
Abstract
Almost 95% of the entire population of the Siberian crane (Grus leucogeranus) winter in Poyang Lake, China, where they forage on the tubers of the submerged aquatic macrophyte Vallisneria spiralis. The Three Gorges Dam on the Yangtze River may possibly affect this food source of the Siberian crane by affecting the light intensity reaching the top of the V. spiralis canopy. In this study, the photosynthetically active radiation at the top of the V. spiralis canopy (PARtc) in Lake Dahuchi was modeled from 1998 to 2006, and the potential impacts of changes in water level and turbidity on the underwater light climate of V. spiralis were analyzed. PARtc was calculated from incident irradiance while the losses due to reflection at the water surface, absorption, and scattering within the water column were taken into consideration. The results indicated significant differences in PARtc between years. Six years of water level and Secchi disk depth records revealed a seasonal switching of the lake from a turbid state at low water levels in autumn, winter, and spring to a clear state at high water levels during the monsoon in summer. The highest PARtc occurred at intermediate water levels, which were reached when the Yangtze River forces Lake Dahuchi out of its turbid state in early summer and the water becomes clear. The intended operation of the Three Gorges Dam, which will increase water levels in May and June, may advance the moment when Lake Dahuchi switches from turbid to clear. We suggest that this might increase production of V. spiralis and possibly improve the food habitat conditions for wintering Siberian crane in Poyang Lake.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Endemic species are vulnerable to extinction when environmental change affects their habitat. Thus, areas with high diversity of endemic species deserve attention in case of environmental change. Poyang Lake, a lake system connects to the Yangtze River of China, hosts significant proportions of the Yangtze finless porpoise (Neophocaena phocaenoides asiaeorientalis Pilleri and Gihr), swan goose (Anser cygnoides L.), and almost 95% of the entire world population of the endangered Siberian crane (Grus leucogeranus Pallas). The Siberian crane winter at this lake and forage on the tubers of the submerged aquatic macrophyte, Vallisneria spiralis L. (Wu & Ji, 2002; Li et al., 2005).
The environmental conditions of Poyang Lake are rapidly changing due to economic development and hydro-engineering projects in the watershed to which it is associated. For example, dredging increases the water turbidity in the northern Poyang Lake (Wu et al., 2007). The environmental conditions might also be influenced by operation of the Three Gorges Dam in the Yangtze River, which is scheduled for completion by 2009 (Wang, 2002). Several authors suggested that the Three Gorges Dam might affect the habitat of the Siberian crane (Liu & Xu, 1994; Jiang & Huang, 1997; Kanai et al., 2002; Wu et al., 2007), without however explaining in detail as to how this will happen. We suggest that the Three Gorges Dam might affect the habitat of the Siberian crane as follows: (1) the Three Gorges Dam may affect the hydrology of Poyang Lake, (2) the changed hydrology may influence the tuber production of V. spiralis in Poyang Lake, and (3) the changed tuber production may affect the foraging by the wintering Siberian crane. So far, however, there is no direct evidence to support this expectation, as little research has been reported on the ecology of the tuber-feeder food chain of Poyang Lake.
Is there evidence to support the premise that a changed tuber production would affect the foraging of tuber-feeders like the Siberian crane? Jonzen et al. (2002), Nolet et al. (2006) and Raymond et al. (2006) reported that tuber density of sago pondweed (Potamogeton pectinatus) affects the foraging of tundra swan (Cygnus columbianus bewickii) in Lake Lauwersmeer in the north of the Netherlands, while Sponberg & Lodge (2005) found that waterfowl in Lake Mattamuskeet, North Carolina, USA, gave up foraging when tuber density of wildcelery (Vallisneria americana) was below a certain threshold.
