESID, and MINED). Mayfly metrics included mayfly richness, mayfly relative abundance (% mayflies), and total proportion of mayfly taxa to total taxa richness. A nonparametric Kruskal–Wallis 1-way analysis of variance was used to test for differences in mayfly richness and relative abundance estimates across landuse categories and paired Mann–Whitney U-tests were done to compare means between each pair of categories. Ordination was accomplished with non-metric multiple dimensional scaling (NMS; PC-ORD v. 4.25, McCune & Mefford, 1999) using the Bray–Curtis similarity coefficient (Bray & Curtis, 1957; McCune & Grace, 2002) based on log10 (x + 1) abundances. The data were computed with 225 iterations, 40 real runs, and 50 randomized runs.
Disturbance groups were also compared with the nonparametric multi-response permutation procedure (MRPP; PC-ORD v. 4.25, McCune & Mefford, 1999) to determine if mayfly assemblage distance differed between disturbance categories. Here, ranked Sorenson distances from the 92 sites were used to test the hypothesis of no difference among categories. MRPP produced an A-statistic, which compared observed versus expected within-site homogeneity based on the distance matrices (positive A-values indicate higher within-site homogeneity than expected by chance, i.e., differences in invertebrate composition between sites) and a P value indicating statistical significance.
In order to explore environmental differences in landuse categories, a principal components analysis (PCA; MVSP v. 3.1, Kovach Computing, London) was performed on log10 (x + 1) transformed environmental variables from 80 of the 92 sites. Twelve sites (4 REF, 3 RESID, 2 MINED/RESID, and 3 MINED) were not analyzed because of missing data (e.g., failed D.O., conductivity, or pH sensor at the time of sampling); PCA requires a symmetric dataset with no missing values. Refer to Table 2 for list of environmental variables analyzed. In order to test for differences in environmental variables between landuse types, Mann–Whitney U-tests were performed. A non-parametric changepoint analysis using the deviance reduction method (R Development Core Team, 2009) was run to determine potential breakpoints or thresholds in environmental–mayfly relationships. In order to determine uncertainty around the mean changepoint, 90% confidence intervals were generated by bootstrap resampling with replacement (1,000 iterations).
Results
Environmental comparisons
Summary statistics for environmental variables are shown in Table 1. Temperature and D.O. (at the time of sampling) did not differ across all landuse categories (Mann–Whitney U-tests; P < 0.05). Specific conductance and pH were significantly lower (Mann–Whitney U-tests; P < 0.05) at REF sites (mean = 51 μS/cm and 6.8 S.U., respectively) than all other disturbed categories. Similarly, RESID sites had significantly lower specific conductance (mean = 242 μS/cm) than MINED (mean = 940 μS/cm) and MINED/RESID (mean = 585 μS/cm) but no difference was detected between MINED and MINED/RESID sites (P > 0.05). Furthermore, no differences were found in pH among the three disturbance categories (P > 0.05). Measures of sedimentation (i.e., embeddedness and sediment deposition scores), riparian zone width, and bank vegetation cover within the three disturbance categories were significantly different from REF sites, but differences between the disturbance types were not significant (P > 0.05).
The PCA exhibited strong separation of REF sites from the bulk of the other three landuse categories (Fig. 2). Axis 1 accounted for nearly 40% of the variance (eigenvalue = 4.7) while axis 2 accounted for only 11.4% (eigenvalue = 1.4). Table 2 shows factor coefficients for environmental variables and indicate that most of the habitat metrics, pH, and specific conductance contributed strongly to the ordination. Riffle embeddedness score and epifaunal substrate score had the highest coefficients (0.81) on axis 1. Temperature and D.O. did not greatly contribute to the ordination but they were measured only once at each site (these two parameters are typically the most variable measurements in stream datasets and so more data would be needed to evaluate their effects).

Principal components analysis ordination for subset of sites. Based on 40 REF sites, 11 RESID sites, 12 MINED/RESID sites, and 17 MINED sites
Mayflies encountered in headwater streams
Springtime collections from the 92 headwater sites yielded a total of 12,640 mayflies representing 48 distinct mayfly taxa from 27 genera, and 9 families. Headwater mayfly assemblages in REF streams were dominated by ephemerellids (chiefly Ephemerella and Drunella), heptageniids (mostly Epeorus and Cinygmula), the ameletid mayfly, Ameletus, and the leptophlebiid (Paraleptophlebia). A list of all taxa and their mean abundance at sites within each landuse category is shown in Table 3. I chose to report mean abundance of individual mayfly genera because median abundance was zero for most taxa within the three disturbed categories. The relative frequency (presence/absence) of taxa observed by landuse category is shown in Fig. 3. The greatest disparity in mayfly presence/absence (i.e., reduction in proportion of sites where taxon was observed compared to REF) was seen with several key taxa: Ephemerella, Ameletus, Epeorus, Cinygmula, and Paraleptophlebia. For example, Epeorus was collected at 41 of 44 REF sites (93%) but was absent from 19 of 20 (95%) MINED sites. A few mayflies which were not typical of REF headwater streams (Fig. 3) were found more frequently at RESID and MINED/RESID sites (e.g., Isonychia and Caenis).

