
Abstract
Aerial seed banks are potentially the main source of sexual recruitment for woody wetland plants. Whilst the importance of soil seed banks for the persistence and recruitment of wetland plants has been examined in many studies, the role of aerial seed banks has been largely neglected. We used seed traps and the seedling emergence technique to quantify the seed rain from aerial seed banks of the Swamp Paperbark Melaleuca ericifolia Sm. (Myrtaceae) in Dowd Morass, a Ramsar-listed, brackish-water wetland in south-eastern Australia. Nine plant species germinated from material collected in seed traps over 2004–2005, but emergents were dominated (80–97%) by M. ericifolia. The mean number of M. ericifolia emergents ranged from <1 to <100 seedlings m−2 day−1, and showed a peak in the summer–autumn period. Regression analysis showed a significant negative correlation (r 2 = 0.738) between the number of M. ericifolia emergents and water depth. Water depth and salinity were negatively correlated (r 2 = 0.819), and increases in the number of M. ericifolia emergents as water levels fell were also associated with high salinities. Increasing air temperature and vapour pressure deficit also stimulated seed release during periods of drying. This study is one of the first to demonstrate the importance of aerial seed banks for sexual recruitment in woody wetland plants and the release of seed in relation to environmental factors. Aerial seed banks warrant consideration alongside soil seed banks for the establishment and long-term survival of woody plants in wetlands.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Aerial seed banks are potentially the most important source of propagules for sexual regeneration in woody wetland plants, given that the seeds of woody wetland plants are often under-represented in the seed banks of wetland sediments (Middleton, 2003; Capon & Brock, 2006) and that aerial seed banks have been demonstrated to play a critical role in sexual recruitment of many taxa of woody plants in terrestrial forests (e.g. Schwilk & Ackerly, 2001; Goubitz et al., 2003; Tapias et al., 2004; Barrett et al., 2005; Russell-Smith & Setterfield, 2006).
The role of seed banks in maintaining floristic diversity in freshwater wetlands has been examined in a very large number of studies (e.g. van der Valk & Davis, 1978; Parker & Leck, 1985; Britton & Brock, 1994; Baldwin et al., 1996; Abernethy & Willby, 1999; Boedeltje et al., 2003; Liu et al., 2006). These studies have tended to emphasise two key ecological processes—the production and dispersal of seeds, and the storage of seeds in soil seed banks—as critical factors in explaining the distribution and regeneration capacity of wetland plants, particularly in response to disturbances such as changes in water level (Brock & Britton, 1995; Capon & Brock, 2006). A significant limitation of most studies to date is that they have concentrated overwhelmingly on soil-based seed banks. Aerial seed banks are an alternative reservoir for seeds, and are a result of mature seed being held in the canopy and progressively released over a period of time, often in response to environmental disturbances such as fire (Goubitz et al., 2003; Tapias et al., 2004).
In contrast to the extensive knowledge of soil seed banks in wetlands and aerial seed banks in terrestrial plants, very few studies have examined the role of aerial seed banks and factors affecting seed rain in the ecology of the woody vegetation of wetlands. The few notable studies include those of Schneider & Sharitz (1988) on Taxodium distichum (L.) Rich and Nyssa aquatica L., and Middleton (2000), also on T. distichum. Middleton (2000) found that seed release and germination in T. distichum was severely restricted in space and time, and recruitment occurred mostly where water levels were drawn down during the growing season. A small number of other papers have dealt with aerial seed banks of the genus Melaleuca (Myrtaceae); much of this research has occurred in southern USA where Melaleuca quinquenervia (Cav.) Blake is a widely distributed and invasive weed. These studies have shown that M. quinquenervia constantly releases a small amount of seed, and that a larger pulse of seed release may be initiated by fire, frost, herbicide or wind damage (e.g. Woodall, 1982; Rayamajhi et al., 2002). Although there is some evidence for the release of seed from aerial seed banks in response to hydrological shifts in other riparian and wetland species in Myrtaceae (e.g. Denton & Ganf, 1994; Pettit & Froend, 2001), to date no study has unravelled the environmental factors that control seed release.