How would hydrology influence the photosynthesis and tuber production of V. spiralis? The growth and production of submerged vegetation is regulated by light intensity reaching its canopy, water temperature, CO2, and nutrient availability (Best & Boyd, 2001; Best et al., 2001; Van Nes, 2002). Light availability likely controls the growth and production of V. spiralis as it is highly variable in the waters with fluctuating water depth and clarity of Poyang Lake. The light response of V. spiralis has not been studied, but light compensation points of 20–25 and 10 mol m−2 s−1, respectively, were reported for the related V. americana (Madsen et al., 1991; Blanch et al., 1998). This corresponds to 1–2% of surface irradiance during sunshine, and agrees with the observation that V. americana maintains photosynthesis above 0.5% of surface irradiance (Korschgen & Green, 1988). Tuber production requires a higher light intensity. Kimber et al. (1995) reported for V. americana a threshold of 5–9% of surface irradiance for tuber production under temperate climatological conditions, which forms the lower limit of the 9% surface irradiance threshold cited for V. americana occurrence in temperate North-Florida (Dobberfuhl, 2007).
How would the hydrology of the Yangtze River affect the hydrological conditions in Poyang Lake, and, thus, indirectly, the tuber production of V. spiralis? The Yangtze River with five confluent rivers together controls the water level in Poyang Lake (Min, 1995; Xu et al., 2001; Shankman et al., 2006). The Siberian cranes forage in lakes located at the periphery of Poyang Lake. So far, no analysis has been made to investigate whether the Yangtze River influences the water level and clarity of these peripheral lakes.
A crucial question is whether the Three Gorges Dam will affect the hydrology in Poyang Lake. Addressing this question requires insight into the consequence of operating the Three Gorges Dam and its impact on the dynamics of the Yangtze River. Such an analysis has recently been presented by Wang et al. (2005). The Three Gorges Dam will be operated for flood control, power generation, and navigation. The water level of the reservoir will alternate between 175 and 145 m in winter and summer, respectively. The discharge will be increased from January to April for power generation, while from May to June the water level of the reservoir will be further lowered to create capacity for flood control during the monsoon season. The discharge will be reduced from October to November to recharge the reservoir. Above we argued that the water level change of the Yangtze River might affect the production of V. spiralis while influencing the light regime. So far, it is not understood how the reported operation of the Three Gorges Dam and the induced water level change in the Yangtze River might influence the light regime of V. spiralis in Poyang Lake.
This paper presents a model simulating the photosynthetically active radiation (PAR) reaching the top of V. spiralis canopy and discusses how the changes in water level of the Yangtze River induced by operation of the Three Gorges Dam may affect the PAR in Lake Dahuchi.
Materials and methods
Study area
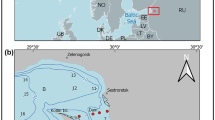
Poyang Lake, the largest freshwater lake in China, is located south of the Yangtze River between 115°47′–116°45′ E, 28°22′–29°45′ N (Fig. 1). It receives runoff from five rivers (Raohe, Xinjiang, Fuhe, Ganjiang, and Xiushui), while a channel leading to the Yangtze River serves as the only outlet of the lake. The Poyang Lake national nature reserve (115°55′–116º03′ E, 29°05′–29°15′ N) in the northwest (Fig. 1) was established in 1988 to conserve the Siberian crane (Wu & Ji, 2002; Li et al., 2005). The current study was carried out in Lake Dahuchi within the nature reserve, which connects to Poyang Lake during high water in summer, while separates from Poyang Lake when water levels are low in spring, autumn, and winter.

Location of Poyang Lake national nature reserve, Lake Dahuchi, and Dahuchi Conservation Station ( ) where daily photosynthetically active radiation (PAR) was measured
) where daily photosynthetically active radiation (PAR) was measured
Water level and Secchi disk depth
Lake Dahuchi, a shallow oxbow lake with its deepest part at 12 mFootnote 1 is surrounded by creek banks at elevations of 14 to 15 m. Its water level is controlled from September to April by a sluice gate, connecting Lake Dahuchi with the Xiushui River. During high water levels in summer, Lake Dahuchi connects with Poyang Lake, and its water level is, thus, affected by the Yangtze River and the five confluent rivers. To evaluate how the water levels of the Xiushui River, Poyang Lake (Ganjiang station), and the Yangtze River (Hukou station) affect the water level in Lake Dahuchi, we explored tentative relationships between gage data.
Water level and Secchi disk depth have been measured in Lake Dahuchi by the Bureau of Jiangxi Poyang Lake National Nature Reserve and by the International Crane Foundation since 1999. Daily water level was recorded using a water level gauge. Secchi disk depth was measured using a standard 20 cm Secchi disk at five locations at weekly intervals from April until October. Daily Secchi disk depths were derived through linear interpolation from the recorded values.