Relative frequency (%) of occurrence for all mayfly genera across all landuse categories
Richness and abundance across Landuse categories
The average proportion of mayfly richness to total benthic taxa richness in the sample was 20% at REF sites, 17% at RESID sites, 14% at MINED/RESID sites, and 6% at MINED sites. Mayfly relative abundance (% mayflies, Fig. 4A) and mayfly richness (Fig. 4B) differed across landuse categories. Compared to REF, % mayflies and mayfly richness were significantly reduced in RESID, MINED/RESID, and MINED categories (Kruskal–Wallis, P < 0.0001) although a few sites within each of these categories yielded similar types and numbers of mayflies as the least-disturbed reference sites. MINED streams had significantly lower % mayflies and mayfly richness than any other category (Mann–Whitney U-tests, P < 0.02). RESID and MINED/RESID did not statistically differ in % mayflies (Mann–Whitney U-test, P = 0.49) or mayfly richness (Mann–Whitney U-test, P = 0.32). In KY, the fifth percentile of the REF distribution for biological condition represents a threshold for determining attainment of ALU using multimetric indices (e.g., Index of Biotic Integrity). Using this same benchmark approach, the degree of departure of mayfly metrics from REF conditions indicated that the majority of sites in each disturbed category fell far below the calculated threshold. For mayfly abundance (threshold = 21%), 71% of RESID, 75% of MINED/RESID, and 90% of MINED sites fell below the threshold. For richness (threshold = 6 genera), 64% of RESID, 73% of MINEDRESID, and 90% of MINED sites fell below the threshold.

Boxplots of relative abundance (%) of mayflies (A) and mayfly species richness (B) among landuse categories. Letters beside individual categories show results of Mann–Whitney U-test where matching letters indicate no significant difference (P < 0.05). Box lines contain lower and upper quartiles and median; whiskers represent range excluding outliers (asterisks and open circles)
Mayfly relative abundance was more strongly correlated to specific conductance (Fig. 5A; r = 0.72) compared to the total RBP habitat score (Fig. 5B; r = 0.59). A strong threshold-type response relationship between % mayflies and increasing specific conductance was seen where significant loss of mayflies occurred when specific conductance was >175 μS/cm (changepoint analysis; lower 90% CL = 124 μS/cm, upper CL = 336 μS/cm). Those MINED sites having relatively high % mayflies had corresponding low to moderately elevated specific conductance (Fig. 5A) or had high numbers of facultative mayflies such as Baetis.

Scatterplot of % mayflies vs. specific conductance (A) and % mayflies vs. total RBP habitat score (B) among landuse categories. Solid vertical lines represent mean changepoint; dashed vertical lines represent 90% confidence interval
For all pairs of site habitat scores and % mayflies, (Fig. 5B), the changepoint analysis returned a habitat score of 145 (lower 90% CL = 139, upper 90% CL = 151) at which point mayflies were significantly greater above this score. Habitat score did not significantly account for loss of mayflies within the MINED category (r = 0.39, P > 0.05). At RESID sites, % mayflies was significantly related (P < 0.05) to total RBP habitat scores (r = 0.56) but not at MINED/RESID sites (r = 0.38, P > 0.05). Other habitat metrics that were relatively strongly correlated to % mayflies across all sites included embeddedness score (r = 0.62) and epifaunal substrate score (r = 0.65). Although % mayflies varied among REF sites (Fig. 4A), there was no clear indication that this metric was significantly (P > 0.05) related to stream size (catchment area; r = 0.02), geography (latitude, r = −0.23; longitude, r = −0.19), seasonality (julian day for spring index period; r = 0.19), habitat quality (r = 0.06), specific conductance (r = −0.17), or pH (r = 0.21). Furthermore, mayfly richness at REF sites was not related to total number of individuals in the sample (r = 0.12, P = 0.43).
Ordination of the Mayfly assemblage
A 3-dimensional solution was recommended by the software with a final stress of 14.8%. NMS axis 1 accounted for 44% of the variance compared to axis 2 (27%) and axis 3 (20%). REF sites were fairly well-clustered in 2 dimensional ordination space while assemblages among the three landuse categories were more spaced apart (Fig. 6). Mayfly genera most strongly associated with REF sites plotting positively along NMS axes 1 and 2 included Ephemerella, Drunella, Cinygmula, Epeorus, and Ameletus. Facultative and tolerant mayflies negatively associated with axes 1 and 2 were Caenis, Baetis, Isonychia, and Stenonema. No clear pattern of mayfly assemblage structure was observed for the three landuse categories, however, nearly half of the mined sites could not be ordinated because they contained no mayflies.

Nonmetric multidimensional scaling ordination for riffle samples at sites having 1 or more mayfly taxa. Percent of the variance explained by each axis shown in parentheses
The separation of sites in NMS ordination space, grouped by landuse disturbance categories, was significant based on the MRPP (A = 0.190, P < 0.0001). Comparatively, REF within-site similarity was significantly higher than RESID (A = 0.104, P < 0.0001), MINED/RESID (A = 0.148, P < 0.0001), and MINED (A = 0.124, P < 0.0001). RESID and MINED sites were also significantly dissimilar, albeit much weaker (A = 0.07, P < 0.01). However, mayfly assemblages were indistinguishable between RESID sites and MINED/RESID sites (A = −0.007, P = 0.54) and between MINED/RESID sites and MINED sites (A = 0.002, P = 0.21).