In the present paper, we examine seed rain in Melaleuca ericifolia Sm. (Swamp Paperbark), a plant that is often the dominant canopy species in coastal (freshwater and estuarine) swamps across southern and eastern Australia, from southern Tasmania through to southern Queensland. Melaleuca spp. seeds are produced and stored in closed woody capsules, and are released following desiccation of the capsule (e.g. see Woodall, 1982; Pettit & Froend, 2001). The mechanism by which capsules open and release seed is not fully understood, but in Eucalyptus spp., a fellow genus in Myrtaceae, it has been postulated that desiccation changes the shape of the ovary wall, forcing the loculi to widen and the capsule valves to open and release seed (Cremer, 1965).
As desiccation of seed capsules may also be required for seed release in M. ericifolia and hydrological factors are likely to be important in controlling the magnitude of seed rain from related species, we examined whether hydrological regimes, salinity, air temperature and atmospheric moisture content influenced the seed rain from aerial seed banks in M. ericifolia. We hypothesised that a pulse of seed rain would be triggered by increased plant water stress as water levels fell and sediments dried out, as water-column salinities increased, and as atmospheric temperature increased and moisture contents decreased. To test these hypotheses, we measured seasonal patterns in seed rain in populations of M. ericifolia that experienced different water regimes in Dowd Morass, a brackish-water wetland in south-eastern Australia.
Materials and methods
Study site
The study was undertaken at Dowd Morass, a large (1,500 ha) brackish-water wetland on the south-western shore of Lake Wellington in Victoria, south-eastern Australia (Fig. 1). Dowd Morass is one of a number of Ramsar-listed wetlands in the larger Gippsland Lakes Ramsar complex. Characteristics of Dowd Morass are described in Raulings et al. (2007) and a general description of M. ericifolia wetlands is provided by Bird (1962) for other wetlands of the Gippsland Lakes. Based on Australian Bureau of Meteorology records since 1971, the area around Dowd Morass experiences maximum/minimum mean daily air temperatures of 25.3/13.1°C and 13.9/3°C in February (summer) and July (winter), respectively. Average monthly rainfall ranges between 40 and 60 mm, with lowest falls in February and highest falls in November.

Study site and locations of seed traps in three areas of Dowd Morass, south-eastern Australia. The internal levees that subdivide the wetland are shown with the light grey lines
The currently accepted paradigm is that, prior to European settlement and consequent changes to the hydrology of the Gippsland Lakes and its rivers, Dowd Morass would have dried periodically, through evaporation and drainage, to expose the sediments approximately every 5 years (Raulings et al., 2007). Since 1975, most of the wetland has been artificially and permanently flooded in order to reduce saline intrusions from Lake Wellington and to provide for waterfowl hunting. Water depths in the permanently flooded areas of the wetland typically fluctuate from ~0.2 m over summer to 0.4–0.8 m over winter. It is believed that this chronic, near-permanent inundation has had adverse impacts on the swamp scrub plant communities of Dowd Morass. In contrast to the central parts of Dowd Morass, inundation of land around the perimeter of the wetland rarely lasts more than a few months (usually July–September) each year. As described later, three areas of Dowd Morass were selected for study to reflect these different hydrological regimes.
Melaleuca ericifolia is the dominant plant species in Dowd Morass, and it forms extensive Swamp Scrub communities over much of the wetland. Phragmites australis (Cav.) Trin. ex Steud., Typha spp. and Vallisneria americana Michx. are present in the understorey. Two growth forms of M. ericifolia are present in Dowd Morass, depending on hydrological regime. In the first growth form, closely spaced upright trees, which form a closed woodland up to 12 m tall, dominate the intermittently flooded sites along the wetland perimeter. The second growth form occurs in the permanently flooded sites, where M. ericifolia occurs as an open woodland up to 4 m tall, with individual trees frequently growing on elevated hummocks composed of woody roots, sediment and organic matter up to 1 m higher than the surrounding sediment and, depending on water regime, extending partially out of the water. Given the usual range of water depths, these hummocks raise the root mass of M. ericifolia towards or above the water level.