The water level and Secchi disk depth records in Lake Dahuchi were incomplete for 1998–2000 and 2002, and, therefore, the water level data from the nearby station in the Xiushi River were used to replace the missing water level data for Lake Dahuchi. A non-linear relation between Secchi disk depth (SDD) and water level (WL) was observed while exploring the recorded water level and clarity data, which was described by a Huisman–Olff–Fresco (HOF) model (Eq. 1) (Huisman et al., 1993):
where c 1, c 2, and c 3 are constants. Equation 1 was then used to predict Secchi disk depth to complete the gaps in the Secchi disk time series from 1998 to 2006.
Modeling photosynthetically active radiation (PAR)
The PAR reaching the top of V. spiralis canopy was estimated as follows. PAR at the water surface (PAR+0) was calculated from the global solar radiation recorded at Nanchang meteorological station (approximately 55 km from Lake Dahuchi), assuming that PAR accounts for 45% of the total global solar radiation (based on values of 50, 38, and 44% according to Doorenbos & Pruitt (1984), Zhang & Qin (2002), and Moon (cited by Zhang et al., 2006)). PAR at the top of the Dahuchi Conservation Station recorded from July 6 to August 12 and from September 6 to November 10, 2006 using an LI-190SA quantum sensor and an LI-1400 data logger (http://www.licor.com) served as a validation data set.
Subsurface PAR (PAR−0) was calculated with Eq. 2, assuming a reflectance rate (k r) of 4% according to Campbell & Aarup (1989) and Joshi (2005):
Then the PAR at water depth d (PAR d ) was calculated according to Lambert–Beer’s law (Eq. 3):
where the attenuation coefficient, k PAR (m−1), was derived from Secchi disk depth (SDD) records according to Eq. 4 (Poole & Atkins, 1929):
where a is a constant, which may vary from case to case for different water types (Kirk, 1994; Scheffer, 1998). The value of K par was determined from measurements of underwater PAR at various depths at 32 locations in Lake Dahuchi on August 12 and October 16, 2005 using an LI-192SA quantum sensor and an LI-250A light meter (http://www.licor.com). At the same time, the Secchi disk depth at each location was measured using a standard 20 cm Secchi disk. Regressing the recorded K par against SDD (Fig. 2) according to Eq. 4 (R 2 = 0.98, F = 1836.495, n = 32, P < 0.005) provided an estimate of the constant of a = 1.133 with a standard error of 0.026, or 2% of the estimate.

Relation between the attenuation coefficient of photosynthetically active radiation (k PAR) and Secchi disk depth
Finally, PAR at water depth d and on the top of V. spiralis canopy (PARtc) was calculated according to Eqs. 5 and 6, respectively, which integrate Eqs. 2, 3, and 4:
where e w and e b are water level and lake bottom elevation, respectively, and l is the height of the V. spiralis, which increases with the number of days since the first day of May (nd) according to a regression equation (Eq. 7) published by Wu & Ji (2002).
Modeling PAR variation against water level
PARtc depends on the lake bottom elevation at which V. spiralis was rooted. In this study, the Eqs. 6 and 7 were combined to simulate the temporal variation of PARtc for V. spiralis rooted at elevations of 12 and 12.8 m in Lake Dahuchi, respectively, which correspond to the lowest and highest locations of V. spiralis in this lake. Since V. spiralis forms tubers from July to September (Wu & Ji 2002), ANOVA was used to explore whether PARtc in this period differed between years.