Discussion
During the spring season, mayflies naturally represent ~25–50% of the total abundance and account for ~1/5th of all taxa collected from headwater riffle samples in relatively undisturbed reference streams in the Appalachian Mountains (this study; Pond et al., 2008), they contribute considerable biodiversity to regional streams (Morse et al., 1997), and they are highly sensitive to environmental degradation. For these very reasons, Ephemeroptera are of critical importance and serve to indicate levels of human disturbance in streams. Water pollution protection laws were written to protect indigenous aquatic life such as sensitive mayflies. States and tribes in the U.S. are required under the U.S. Clean Water Act to designate aquatic life uses (ALU) and to establish water quality standards (WQS; narrative and numeric criteria for pollutants or conditions) to protect the aquatic life inhabiting the waterbodies within their jurisdiction. There are few states that have promulgated numeric biological criteria for ALUs based on macroinvertebrates or fish; most rely on the use of biological data to translate narrative statements that serve as formal criteria in their WQS.
In Kentucky headwater streams, mayflies responded negatively to two pervasive landuses, rural residential development, and surface coal mining. Absence of mayflies certainly translates to impairment of the ALU based on Kentucky narrative WQS, especially when the impairment is linked to specific chemical narrative criteria. For example, the Kentucky regulatory narrative standard (KYDEP, 2007) for ionic strength states that: “…conductivity shall not be changed to the extent that the indigenous aquatic community is adversely affected.” This WQS would apply and might be exceeded in particular streams where wholesale loss of common mayfly genera from MINED sites with highly elevated conductivity was observed. Until recently, there has been little focus on ionic stress-type criteria for waters within Appalachian states (Bodkin et al., 2008).
Landuse induced effects on Mayfly Loss
A recent review by Weijters et al. (2009) on catchment disturbance and biodiversity indicated that a 10% reduction in natural catchment landcover results in a 6% loss of native species; and 50% reduction results in a 25% loss of Ephemeropter + Plecoptera + Trichoptera (EPT) taxa. Land use area changed by surface mining in small Appalachian catchments frequently totals 25–75% of the contributing catchment but often results in wholesale extirpation (~100% reduction) of sensitive mayflies. Much of the land cover change due to surface mining in Appalachia is in the loss of spatially defined “interior” forest (Wickham et al., 2007), a more ecologically sensitive forest type compared to generalized forest landcover. However, the loss of taxa also depends more on the exceptionally high chemical loading to the receiving water than on the total area of watershed disturbed (Hartman et al., 2005; Pond et al., 2008). As with acid mine discharge, toxicity of alkaline surface mining effluent to some mayflies might result from exposure to or ingestion of trace heavy metal compounds (Clements et al., 1992; Clements 1994, 2004), or from interference with osmoregulation (i.e., gill function and respiration) by the rapid increase in conductivity and component ions (Kennedy et al., 2003). Some mayflies found in this study appeared to be less sensitive to increases in ionic strength (e.g., Baetis, Isonychia, and Caenis). Although Pond et al. (2008) did not find strong correlations of mayfly abundance and richness with mining-related trace metals in the water column (except Se), they state that possible exposure to metals through dietary uptake (Buchwalter & Luoma, 2005; Buchwalter et al., 2007) or potential microhabitat smothering by metal hydroxide precipitate (U.S. Environmental Protection Agency, 2005) or iron bacteria blooms (Wellnitz et al., 1994) could not be ruled out. Analyses from WV mining areas (Hartman et al., 2005; Merricks et al., 2007; Pond et al., 2008) indicated that the decline of mayflies from mountaintop mining correlates most strongly to specific conductance. However, Pond et al. (2008) showed that all individual anions and cations except for Na and Cl were strongly related to mayfly metrics in mined watersheds in the Central Appalachian ecoregion. While increased SO4 concentration is most popularly associated with coal mining activities, other potentially toxic ions (e.g., Mg or HCO3; after Mount et al., 1997) are highly elevated downstream of Appalachian coal mining operations (Bryant et al., 2002). The ionic makeup of mined watersheds in the Appalachian coalfields is relatively consistent and predictable with four principal ions: SO4 > HCO3 > Ca ≈ Mg; K, Na; and Cl are only slightly elevated from surface coal mining (Wunsch et al., 1996; Bryant et al., 2002; Pond et al., 2008). Clearly, further research on the mechanisms of mayfly ionic toxicity, including individual ion or synergistic effects, is warranted.