Sampling design
Changes in seed rain (measured as seedling emergents: see seedling emergence subsection) in response to hydrologic regime were tested during an experimental draw-down of Dowd Morass carried out as part of a larger study on wetland rehabilitation over the period 2003–2005. Seed rain was measured using traps placed in three areas of Dowd Morass selected on the basis of pre-existing discrete zones within the wetland. These areas were delimited in large part by a series of internal levees built in the 1970s and showed contrasting hydrological regimes (Fig. 1). Both the Flooded west (Flooded-w) and the Flooded east (Flooded-e) sites are more or less permanently inundated, and dominated by the open woodland growth form of M. ericifolia. It was intended that the Flooded-w site would be subjected to a drying event as part of the larger experimental draw-down of water levels in Dowd Morass, where water was intended to be drained from the Flooded-w site using a series of gated drains. To serve as a control area, it was intended that the Flooded-e site would remain flooded whilst the western portions of the wetland were dried out over the summers of 2003–2004 and 2004–2005. The third area used in the study was located at the perimeter of the wetland and was intended to act as a reference area for the hydrological manipulations, since it naturally experienced intermittent flooding rather then the near-permanent inundation that characterised the other two sites. This third area is termed the Intermittent site, and is dominated by the closed woodland growth form. It was selected to assess patterns and triggers in seed rain in M. ericifolia that grew under drier conditions.
Unfortunately, we were not as successful in manipulating water levels as we had anticipated when planning the experimental draw-down for three reasons. Firstly, there was a greater connection between the Flooded-e site and Lake Wellington than previously realised; thus water flowed from the lake into the eastern parts of the wetland. Secondly and most importantly, an act of vandalism perpetrated on the water-control structures allowed water from the adjacent La Trobe River to enter the Flooded-w site while it was in the process of being drawn down; Thirdly, the influx of river water was followed by a high rainfall event (109 mm) in April 2004. Even so, the contrasting environmental conditions in different parts of Dowd Morass still permitted us to examine the effects of key factors on seed release, as well as to quantify overall patterns of seed rain for the wetland.
Seed traps
We placed 20 seed traps below the canopy of individual trees in each of the Flooded-w, Flooded-e and Intermittent sites. Seed traps were based on the funnel trap design of Kollmann & Goetze (1998). Although this trap design may be less effective than some others in capturing wind-blown seed because it is not positioned at the soil surface (e.g. Page et al., 2002), the design allowed us to deploy seed traps in flooded locations with fluctuating water levels. The trap comprised a plastic funnel (220 mm diameter, surface area of the outer perimeter of the funnel 0.038 m²) placed into a PVC pipe rammed into the soil such that the funnels were ~1 m above the soil surface. Trap contents were collected in a muslin bag (0.1 mm mesh size) attached to the base of the funnel. Two holes (5 mm diameter) were drilled into the PVC pipe below the muslin bag to prevent it from filling with rainwater. Bird mesh (20 mm mesh size) was placed across the top of each funnel trap to allow seed to pass into the trap but prevent birds from resting on or entering into the trap. Traps were emptied on 12 occasions between January 2004 and April 2005, but due to loss or damage to traps sufficient data was only available to analyse six of the sampling periods. The contents of each seed trap were stored in paper bags, air-dried, and stored at 20°C.
Seedling emergence
Although the most desirable measure of seed rain would be to count the number of individual seeds from each trap collection, it was not possible to adopt this approach because of the extremely small size of the seeds (0.5–0.8 mm long, 0.2–0.3 mm wide) and the entanglement of seeds with detritus in the diverse materials retained in the seed traps (e.g. spider webs, partly degraded leaf material). Instead, the seedling emergence technique of Roberts (1981) and Brown (1992) was used to estimate the number of viable seeds of M. ericifolia and other species that were caught in the traps.
Perforated plastic trays (160 mm long × 115 mm wide × 40 mm deep) were filled with sifted (10 mm mesh) soil mix containing composted pine bark and coir pith (Mulch ‘N’ Feed, Debco Pty Ltd, Victoria). Material from each trap was spread on top of the soil mix and trays were placed into larger plastic trays (225 mm long × 150 mm wide × 65 mm deep) and watered using the ‘bog method’, the most effective method for germinating Melaleuca seed (Doran & Gunn, 1994). Tap water was poured into the larger tray, which allowed moisture to reach the seeds by capillary action and ensured that the soil mix did not dry out but the water level did not inundate the germination substrata. Water was added weekly or otherwise, as required. Trays were sprayed lightly with an anti-fungus agent (Garden King Fongarid Systemic Fungicide, Envirogreen Pty Ltd, NSW) when necessary.