The stepwise relation between water level and water turbidity might induce non-linearity in the relation between the PAR at a selected elevation and water level. Thus, by integrating the Eqs. 1, 2, 3, and 4 into a new Eq. 8, PAR at a selected elevation (PARele) in the lake can be calculated as a function of water level:
Results
Figure 3 shows that water level in Lake Dahuchi, Xiushui River, Poyang Lake (Ganjiang station), and Yangtze River (Huhou station) fluctuates seasonally, with maxima during the monsoon reached between June and September. From mid-September to mid-May, the water levels in Lake Dahuchi remained below 14 m and were independent of those of the Xiushui River, Poyang Lake and the Yangtze River. More pronounced fluctuations of Lake Dahuchi between May and September were correlated with the fluctuations in water levels of the Xiushui River (r = 0.84 and 0.94 in 2004 and 2005), Poyang Lake (r = 0.83 and 0.93 in 2004 and 2005), and the Yangtze River (r = 0.81 and 0.93 in 2004 and 2005). The water levels of the Yangtze River, Poyang Lake, and the Xiushui River receded in September in both years, when the level of Lake Dahuchi started to deviate again from those of the other systems. The Lake Dahuchi water level was controlled and slowly drawn down via a sluice to gradually expose extensive areas of shallow water. This particular water level management prescribed by the authority of the nature reserve enables Siberian crane, which requires shallow water conditions to reach the tubers of V. spiralis, to forage from the end of October onwards.

Water levels (m) in Lake Dahuchi (—), Xiushui River (– –), Poyang Lake at Ganjiang (·····), and the Yangtze River at Hukou (- - -) in 2004 (A) and 2005 (B). Vertical lines mark the onset of water level in Lake Dahuchi being dependent and independent of that of Xiushui River in spring (about May 14) and autumn (about September 22) respectively
Although the water levels of Lake Dahuchi were linearly related to that of the Yangtze River at Hukou when the Yangtze River water level was higher than 14 m (Fig. 4), there was no relationship when the Yangtze River water level at Hukou was less than 14 m. In other words, below 14 m Lake Dahuchi water level appears to be independent of that of the Yangtze River, while it is related above 14 m.

Relation between the water level of Lake Dahuchi and that of the Yangtze River at Hukou from May 14 to September 22 in 2004 and 2005. The solid line represents the 1:1 relation
The daily PAR recorded at Nanchang meteorological station varied seasonally with the inclination of the sun and the variation in atmospheric conditions (Fig. 5A). One-way ANOVA revealed that the PAR did not differ significantly between years (F = 1.728, d.f. = 8, P = 0.08). Daily PAR extrapolated from Nanchang station predicted 88% of the variation of PAR at Dahuchi Conservation Station (F = 745.29, n = 105, P < 0.001). The prediction was unbiased as the intercept and slope of the regression did not differ significantly from zero (a = 0.23, s.e. = 0.26, t = 0.909, d.f. = 104, P = 0.39) and one (b = 1.02, s.e. = 0.04, t = 0.535, d.f. = 104, P = 0.59), respectively.

(A) Estimated daily PAR at Nanchang station, (B) recorded daily water level in Lake Dahuchi (—) or Xiushui River (·····), (C) recorded (·), interpolated (—), or predicted (·····) Secchi disk depth, and (D) and (E) estimated daily PAR (averaged over 2 week periods) reaching the top of canopy of V. spiralis rooted at 12 and 12.8 m. Horizontal lines in (D) and (E) represent a limit of 9% surface radiation during sunshine period (Kimber et al., 1995; Dobberfuhl, 2007), the threshold for tuber production in V. americana
The water levels (Fig. 5B) revealed a seasonal pattern, from high in summer to low in winter. Summer water levels were high in 1998 and 1999 and low in 2001, 2004, and 2006.
The Secchi disk depth in Lake Dahuchi (Fig. 5C) changed from almost or close to zero (turbid) at a water level below 13.5 m to around 1.1 m on average (clear) at a water level of 14.5 m. Water level (WL) explained (Fig. 6) 60% of the variation of Secchi disk depth (SDD) with the HOF model (SDD = 1.125/(1 + e 46.837−3.352*WL, F = 247.990, n = 155, P < 0.005). The low Secchi disk depth at a water level less than 13.5 m (Fig. 6) coincided with the period of lake recession between September–October and April–May of the following year (Fig. 5B). From Figs. 5 and 6, it can be concluded that Lake Dahuchi switches from a turbid state between autumn and spring to a clear state in summer. Figure 6 reveals variation in SDD of an order of magnitude (from 0.2 to 2 m at 14.5 m) at higher water levels. The regression equation, which describes the average response, was used for further modeling to complete the Secchi disk depth record from 1998 to 2006 (called ‘predicted’) in Fig. 5C.