Mayflies were also affected by habitat degradation (total RBP habitat score) when compared across all sites, but not significantly within the MINED category. Similarly, Pond et al. (2008) found only weak relationships between mayfly metrics and RBP habitat parameters downstream of mined Central Appalachian headwater streams in WV. This suggests that degradation of water quality and the resultant increases in specific conductivity, component ions, and trace metals limit aquatic life regardless of habitat quality. This has implications for the use of natural channel design as a stream restoration tool in these regions where full attainment of the aquatic life use is the goal of the restoration. Streams with highly degraded water quality will not fully attain their ALU by restoring physical habitats alone, and thus stream restoration following a “Field of Dreams Hypothesis” approach (e.g., Bond & Lake, 2003; Hughes, 2007) will probably not succeed in mining- or urban-impacted streams where water quality drives the structure of invertebrate assemblages. Hence, the first obvious step in rehabilitating streams in this region should be to control water quality associated with discharge from mining and residential landuses.
Cuffney et al. (2005) reported that nearly all genera of mayflies were extirpated from high intensity urban streams in three separate geographic areas of the USA and proposed urban-tolerance values for many benthic taxa. “Urban” intensity varied in my study, but most RESID sites had low intensity with only a scattering of houses upstream of the sample reach. Soucek (2001) found that urbanization effects on benthos were often worse than mining effects in Appalachian streams of Virginia; however, conductivity was relatively low in his dataset, except where AMD occurred. The loss of mayflies from some RESID sites that had elevated nutrients or organic wastes could be due to observed filamentous bacterial infestations. This assumption is supported by field and laboratory studies from Lemly (1998, 2000) that showed 100% mortality of headwater mayfly taxa (e.g., Epeorus) when their bodies were more than 25% covered in the sewage-bacterium, Sphaerotilus. Lemly also reported that even low to moderate increases in nitrogen and phosphorus can stimulate blooms of filamentous bacteria in normally nutrient-poor stream systems in the Appalachian Mountains. Although not quantified, Sphaerotilus infestations were commonly observed on various taxa at RESID and MINED/RESID sites. It is expected that RESID and MINED/RESID sites had elevated nutrient concentrations where improper on-site wastewater treatment systems (i.e., poorly designed septic systems and “straightpipes”) were common and thus provided a plausible source of energy for Sphaerotlilus infestations. In addition to the increase in nutrients from residential landuse, discharge of household chemicals and detergents directly into streams can cause harm to aquatic organisms. Soapy or oily discharges from gray-water “straightpipes” are commonly seen in small eastern KY streams. Other likely pollutants from homes include oil, grease, garbage, animal wastes, solvents, paint and masonry wastes, detergents, and pesticides and fertilizers. Hence, mayflies might be exposed to a potpourri of potentially harmful chemicals in residential areas. Until recently, there has been very little or no regulatory oversight on wastewater issues for rural residential development in eastern KY.
Recolonization potential and implications for conservation
Both the timing and magnitude of disturbance events structure benthic macroinvertebrate assemblages (Wallace, 1990). Studies show that some Appalachian mayflies can quickly re-colonize habitats and even increase in abundance following pulse-type disturbances like clearcut or selective logging (Haefner & Wallace, 1981; Stone & Wallace, 1998; Kreutzweiser et al., 2005), catastrophic floods and debris flows (Snyder & Johnson, 2006), or experimental substrate disturbance (Reice, 1985). However, some mayflies may be permanently extirpated from streams where press-type disturbances like coal mining and urbanization cause long-term habitat modifications and incur unabated chemical stress to downstream benthic communities. Nevertheless, chronic effects (i.e., communities continue to display differences in community structure compared to reference or control sites) from any disturbance might last for weeks to several decades (see Wallace, 1990). Data from Merricks et al. (2007) and Pond et al. (2008) indicate that mayflies were still absent or severely reduced in WV streams >15 years after mining ceased.
Sustained impacts to these headwater streams cause problems for re-colonization by indigenous macroinvertebrate communities. In most Appalachian surface mining situations, much of the intense disturbance occurs at the stream origin and progresses downstream, which means that few or no organisms may be available to re-colonize the affected streams through drift after elimination of the organisms by physical disturbance or chemical toxicity. Aerial dispersal from adjacent tributaries (if not impacted) would be the only source of colonization but is likely a slow process. By comparison, RESID sites usually have intact uplands and the headwaters do provide a source of colonization. This might partially explain why some sensitive mayflies were not entirely extirpated from RESID sites (i.e., they occurred as transient, drifting propagules). Many aquatic insects including mayflies have low lateral dispersal ability and adults seldom fly more than 100 m perpendicular to the stream channel (Griffith et al., 1998; Petersen et al., 2004), but adult Baetis (an opportunist genus in this region) have been reported up to 1 km from the stream (Hershey et al., 1993).
In the highly dissected terrain of the Appalachians, headwater populations can become quickly isolated when receiving streams are cumulatively being degraded by increasing human disturbance. However, if a network of headwater streams is intact, colonization of individual disturbed sites within the network would presumably occur faster based simply on proximity and a higher availability of propagules. In such a network, downstream drift of larvae to the confluence of a joining headwater stream could potentially provide the adult stage an opportunity to colonize an adjacent stream (Griffith et al., 1998) and upstream longitudinal colonization might occur through subsequent generations. Although Masters et al. (2006) documented inter-basin transfer of adults from forested sites to nearby acidified streams, they suggested other factors such as stressor persistence and insect mating or oviposition behavior could delay recovery. Unfortunately, many reclaimed mined lands are basically tree-less, as most companies have revegetated large tracts of land with herbaceous species, further prolonging recovery of stream communities by fragmenting potential routes of insect dispersal. Future research should focus on the temporal and spatial influences on recolonization potential in streams disturbed by mining and rural residential development in Appalachian watersheds.