Seeds were germinated in a constant temperature room at 20°C with a 12-h light/dark cycle. Due to time and space constraints, we only germinated a randomly selected sub-sample of 12 of the 20 trap samples per site at each of the six sampling times; the same 12 traps per site were used throughout the germination trials. A pilot trial established that up to 95% of M. ericifolia seeds germinated within 14 days of sowing and the remainder germinated within 28 days; the germination trial therefore lasted 31 days. To confirm that the soil mix was not contaminated with other seeds, four soil-only trays were included to check for contamination. Emergent M. ericifolia seedlings were counted and removed twice weekly. At the end of the trial, trays containing seedlings which were not M. ericifolia were moved to a glasshouse and left to grow until they were mature enough to be identified.
Hydrological and environmental data
To assess whether differences in seed rain among sites or over time were related to key environmental factors, we quantified water depth, salinity, air temperature and atmospheric moisture content over the study period. Dataloggers that recorded water depth and electrical conductivity (EC) were located in the Flooded-e and Flooded-w sites within 500 m of the seed traps. As the Intermittent site is usually dry, dataloggers were not installed and flooding events were assessed during our regular site visits. Dataloggers installed in the two flooded sites were downloaded by contractors from Theiss Services Pty Ltd. We used these water depth data to infer temporal changes in water depths adjacent to the trees where the seed traps were located. Water depths were measured approximately monthly around the perimeter of each of the 20 trees in the Flooded-e and Flooded-w sites from March 2004 to March 2005; values for each site were averaged. To assess daily changes in water depth within each of the flooded sites, we calculated the difference between the average water depth at trees in each site that we had recorded monthly and the corresponding datalogger depths for those given days. Over the study period, we calculated that the datalogger sites were an average of 29.2 cm (SE = ±0.81) and 24.0 cm (SE = ±0.2) deeper than areas where the traps were installed in the Flooded-w and Flooded-e sites, respectively. Over the study period, daily values of water depth at each site were derived by subtracting this amount from the water-depth data collected by the dataloggers. However, as M. ericifolia occurs on hummocks in both the Flooded-w and Flooded-e sites, water depth per se may not be a true indication of the conditions experienced by the trees. We therefore calculated the percentage of hummock exposed as the difference between the water depth and the average hummock height (Flooded-w site: 42 ± 5 cm; Flooded-e site: 35 ± 5 cm) in each site.
Mean daily air temperature and vapour pressure deficit (VPD; an indicator of atmospheric moisture content) data were provided by the Australian Bureau of Meteorology using data collected at the nearby East Sale Airport meteorology station.
Data analysis
The mean number of M. ericifolia seedlings that emerged during incubation of seed-trap material was standardised to yield the mean number of seedling emergents per metre square of seed trap area per day. This value was calculated using the known surface area of the funnel (0.038 m²) and the number of days between seed trap collections. Data were then transformed (double square root) and analysed using a repeated measures ANOVA with Systat® version 11 (Systat Software Inc, 2004). Box plots and plots of the residual against model estimates were used to confirm normality and homogeneity of variance. Sphericity was tested using the Greenhouse–Geisser (G–G) and Huynh–Feldt (H–F) estimates of epsilon; values less than 1 indicate non-sphericity and require adjustment of the P value (Quinn & Keogh, 2002). We adjusted P values using the G–G adjustment when G–G epsilon values were <0.75, and with the H–F adjustment, which is less conservative, when they were >0.75 (Quinn & Keogh, 2002). Where there was a significant interaction term, we tested for differences between sites at each sampling time using a single factor ANOVA; where significant, a Tukey’s multiple comparison test (α = 0.05) identified sites that differed significantly.
The strength of the linear relationships between water depth and EC in the two flooded sites and between temperature and VPD were analysed by the Pearson correlation coefficient using the Systat® version 11 package. Regression analyses between the mean number of seedling emergents (m−2 day−1) with each of the four environmental variables (water depth, EC, temperature and VPD) were determined with JMP® (Version 3.2.2).