Relation between water turbidity (Secchi disk depth) and water level in Lake Dahuchi (● = measured in the field, □ = estimated from MODIS images (Wu et al., 2008))
Daily PAR (averaged over 2 week periods) reaching the top of the V. spiralis canopy rooted at an elevation of 12 m was generally low in spring and increased in summer and autumn (Fig. 5D), while radiation reaching the top of the V. spiralis canopy rooted at an elevation of 12.8 m was considerably higher (Fig. 5E). One-way ANOVA revealed that the annual PARtc from July to September differed significantly between years for plants rooted at elevations of 12 m (F = 92.07, d.f. = 8, P < 0.001) and 12.8 m (F = 112.19, d.f. = 8, P < 0.001), respectively. PAR levels at 12.8 m over this period remained below the 9% irradiance threshold required for tuber production in the related species V. americana in 1998, 1999, and 2006, while for plants rooted at 12 m, this threshold was exceeded in 2001, 2003, and 2004 only.
The water level in Lake Dahuchi was extremely high from July to September in 1998 and 1999 (Fig. 5B). PARtc was also reduced in both years (Fig. 5D, E), despite the fact that water was very clear in 1999. PARtc remained low as well when water levels were extremely low in 2005 and 2006.
The relationship between water level and water turbidity in Lake Dahuchi (Fig. 6) indicates that an increase in water level might increase rather than decrease the light availability at a certain elevation in the water column. Equation 8 was thus used to calculate the PAR at various elevations as a function of water level (Fig. 7). It turned out that the light intensity increased at any elevation between 12 and 13 m when the water level increased above 13.5 m, and peaked around 14.5–15 m. For submerged vegetation with canopy at an elevation of 13 m, the PAR at a water level of 16 m is similar to that at water level of 13.5 m (about 5% of incident radiation); however, the light intensity of about 17% of incident radiation at a water level of 14.5 m is three times higher. Thus, submerged aquatic vegetation canopies at elevations between 12 and 13 m receive less light than the requirement for tuber formation in the related V. americana at water levels less than 13.5 m. Increasing the water level increases the light intensity to levels surpassing the threshold for tuber production of V. americana.

Percentage of surface irradiance reaching a certain elevation (m above datum) below the water surface as a function of water level. Light compensation point (1% surface irradiance, (A)) and threshold for tuber production (9% surface irradiance, (B)) reported for V. americana indicated by thick lines
Discussion
This study revealed a strong seasonal and inter-annual fluctuation in PAR reaching the top of a V. spiralis canopy. These fluctuations were caused by variations in irradiance reaching the water surface, water level, attenuation within the water column, and height of the V. spiralis vegetation. Irradiance at the water surface varied strongly from day to day, but ANOVA revealed that it did not vary significantly between years. To what extent does the length of leaves influence the year to year variation in PARtc? Aquatic plant species elongate their leaves in response to low light intensities (Bowes et al., 1977; Cooling et al., 2001). The model used here (Eq. 7) does not account for such flexibility in leaf elongation, as it was based on observations from 2 years (1998 and 1999) with extremely high water levels. The predicted inter-annual variation in PAR reaching the top of canopy as shown in Fig. 5 can thus not be attributed to year to year variation in height of the V. spiralis vegetation, as we used the same model to simulate the height for all years. We, thus, argue that the predicted inter-annual fluctuations in PAR reaching the canopy depend on fluctuations in water level and light attenuation within the water column. As light attenuation appeared in turn determined by water level, we conclude that water level exerts an overriding impact on the PAR reaching the canopy of V. spiralis.
We also reported a non-linear relation between water level and PAR at a given elevation in the lake water column. This non-linearity was caused by the sigmoid relation between water level and Secchi disk depth. Water with a high turbidity at a water level less than 13.5 m switched to a greater clarity at a level above 14.5 m. The high turbidity at low water levels (below 13.5 m) is attributed to wave-induced sediment resuspension at the lake periphery, as is common in shallow lakes (Scheffer, 1998). The increased clarity at elevated water levels above 14.5 m can be explained by reduced sediment resuspension since the lake fringe then overlays perennial grassland.