Temporal and spatial scales of both natural and anthropogenic disturbance types must be evaluated before attempting to rehabilitate impaired waters. For example, in Maryland, Smith & Lamp (2008) detected significant urban impacts to headwater benthos (including mayflies), they reported that longitudinal position of individual reaches in relation to urban landuse and proximity to their receiving mainstem streams accounted for more variation than direct habitat and chemical effects. Alexander (2007) found that Ephemerella (a dominant genus in my study) recolonization rates were decreased in deforested Maryland headwaters following drought compared to intact forested sites. However, data from clearcut logging experiments in the southern Appalachians have shown that while mayfly genera densities shift, they are seldom extirpated from logged reaches and might resume normal densities in 5–10 years as canopy cover closes and food resources shift back to pre-logging distributions (Wallace & Gurtz, 1986; Wallace et al., 1988). This is in strong contrast to many of the MINED sites in KY where, despite similar logging intensity associated with the mining operation, most mayfly taxa were completely eliminated from affected watersheds and confirm chronic chemical disturbance due to coal mining. These results coincide with relationships between mayflies and increasing specific conductance found in WV-mined catchments (Pond et al., 2008).
Conclusion
Based on abundance and frequency of occurrence, the mayfly genera best represented in least-disturbed eastern KY streams were Ephemerella, Ameletus, Epeorus, Paraleptophlebia, and Cinygmula and form the basic core taxa expected in healthy Appalachian headwaters of KY. These genera were the most sensitive to anthropogenic disturbance and are thus appropriate for monitoring impacts in these systems. Only two species encountered in this study (Epeorus namatus and Leucrocuta thetis) are listed as regionally vulnerable. However, habitat fragmentation and deforestation have been shown to lead to loss of genetic diversity in some Ephemerella populations (Alexander, 2007) and this scenario is probable for other mayfly taxa. Barber-James et al. (2008) indicated that while very few mayflies are globally “red-listed” on the IUCN, it is because of a lack of knowledge on most species distribution and ecology. Of regional importance is the documented extirpation of mayfly species, genera, and even families from particular landuses and associated stressors (e.g., urbanization, mining, and acid rain) despite the natural ubiquity, fecundity, and resiliency of most Ephemeroptera, and existing laws to protect them. Clearly, habitat and water quality factors play a role in structuring the mayfly assemblages in Appalachian headwater streams of KY. Specific conductance was found to be an excellent predictor of mayfly abundance and richness and should be monitored closely in association with human disturbance and subsequent remediation efforts across the region. Because of the enduring and widespread occurrence of residential and mining influenced streams across the Appalachian coalfields, preserving sufficient undisturbed watersheds in the immediate vicinity of these disturbances is crucial to maintain regional and local mayfly biodiversity by providing refugia, a potential source for future recolonization, and to provide freshwater dilution for chemical stressors affecting Appalachian headwaters.
References
Alexander, L. C., 2007. Genetic diversity and persistence of mayfly populations in disturbed headwater streams. PhD. Dissertation. University of Maryland, College Park, MD.
Barber-James, H. M., J. Gattolliat, M. Sartori & M. D. Hubbard, 2008. Global diversity of mayflies (Ephemeroptera, Insecta) in freshwater. Hydrobiologia 595: 339–350.
Barbour, M. T., J. Gerritsen, B. D. Snyder & J. B. Stribling, 1999. Rapid Bioassessment Protocols for use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates, and Fish, Second Edition. EPA 841-B-99-002. U.S. Environmental Protection Agency, Office of Water, Washington, DC.
Bauernfeind, E. & O. Moog, 2000. Mayflies (Insecta: Ephemeroptera) and the assessment of ecological integrity: a methodological approach. Hydrobiologia 422(423): 71–83.
Beketov, M. A., 2004. Different sensitivities of mayflies (Insecta, Ephemeroptera) to ammonia, nitrite and nitrate: linkage between experimental and observational data. Hydrobiologia 528: 209–216.
Blakely, T. J., J. S. Harding, A. R. Mcintosh & M. J. Winterbourn, 2006. Barriers to the recovery of aquatic insect communities in urban streams. Freshwater Biology 51: 1634–1645.
Bodkin, R., J. Kern, P. McClellon, A. J. Butt & C. Martin, 2008. Linking total dissolved solids to protect aquatic life. Journal of Soil & Water Conservation 62: 57–61.
Bond, N. R. & P. S. Lake, 2003. Local habitat restoration in streams: constraints on the effectiveness of restoration for stream biota. Ecological Management and Restoration 4: 193–198.
Braun, E. L., 1950. Deciduous Forests of North America. Hafner, New York, NY.
Bray, J. R. & J. T. Curtis, 1957. An ordination of the upland forest communities in southern Wisconsin. Ecological Monographs 27: 325–349.
Brinkman, S. F. & W. D. Johnston, 2008. Acute toxicity of aqueous copper, cadium, and zinc to the mayfly Rhithrogena hageni. Achives of Environmental Contamination and Toxicology 54: 466–472.