Results
Environmental conditions
Despite their different connections with Lake Wellington and the La Trobe River (Fig. 1), the Flooded-w and Flooded-e sites showed broadly similar temporal patterns in water levels (Fig. 2a). Both sites experienced their lowest water levels between February and April 2004, and water depths increased quickly in April 2004 after the vandalism event and subsequent heavy rain (Fig. 2a). With the exception of the February to May 2004 period, where Flooded-e was 0.15–0.35 m shallower than Flooded-w, water levels in the two parts of the wetland did not differ by more than 0.1 m during the study. Approximately 40–67% of the average hummock in the Flooded-w site and 69–100% of the average hummock in the Flooded-e site became exposed during summer. In contrast to these two more or less permanently inundated sites, only occasional pools of water (not more than 5 cm deep) were present in the Intermittent site from May to November.

a Mean daily water depth at hummocks in the two flooded sites (Flooded-e and Flooded-w) of Dowd Morass during the study period. b Mean daily electrical conductivity (dS m−1) of the two flooded sites during the study period. c Mean daily air temperature (ºC) and vapour pressure deficit (hPa) for the region (East Sale meteorological station) over the study period. The absence of data in some parts of (a) and (b) corresponds to a period when the datalogger in the Flooded-w was out of the water
Consistent with the two flooded sites having similar hydrological regimes, the Flooded-w and Flooded-e sites also experienced broadly similar salinity regimes (Fig. 2b). Water-column salinity rose in February 2004 and reached its highest value in the Flooded-e site in February-March 2004. When there was little water in the Flooded-e site, salinities could be as high as 27 dS m−1 (~½ sea water). Salinity fell in the Flooded-e site in April 2004, probably as a consequence of dilution following the heavy rainfall that caused the increase in water levels. Salinities in both sites rose gradually and did not fall again throughout the rest of the study period. A strong negative linear correlation (r 2 = 0.819) was evident between water depth and salinity for these sites. As VPD is a function of air temperature and atmospheric moisture content, the two factors are correlated (r 2 = 0.661) and showed similar patterns throughout the study period (Fig. 2c); both variables were highest between January and April and lowest in August 2004.
Seedling emergence
Although nine plant species (M. ericifolia, Atriplex prostrata Boucher ex DC., Senecio glomeratus Desf. ex Poir., Chenopodium glaucum L., two unidentified monocots and three unidentified dicots) germinated from the material collected in the seed traps, the emergents were markedly dominated by M. ericifolia (80% of all the emergents in the Intermittent site, 83% of emergents in the Flooded-w site, and 97% of emergents in the Flooded-e site). The mean number of M. ericifolia emergents was higher in the Flooded-e (135.7 ± 4.9 emergents m−2 day−1) and Intermittent (134.5 ± 26.2 emergents m−2 day−1) sites than in the Flooded-w (41.7 ± 4.9 emergents m−2 day−1) site. There were no emergents in any of the control trays that contained soil only, indicating there was no prior contamination of the soil mix.
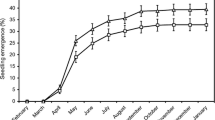
Although we anticipated differences in seed rain between the two flooded sites and the intermittently flooded site, the general pattern of seed release from M. ericifolia was in fact similar between the Intermittent site and the Flooded-e site. Both sites produced similar numbers of emergents (up to ~100 emergents m−2 day−1) at comparable times. As shown in Fig. 3, a pulse of seed release in the Flooded-e and Intermittent sites occurred in February (33 ± 11 emergents m−2 day−1 in the Flooded-e; 45 ± 5 in the Intermittent site) and April 2004 (68 ± 30 in the Flooded-e site; 55 ± 27 in the Intermittent site). During the same period, however, the Flooded-w site produced far fewer emergents (9 ± 2 and 6 ± 2 emergents m−2 day−1 in February and April, respectively).