How would the combination of changes in water level and transparency influence the production of V. spiralis? Our results revealed that at water levels below an elevation of 13 m V. spiralis is exposed to light intensities below the compensation point reported for the related V. americana. We presume that this reflects the threshold of V. spiralis, as most submerged aquatic plants have compensation points close to 1% of surface irradiance (e.g., Korschgen 1988; Madsen et al., 1991; Blanch et al., 1998). The model presented here further suggests that water levels surpassing an elevation of 14 m are required to create the light conditions permitting tuber production. This suggestion is based on the assumption that the light requirement of V. spiralis for tuber formation is the same as that of the related V. americana, i.e., 9% of surface irradiance. Indeed we propose to further study the light requirements of V. spiralis regarding photosynthesis and reproduction.
We demonstrated that the water level of Lake Dahuchi tracked that of the Xiushui River, Poyang Lake, and the Yangtze River at high water levels from May to September, which is the growing season of V. spiralis. We, thus, suggest that the levels of the Yangtze River and the inflows from the confluent rivers influence the production of V. spiralis through their impact on water level, clarity, and light regime in Lake Dahuchi. Because the wintering Siberian crane forage on the tuber of V. spiralis, we expect that the water regimes of the Yangtze River and the confluent rivers would be crucial for the productivity of the food source for the wintering Siberian crane.
Wang et al. (2005) reported that the Three Gorges Dam will change the water level of the Yangtze River in May and June. An advancement of the rise of the water level of the Yangtze River during the growth season of V. spiralis may force Lake Dahuchi out of its turbid state earlier and increase the light intensity reaching the canopy and the production of V. spiralis. On the other hand, a water level surpassing 14.5 m elevation would decrease the light intensity reaching the plant canopy. We anticipate, however, that the first possibility would occur more frequently than the second possibility, and conclude that the operation of the Three Gorges Dam is more likely to increase than decrease the light availability and productivity of V. spiralis on the short term. It remains difficult to assess the consequences of this for the Siberian crane, as we do not have accurate models to predict tuber biomass of V. spiralis, and do not know to what extent crane populations are limited by tuber biomass. However, as our results suggest that Three Gorges Dam is likely to improve the light conditions for biomass production we conclude that there is no reason to support the concerns that the Three Gorges Dam would negatively affect the production of food of Siberian crane.
Wang et al. (2005) also reported that the Three Gorges Dam will lower the bed of the Yangtze River due to increased erosion. Water levels in Shashi and Wuhan city are expected to recede by about 3 and 0.75 m, respectively, at a discharge of 30,000 m3 s−1 in the next 40 years. Such receding water levels may negatively impact the light availability and production of V. spiralis, but their impact would be difficult to assess since no prediction of the magnitude of such a water level recession on the Poyang Lake system exists.
The main finding of this research is the new insight that optimal light conditions for submerged vegetation are attained when water levels are pushed up by the Yangtze and other rivers. We suggest that this insight is crucial when aiming to predict the response of tuber forming macrophytes to hydrological change. In this respect it is interesting to note that hydrology will not only be affected by the Three Gorges Dam. A range of other large scale planning or ongoing engineering projects could influence the water levels of the Yangtze River and the confluent rivers of the Poyang Lake system as well. The South-to-North Water Transfer Project will reduce the discharge, and, thus, reduce the water level of the middle and lower Yangtze River (Cai et al., 2002). The hydrology of Poyang Lake will also be affected by reservoirs in its confluents (Song & Jin, 2001). Water levels will be strongly influenced by the proposed Poyang Lake Control Project which aims to manage the lake level for multiple purposes (Xiong & Hu, 2003). All these hydro-engineering projects might change the hydrology of Lake Dahuchi. We suggest that conservation of these wetlands and their fauna should be based on sound insights in their hydro-ecology.
Notes
All elevations and water levels refer to the Chinese National Vertical Datum of 1985.
References
Best, E. P. H. & W. A. Boyd, 2001. A Simulation Model for Growth of the Submersed Aquatic Macrophyte American Wildcelery (Vallisneria americana Michx.). ERDC/EL TR-01-5, U.S. Army Engineer Research and Development Center, Vicksburg, MS.