Brittain, J. E. & S. J. Saltveit, 1989. Review of effect of river regulation on mayflies (Ephemeroptera). Regulated Rivers Research and Management 3: 191–204.
Bryant, G., S. McPhilliamy & H. Childers, 2002. A Survey of the Water Quality of Streams in the Primary Region of Mountaintop/valley Fill Coal Mining. Mountaintop Mining/valley Fill Programmatic Environmental Impact Statement. Region 3, US Environmental Protection Agency, Wheeling, WV.
Buchwalter, D. B. & S. N. Luoma, 2005. Differences in dissolved cadmium and zinc uptake among stream insects: mechanistic explanations. Environmental Science and Technology 39: 498–504.
Buchwalter, D. B., D. J. Cain, W. H. Clements & S. N. Luoma, 2007. Using biodynamic models to reconcile differences between laboratory toxicity tests and field biomonitoring with aquatic insects. Environmental Science and Technology 41: 4821–4828.
Buss, D. F. & F. F. Salles, 2007. Using Baetidae species as biological indicators of environmental degradation in a Brazilian River basin. Environmental Monitoring and Assessment 130: 365–372.
Chadwick, M. A., H. Hunter, J. M. Feminella & R. P. Henry, 2002. Salt and water balance in Hexagenia limbata (Ephemeroptera: Ephemeridae) when exposed to brackish water. Florida Entomologist 85: 650–651.
Chambers, D. B. & T. Messer, 2000. Benthic Invertebrate Communities and Their Responses to Selected Environmental Factors in the Kanawha River Basin, West Virginia, Virginia, and North Carolina. Water-Resources Investigations Report 01-4021. US Geological Survey, Charleston, WV.
Clements, W. H., 1994. Benthic invertebrate community responses to heavy metals in the upper Arkansas River Basin, Colorado. Journal of the North American Benthological Society 19: 30–44.
Clements, W. H., 2004. Small-scale experiments support causal relationships between metal contamination and macroinvertebrate community response. Ecological Applications 14: 954–967.
Clements, W. H., D. S. Cherry & J. H. Van Hassel, 1992. Assessment of the impact of heavy metals on benthic communities at the Clinch River (Virginia): evaluation of an index of community sensitivity. Canadian Journal of Fisheries and Aquatic Sciences 49:1686–169.
Courtney, L. A. & W. H. Clements, 2000. Sensitivity to acidic pH in benthic invertebrate assemblages with different histories of metal exposure. Journal of the North American Benthological Society 19: 112–127.
Cuffney, T. F., H. Zappia, E. M. P. Giddings & J. F. Coles, 2005. Effects of urbanization on benthic macroinvertebrate assemblages in contrasting environmental settings: Boston, Massachusetts; Birmingham, Alabama; and Salt Lake City, Utah. American Fisheries Society Symposium 47: 361–407.
Goetsch, P. A. & C. G. Palmer, 1996. Salinity tolerances of selected macroinvertebrates of the Sabie River, Kruger National Park, South Africa. Archives of Environmental Contamination and Toxicology 32: 32–41.
Green J., M. Passmore & H. Childers, 2000. A survey of the condition of streams in the primary region of mountaintop mining/valley fill coal mining. Mountaintop Mining/Valley Fill Programmatic Environmental Impact Statement. U.S. Environmental Protection Agency, Region III. Wheeling, WV. http://www.epa.gov/region3/mtntop/eis2003appendices.htm.
Griffith, M. B., E. M. Barrows & S. A. Perry, 1998. Lateral dispersal of adult aquatic insects (Plecoptera, Trichoptera) following emergence from headwater streams in forested Appalachian catchments. Annals of the Entomological Society of America 91: 195–201.
Haefner, J. D. & J. B. Wallace, 1981. Shifts in aquatic insect populations in a first-order Appalachian stream following a decade of old field succession. Canadian Journal of Fisheries and Aquatic Science. 38: 353–359.
Hartman, K. J., M. D. Kaller, J. W. Howell & J. A. Sweka, 2005. How much do valley fills influence headwater streams? Hydrobiologia 532: 91–102.
Hassell, K. L., B. J. Kefford & D. Nugegoda, 2006. Sub-lethal and chronic salinity tolerances of three freshwater insects: Cloeon sp. and Centroptilum sp. (Ephemeroptera: Baetidae) and Chironomus sp. (Diptera: Chironomidae). Journal of Experimental Biology 209: 4024–4032.
Herlihy, A. T., J. L. Stoddard & C. B. Johnson, 1998. The relationship between stream chemistry and watershed land cover data in the Mid-Atlantic region, US. Water, Air, and Soil pollution 105: 377–386.
Hershey, A. E., J. Pastor, B. J. Peterson & G. J. Kling, 1993. Stable isotopes resolve the drift paradox for Baetis mayflies in an arctic river. Ecology 74: 2415–2425.
Hughes, J., 2007. Constraints on recovery: using molecular methods to study connectivity of aquatic biota in rivers and streams. Freshwater Biology 52: 616–631.