Seed-trap emergence data for Dowd Morass, showing mean number of M. ericifolia emergents (±SE, n = 12) counted during incubation of seed-trap material from the three areas over the study period
A repeated-measures ANOVA showed a significant difference among the three sites in the number of M. ericifolia emergents (F 2,33 = 10.236, P < 0.001), as well as significant differences with time (F 5,165 = 25.042, P < 0.001, H–F adjustment) and a significant interaction term (F 10,165 = 3.445, P = 0.001, H–F adjustment). Due to the significant interaction term, a subsequent single-factor ANOVA was conducted to test for site effects at each sampling time. This analysis indicated site effects at only two of the six sampling times: February 2004 (F 2,33 = 12.273, P < 0.001) and April 2004 (F 2,33 = 9.138, P < 0.001). Multiple comparisons using Tukey’s tests (P < 0.05) demonstrated that the Flooded-w site produced significantly less emergents than the other two sites in both the February 2004 and April 2004 sampling periods. Indeed, the Flooded-w site contributed only 13% of all M. ericifolia emergents over the entire period of observation.
Relationships between seedling emergence and environmental conditions
For the Flooded-e site, regression analysis showed that there was a significant negative correlation between water depth and the number of emergents per unit of wetland area: log (emergents) = 3.914 − 0.042 (water depth), r 2 = 0.71, F 1,4 = 9.56, P = 0.03 (Fig. 4a). The relationship, however, was strongly influenced by two data points which had a high number of emergents at water depths less than 10 cm. The large standard error associated with these data points suggests the response to low water levels was variable among different trees. In water deeper than about 16 cm, relatively fewer seeds were released and the response was similar across trees and sites, as indicated by the small standard error bars of these points. In the Flooded-w site, water levels did not fall to the same extent as observed in the Flooded-e site, and seed rain was not as strongly influenced by water level: log (emergents) = 2.551 − 0.0240 (water depth), r 2 = 0.281, F 1,4 = 1.56, P = 0.28 (Fig. 4a).

Relationships between mean number of M. ericifolia emergents with: a mean water depth, b mean daily electrical conductivity of the water column and, c mean daily air temperature. Regressions and full statistical results are provided in the text. Hatched circles, filled circles and open squares represent the Flooded-e, Flooded-w and Intermittent sites, respectively. The Intermittent site is not included in (b) as it was dry and hence there were no salinity data available for this site
As water depth and salinity were themselves correlated as described earlier, it is not surprising that we found a similar pattern of seed release in response to salinity as observed for water depth. As noted earlier for the effect of water depth, however, the relationship for water-column salinity was largely driven by two extreme data points from the Flooded-e site that were exposed to salinities greater than ~20 dS m−1. The regression analysis for the relationship between salinity and the number of emergents per unit of wetland area was not significant in either the Flooded-e (log (emergents) = 0.657 + 0.133 (salinity), r 2 = 0.507, F 1,4 = 4.11, P = 0.113) or the Flooded-w (log (emergents) = 1.013 + 0.09 (salinity), r 2 = 0.188, F 1,4 = 0.926, P = 0.39) sites. Lack of statistical significance in response to salinity is likely to have resulted from high variability among trees at each site requiring greater replication. The weaker response of seed rain to salinity in the Flooded-w site was most likely because the site was not exposed to salinities above 12 dS m−1.
Although water depth and salinity strongly influenced seed rain in M. ericifolia at the flooded sites, large variation in seed rain was also observed at the Intermittent site, where trees were not subjected to near-permanent flooding with water of variable salinity. Responses observed at the Intermittent site suggest that other factors may also influence seed rain, in particular mean daily air temperature (Fig. 4c). However, the regression analysis was not significant in the Intermittent site: log (emergents) = 0.189 + 0.181 (air temperature), r 2 = 0.46, F 1,4 = 3.41, P = 0.238. Neither were the relationships significant for either the Flooded-e site (log (emergents) = 0.355 + 0.158 (air temperature), r 2 = 0.37, F 1,4 = 2.323, P = 0.202) or the Flooded-w site (log (emergents) = −0.2751 + 0.13545 (air temperature), r 2 = 0.47, F 1,4 = 3.62, P = 0.129). As the response to air temperature was similar for the Intermittent and Flooded-e sites, both of which experienced drying, the data from the two sites were pooled for this analysis. Variations in mean air temperature explained ~41% of variation in seed rain: log (emergents) = 0.18681 + 0.16904 (air temperature), r 2 = 0.41, F 1,10 = 7.03, P = 0.024 (Fig. 4c). Relationships between the number of emergents and VPD were weak and, in general, similar to those reported for mean air temperature.