Best, E. P. H., C. P. Buzzelli, S. M. Bartell, R. L. Wetzel, W. A. Boyd, R. D. Doyle & K. R. Campbell, 2001. Modeling submersed macrophyte growth in relation to underwater light climate: modeling approaches and application potential. Hydrobiologia 444: 43–70.
Blanch, S. J., G. G. Ganf & K. F. Walker, 1998. Growth and recruitment in Vallisneria americana as related to average irradiance in the water column. Aquatic Botany 61: 181–205.
Bowes, G., T. K. Van, L. A. Garrard & W. T. Haller, 1977. Adaptation to low light levels by Hydrilla. Journal of Aquatic Plant Management 15: 32–35.
Cai, S., Y. Du & Y. Zeng, 2002. Main problems of the water and soil environment in the middle and lower reaches of Yangtze River and possible countermeasures. Resources and Environment in the Yangtze Basin 11: 564–568.
Campbell, J. W. & T. Aarup, 1989. Photosynthetically available radiation at high latitudes. Limnology and Oceanography 34: 1490–1499.
Cooling, M. P., G. G. Ganf & K. F. Walker, 2001. Leaf recruitment and elongation: an adaptive response to flooding in Villarsia reniformis. Aquatic Botany 70: 281–294.
Dobberfuhl, D. R., 2007. Light limiting thresholds for submerged aquatic vegetation in a blackwater river. Aquatic Botany 86: 346–352.
Doorenbos, J. & W. O. Pruitt, 1984. Guidelines for Predicting Crop Water Requirements. FAO Irrigation and Drainage Paper 24. FAO, Rome.
Huisman, J., H. Olff & L. F. M. Fresco, 1993. A hierarchical set of models for species response analysis. Journal of Vegetation Science 4: 37–46.
Jiang, J. & Q. Huang, 1997. A study of the impact of the three gorges project on the water level of Poyang Lake (in Chinese). Journal of Nature Resources 12: 219–224.
Jonzen, N., B. A. Nolet, L. Santamaria & M. G. E. Svensson, 2002. Seasonal herbivory and mortality compensation in a swan-pondweed system. Ecological Modelling 147: 209–219.
Joshi, S., 2005. Feature article: underwater light field and its comparison to metal halide lighting. Advanced Aquarist’s Online Magazine 8.
Kanai, Y., M. Ueta, N. Germogenov, M. Nagendran, N. Mita & H. Higuchi, 2002. Migration routes and important resting areas of Siberian cranes (Grus Leucogeanus) between northeastern Siberia and China as revealed by satellite tracking. Biological Conservation 106: 339–346.
Kimber, A., J. L. Owens & W. G. Crumpton, 1995. Light availability and growth of wildcelery (Vallisneria americana) in upper Mississippi River backwaters. Regulated Rivers: Research & Management 11: 167–174.
Kirk, J. T. O., 1994. Light and Photosynthesis in Aquatic Ecosystems. Cambridge University Press, New York, NY, USA.
Korschgen, C. E. & W. L. Green, 1988. American wildcelery (Vallisneria americana): ecological considerations for restoration. U.S. Fish and Wildlife Service, Fish and Wildlife Technical Report: 24.
Li, F., W. Ji, N. Zeng, J. Wu, X. Wu, W. Yi, Z. Huang, F. Zhou, J. Barzen & J. Harris, 2005. Aerial survey of Siberian cranes in the Poyang Lake Basin (in Chinese). In Wang, Q. & F. Li (eds), Crane Research in China. Crane and Waterbird Specialists Group of Chinese Ornithological Society, International Crane Foundation, Yunnan Educational Publishing House, China: 58–65.
Liu, Y. & Y. Xu, 1994. The study of influence and countermeasure of Sanxia Project to Poyang’s migratory birds reserve (in Chinese). Journal of Jiangxi Normal University 18: 375–380.
Madsen, J. D., C. F. Hartleb & C. W. Boylen, 1991. Photosynthetic characteristics of Myriophyllum spicatum and six submersed aquatic macrophyte species native to Lake George, New York. Freshwater Biology 26: 233–240.
Min, Q., 1995. On the regularities of water level fluctuations in Poyang Lake (in Chinese). Journal of Lake Sciences 7: 281–288.