Kennedy, A. J., D. S. Cherry & R. J. Currie, 2003. Field and laboratory assessment of a coal processing effluent in the Leading Creek watershed, Meigs County, Ohio. Archives Environmental Contamination and Toxicology 44: 324–331.
Kentucky Department for Environmental Protection (KYDEP), 2007. Kentucky Administrative Regulations, 401 KAR 5:031 Section 4. Kentucky Department for Environmental Protection, Frankfort, Kentucky. http://www.lrc.ky.gov/kar/401/010/031.htm.
Kentucky Department for Environmental Protection (KYDEP), 2008. Methods for Assessing Biological Integrity of Surface Waters in Kentucky. Kentucky Energy and Environment Cabinet. http://www.water.ky.gov/sw/swmonitor/sop/.
Kondratieff, B. C. (coordinator), 2000. Mayflies of the United States. Northern Prairie Wildlife Research Center Online, Jamestown, ND. Accessed January 7, 2009. http://www.npwrc.usgs.gov/resource/distr/insects/mfly/index.htm (Version 12DEC2003).
Kreutzweiser, D. P., S. S. Capell & K. P. Good, 2005. Macroinvertebrate community response to selection logging and upland areas of headwater catchments in a northern hardwood forest. Journal of the North American Benthological Society 24: 208–222.
Lemly, A. D., 1998. Bacterial growth on stream insects: potential for use in bioassessment. Journal of the North American Benthological Society 17(2): 228–238.
Lemly, A. D., 2000. Using bacterial growth on insects to assess nutrient impacts in streams. Environmental Monitoring and Assessment 63: 431–446.
Masters, Z., I. Petersen, A. G. Hildrew & S. J. Ormerod, 2006. Insect dispersal does not limit the biological recovery of streams from acidification. Aquatic Conservation: Marine and Freshwater Ecosystems 17: 375–383.
McClurg, S. E., J. T. Petty, P. M. Mazik & J. L. Clayton, 2007. Stream ecosystem response to limestone treatment in acid impacted watersheds of the Allegheny Plateau. Ecological Applications 17: 1087–1104.
McCune, B. & J. B. Grace, 2002. Analysis of Ecological Communities. MjM Software Design, Gleneden Beach, OR.
McCune, B. & M. J. Mefford, 1999. Multivariate Analysis of Ecological Data. Version 4.25. MjM Software, Gleneden Beach, OR.
Merricks, T. C., D. S. Cherry, C. E. Zipper, R. J. Currie & T. W. Valenti, 2007. Coal mine hollow fill and settling pond influences on headwater streams in southern West Virginia, USA. Environmental Monitoring and Assessment 129: 359–378.
Moog, O., E. Bauernfeind & P. Weicshelbaumer, 1997. Use of Ephemeroptera as saprobic indicators in Austria. In Landolt, P. & M. Sartori (eds), Ephemroptera and Plecoptera: Biology-Ecology-Systematics. MTL, Fribourg: 254–260.
Morse J. C., B. P. Stark, W. P. McCafferty & K. J. Tennessen, 1997. Southern Appalachian and other southeastern streams at risk: implications for mayflies, dragonflies, stoneflies, and caddisflies. In Benz, G. W. & D. E. Collins (eds), Aquatic Fauna in Peril: The Southeastern Perspective. Special Publication 1, Southeastern Aquatic Research Institute. Lenz Design and Communications, Decatur, GA: 17–42, 554.
Mount, D. R., D. D. Gulley, J. R. Hoickett, T. D. Garrison & J. M. Evans, 1997. Statistical models to predict the toxicity of major ions to Ceriodaphnia dubia, Daphnia magna, and Pimephales promelas (fathead minnows). Environmental Toxicology and Chemistry 16(10): 2009–2019.
NOAA, 2009. http://www.ncdc.noaa.gov/oa/climate/research/drought/palmer-maps/.
Parker, C. R., O. S. Flint, L. M. Jacobs, B. C. Kondratieff, W. P. McCafferty & J. C. Morse, 2007. Ephemeroptera, Plecoptera, Megaloptera, and Trichoptera of Great Smoky Mountains National Park. Southeastern Naturalist Special Issue 1: 159–174.
Paul, M. J. & J. L. Meyer, 2001. Streams in the urban landscape. Annual Review of Ecology and Sytematics 32: 333–365.
Petersen, I., Z. Masters, A. G. Hildrew & S. J. Omerod, 2004. Dispersal of adult aquatic insects in catchment of differing land use. Journal of Applied Ecology 41: 934–950.
Pond G. J. & S. E. McMurray, 2002. A macroinvertebrate bioassessment index for headwater streams in the eastern coalfield region, Kentucky. Kentucky Department for Environmental Protection, Division of Water, Frankfort, KY. http://www.water.ky.gov/NR/rdonlyres/4CA8D7C4-309B-4175-ACC4-1CBDDDF73798/0/EKyMBI.pdf.
Pond G. J., S. C. Call, J. F. Brumley & M. C. Compton, 2003. The Kentucky macroinvertebrate bioassessment index: derivation of regional narrative criteria for headwater and wadeable streams. Kentucky Department for Environmental Protection, Division of Water, Frankfort, KY. http://www.water.ky.gov/NR/rdonlyres/7F189804-4322-4C3E-B267-5A58E48AAD3F/0/Statewide_MBI.pdf.