Discussion
Aerial seed banks comprise a source of propagules for many taxa of woody wetland plants. However, despite the potential importance of this source of propagules for the regeneration of these species, the size of the contribution, the patterns of seed fall and triggers for seed release have been subject to surprisingly little study in the past. In our study, we quantified the seed rain from the aerial seed bank (as emergent seedlings of M. ericifolia) and investigated the possible role of a number of environmental variables in this release of canopy-stored seed in a type of brackish-water wetland that is common across south-eastern Australia. Under flooded conditions, we observed a low but consistent background seed rain from the canopy of M. ericifolia and a pulse of seed release during the draw-down of water levels in April 2004. The lowered water levels of March 2005 did not however achieve a similar pulse of seed release. We believe that these observations have important implications for understanding the mechanism and frequency with which woody plants recruit in wetlands, as well as for strategies to rehabilitate wetlands that are vegetated with emergent woody taxa and have suffered prior degradation.
In some aquatic systems, the soil seed bank represents a major mechanism for plant regeneration, but this situation seems not to apply to the brackish-water wetlands of the Gippsland Lakes, where woody vegetation is the dominant plant form. For example, sediment cores from Hacks Lagoon in South Australia and Cooper Creek (Lake Eyre Basin) in Central Australia produced 78,370 seedlings (Nicol et al., 2003) and 10,514 seedlings (Capon & Brock, 2006) per m2 of soil, respectively. Thus, wetland soils in these two sites represent a crucial and substantial reservoir of seeds which allow plant communities to recover after natural or anthropogenic disturbance. The soil seed bank at Dowd Morass is, by comparison, extremely depleted; preliminary studies indicate a soil seed bank of only 129 seedlings per m2 across all plant taxa in this wetland, and we have never observed M. ericifolia seedlings to emerge from soil cores collected at Dowd Morass (unpublished data). Similar results for another M. ericifolia wetland have been reported earlier by Ladiges et al. (1981), who found that there was no soil-based reservoir of M. ericifolia seed at Koonwarra, a wetland site also located in the Gippsland region of south-eastern Victoria. In the absence of a significant soil seed bank, sexual regeneration in M. ericifolia must be reliant on the release of seed from aerial seed banks when environmental conditions are suitable for the seeds to germinate and establish in the population.
Earlier studies of aerial seed banks in terrestrial environments have shown that the conditions required for seed release vary among vegetation types. For monsoon rainforest trees, peak seed release occurs during the wet season in perennially wet sites, and in sites that are seasonally dry, peak seed release occurs in or shifts towards the dry season (Russell-Smith & Setterfield, 2006). Even during the period of peak seed release, however, less than 15 seedlings m−2 day−1 were produced from these terrestrial monsoonal areas. In our study, M. ericifolia had a peak of 100 m−2 day−1 in April 2004 and the release was strongly associated with lowering of water levels. Studies of seed held in the canopy of terrestrial plants have shown that seed release occurs largely in response to fire for a wide range of genera (e.g. Pinus: Schwilk & Ackerly, 2001; Goubitz et al., 2003; Tapias et al., 2004; Leptospermum: Harris, 2002; Hakea and Eucalyptus: Enright & Goldblum, 1999). Although fire could similarly play a role in seed release from wetlands dominated by woody taxa, and Swamp Scrub wetland communities in south-eastern Australia do occasionally catch fire, our study demonstrated that water level, salinity, air temperature and VPD influence seed rain from M. ericifolia in Dowd Morass.