Nolet, B. A., V. N. Fuld & M. E. C. van Rijswijk, 2006. Foraging costs and accessibility as determinants of giving-up densities in a swan-pondweed system. Oikos 112: 353–362.
Poole, H. H. & W. R. G. Atkins, 1929. Photo-electric measurements of submarine illumination throughout the year. Journal of the Marine Biological Association of the United Kingdom 16: 297–324.
Raymond, H. G. K., B. A. Nolet & D. Bankert, 2006. Movement of foraging tundra swans explained by spatial pattern in cryptic food densities. Ecology 87: 2244–2254.
Scheffer, M., 1998. Ecology of Shallow Lakes. Chapman & Hall, London: 357.
Shankman, D., B. D. Keim & J. Song, 2006. Flood frequency in China’s Poyang Lake region: trends and teleconnections. International Journal of Climatology 26: 1255–1266.
Song, L. & L. Jin, 2001. Advances of the study on flood control and disaster reduction Poyang Lake (in Chinese). Environment and Exploitation 16: 4–6.
Sponberg, A. F. & D. M. Lodge, 2005. Seasonal belowground herbivory and a density refuge from waterfowl herbivory for Vallisneria americana. Ecology 86: 2127–2134.
Van Nes, E. H., 2002. Controlling Complexity in Individual-based Models of Aquatic Vegetation and Fish Communities. Wageningen University, Wageningen.
Wang, J., 2002. Three Borges Project: the largest water conservancy project in the world. Public Administration and Development 22: 369–375.
Wang, Z., J. H. W. Lee & D. Cheng, 2005. Impacts of the TGP project on the Yangtze River ecology and management strategies. International Journal of River Basin Management 3: 237–246.
Wu, Y. & W. Ji, 2002. Study on Jiangxi Poyang Lake National Nature Reserve. Forest Publishing House, Beijing: 231.
Wu, G., J. De Leeuw, A. K. Skidmore, H. H. T. Prins & Y. Liu, 2007. Concurrent monitoring of vessels and water turbidity enhances the strength of evidence in remotely sensed dredging impact assessment. Water Research 41: 3271–3280.
Wu, G., J. De Leeuw, A. K. Skidmore, H. H. T. Prins & Y. Liu, 2008. Comparison of MODIS and Landsat TM5 images for mapping tempo-spatial dynamics of Secchi disk depths in Poyang Lake national nature reserve, China. International Journal of Remote Sensing 29: 2183–2198.
Xiong, X. & X. Hu, 2003. Effects of Poyang Lake Control Project on the wetland ecosystem in Poyang Lake region. Journal of Jiangxi Normal University 27: 89–93.
Xu, D., M. Xiong & J. Zhang, 2001. Analysis on hydrologic characteristics of Poyang Lake. Yangtze River 32: 21–27.
Zhang, Y. & B. Qin, 2002. The basic characteristic and climatological calculation of the photosynthetically available radiation in Taihu region (in Chinese). Acta Energiae Solaris Sinica 23: 1–6.
Zhang, Y., B. Qin, G. Zhu, G. Gao, L. Luo & W. Chen, 2006. Effect of sediment resuspension on underwater light field in shallow lakes in the middle and lower reaches of the Yangtze River: a case study in Longgan Lake and Taihu Lake. Science in China Series D: Earth Sciences 49: 114–125.
Acknowledgments
This study was supported by Netherlands Foundation for the Advancement of Tropical Research (WOTRO) (Grant: WB 84-550), the International Institute for Geo-information Science and Earth Observation (ITC), the Netherlands, the School of Resource and Environmental Science, Wuhan University, China, the Bureau of Jiangxi Poyang Lake National Nature Reserve, China, and the International Crane Foundation (ICF).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Luigi Naselli-Flores
Rights and permissions
About this article
Cite this article
Wu, G., de Leeuw, J., Skidmore, A.K. et al. Will the Three Gorges Dam affect the underwater light climate of Vallisneria spiralis L. and food habitat of Siberian crane in Poyang Lake?. Hydrobiologia 623, 213–222 (2009). https://doi.org/10.1007/s10750-008-9659-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-008-9659-7