Pond, G. J., M. E. Passmore, F. A. Borsuk, L. Reynolds & C. J. Rose, 2008. Downstream effects of mountaintop coal mining: comparing biological conditions using genus- and family-level bioassessment tools. Journal of the North American Benthological Society 27: 717–737.
R Development Core Team, 2009. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. http://www.R-project.org.
Randolph R. P. & W. P. McCafferty, 1998. Diversity and distribution of the mayflies (Ephemeroptera) of Illinois, Indiana, Kentucky, Michigan, Ohio, and Wisconsin. Ohio Biological Survey Bulletin (new series) 13(1): 188 pp.
Reice, S. R., 1985. Experimental disturbance and the maintenance of species diversity in a stream community. Oecologia 67: 90–97.
Slonecker, E. T. & M. J. Benger, 2002. Remote sensing and mountaintop mining. Remote Sensing Reviews 20: 293–322.
Smith, R. F. & W. O. Lamp, 2008. Comparison of insect communities between adjacent headwater and main-stem streams in urban and rural watersheds. Journal of the North American Benthological Society 27: 161–175.
Snyder, C. D. & Z. B. Johnson, 2006. Macroinvertebrate assemblage recovery following a catastrophic flood and debris flows in an Appalachian mountain stream. Journal of the North American Benthological Society 25: 825–840.
Soucek, D. J., 2001. Integrative bioassessment of acid mine drainage impacts on the Upper Powell River watershed, Southwestern Virginia. PhD dissertation. Virginia Polytechnic Institute and State University, Blacksburg, Virginia.
Stone, M. K. & J. B. Wallace, 1998. Long-term recovery of a mountain stream from clear-cut logging: the effects of forest succession on benthic community structure. Freshwater Biology 39: 151–169.
U.S. Environmental Protection Agency, 2005. Mountaintop Mining/Valley Fills in Appalachia, Final Programmatic Environmental Impact Statement. http://www.epa.gov/region3/mtntop/index.htm.
Wallace, J. B., 1990. Recovery of lotic macroinvertebrate communities from disturbance. Environmental Management 14: 605–620.
Wallace, J. B. & M. E. Gurtz, 1986. Response of Baetis mayflies (Ephemeroptera) to catchment logging. American Midland Naturalist 115: 25–41.
Wallace, J. B., M. E. Gurtz & F. Smith-Cuffney, 1988. Long-term comparison of insect abundances in disturbed and undisturbed Appalachian headwater streams. Verhandlungen der Internationalen Vereinigung fur Theoretische und Angewandte Limnologie 23: 1224–1231.
Weijters, M. J., J. H. Janse, R. Alemade & J. T. A. Verhoeven, 2009. Quantifying the effect of catchment land use and water nutrient concentrations on freshwater river and stream biodiversity. Aquatic Conservation: Marine and Freshwater Ecosystems 19: 104–112.
Wellnitz, K. A., S. Grief & S. P. Sheldon, 1994. Response of macroinvertebrates to blooms of iron-depositing bacteria. Hydrobiologia 281: 1–17.
Wickham, J. D., K. H. Riitters, T. G. Wade, M. Coan & C. Homer, 2007. The effect of Appalachian mountaintop mining on interior forest. Landscape Ecology 22: 179–187.
Woods A. J., J. M. Omernik, W. H. Martin, G. J. Pond, W. M. Andrews, S. M. Call, J. A. Comstock & D. D. Taylor, 2002. Ecoregions of Kentucky (2 sided color poster with map, descriptive text, summary tables, and photographs). US Geological Survey (map scale 1:1,000,000), Reston, VA.
Wunsch, D. R., J. S. Dinger, P. B. Taylor, D. I. Carey & C. D. R. Graham, 1996. Hydrogeology, Hydrogeochemistry, and Spoil Settlement at a Large Mine-spoil Area in Eastern Kentucky: Star Fire Tract. Report of Investigations 10, Series XI. Kentucky Geological Survey, University of Kentucky, Lexington, KY.
Acknowledgements
Special thanks for field and laboratory assistance go to present and former KYDEP biologists S. McMurray, M. Compton, D. Peake, R. Pierce, M. Vogel, R. Payne, C. Schneider, J. Brumley, E. Eisiminger, and S. Call. Thanks also to U.S. EPA’s L. Yuan for statistical support and to U.S. EPA’s M. Passmore, L. Alexander, J. Forren, and K. Krock, and KYDEP’s B. Marbert, for providing helpful comments on earlier versions of the manuscript. The final manuscript was improved by R. Bailey and 2 anonymous referees. Views and opinions are those of the author and this manuscript does not necessarily reflect the official views of either the U.S. Environmental Protection Agency or the KY Department for Environmental Protection.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Robert Bailey
Rights and permissions
About this article
Cite this article
Pond, G.J. Patterns of Ephemeroptera taxa loss in Appalachian headwater streams (Kentucky, USA). Hydrobiologia 641, 185–201 (2010). https://doi.org/10.1007/s10750-009-0081-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-009-0081-6