The mechanism by which changing water levels drive the release of seed from the canopy of M. ericifolia is probably related to water availability to adult plants. We posit that draw-down of water level deprives the roots of M. ericifolia plants growing on hummocks access to water, which in turn promotes the desiccation of plant organs, including the seed capsules, and ultimately facilitates seed release. Certainly trees in the Flooded-e site released a pulse of seed when the water fell to less than about ~10 cm in April 2004, a level that exposed at least 69% of each hummock and, presumably, most if not all of the M. ericifolia roots. The hummocks in the Flooded-w sites were never completely exposed in this study, and trees are likely to have utilised the surface water that surrounded the base of the hummocks. It is important to note that M. ericifolia is a shallow-rooted plant, even though it can grow to over 12 m in height (e.g. see Bird, 1962), and presumably can utilise shallow surface waters. However, the draw-down of March 2005 did not trigger a similar pulse of seed release like we had observed during the draw-down of April 2004. The extent of draw-down in the Flooded-e site in March 2005 was not as great as the previous year, and exposed up to 57% of each hummock. In addition, we postulate that after ~30 years of flooding, the 2004 draw-down resulted in an almost total loss of mature seed from the canopy of M. ericifolia, and that there were few mature seeds present on the canopy during the draw-down event of the following year. This has important implications for the frequency with which draw-down events occur. Melaleuca ericifolia produces seed annually which take 2–3 years to mature and can be held on the canopy for up to 5 years. If draw-downs occur close together in time (less than 2 years apart) they are likely to be unsuccessful in facilitating the sexual recruitment of M. ericifolia; conversely, if flooded conditions persist longer than 5 years, seed of M. ericifolia will be lost from the canopy and are likely to be wasted in the flooded conditions. We propose that sexual recruitment of M. ericifolia will be maximised if draw-down events occur 2–5 years apart.
Our data indicate that high water-column salinity also contributed positively to seed release in M. ericifolia. This effect is understandable since salinity influences water availability via its effect on osmotic potential of the soil solution (Kozlowski, 1997). It is not possible, however, to fully unravel the individual roles of water level draw-down and salinity on seed release with our data because the strong inverse relationship between salinity and water depth complicate the exact influence each has on water availability and plant desiccation.
Seed release from the Intermittent site was highly variable and changes in soil moisture or soil salinity, which were not monitored at this site, may have contributed to the variation in seed rain observed at these peripheral areas of the wetland that experience only intermittent inundation. Changes in seed rain were explained in part by changes in air temperatures at both the Intermittent site and the Flooded-e site. In contrast, at the Flooded-w site where hummocks were not fully exposed, air temperatures exerted only a small effect on seed rain. Changes in VPD exerted similar effects on seed release to that observed for air temperature, and as these factors were closely correlated it is unclear which is the primary driver, although relationships with air temperature were slightly stronger than for VPD. We conclude that high air temperatures and/or high VPD may exert a stronger influence on seed release when plants experienced a water deficit associated with soil drying or increasing salinity. This hypothesis needs to be tested through more detailed studies of the relationships between soil conditions, plant water stress and seed rain.
In a wetland system where the regeneration of woody plants is reliant on aerial seed banks, sexual recruitment will be dependent on regular seed rain from canopy-held seed and the successful germination and establishment of the resultant seedlings. Accordingly, seeds need safe sites (Fowler, 1988) in order to contribute to future recruitment success. The environmental requirements for seed germination and seedling establishment of M. ericifolia are quite well known: seeds germinate on bare sediment that is fresh or slightly saline and cannot establish well under inundated conditions (Ladiges et al., 1981). In the permanently flooded areas of Dowd Morass, M. ericifolia seeds would need to float away from flooded adult trees to more suitable areas elsewhere in the wetland (e.g. intermittently flooded areas around the perimeter of the wetland) for sexual recruitment to be successful. During draw-down, dispersal would presumably be confined to restricted areas around the parent plant unless wind dispersed seeds further afield. Either way, it is the duration of the draw-down period that would be most important to the sexual recruitment of M. ericifolia since it establishes the conditions that not only allow for seed release but also those conditions that are best for germination and seedling establishment. Although M. ericifolia seedlings do not appear to tolerate long periods of complete submergence in summer, particularly if conditions are saline (Raulings et al., 2007; Salter et al., 2007), Salter et al. (2008) have found that if M. ericifolia seedlings are flooded with fresh water in winter when dormant, and re-exposed in spring during their growth period, then the young seedlings can rapidly recover from these seemingly adverse environmental conditions. The findings of Salter et al. (2008) suggest that the impact of flooding on sexual recruitment in M. ericifolia would be strongly influenced by how hydrological events link to the dormant and active phases of a plant’s growth cycle.
The results reported in this paper have important implications for the management and rehabilitation of wetlands dominated by woody taxa with aerial seed banks. Rehabilitation of Melaleuca-dominated wetland ecosystems is a conservation priority in south-eastern Australia because of the high conservation value of these habitats and their disproportionate loss and degradation arising from nearly two centuries of human interference (de Jong, 1997). Rehabilitating wetland habitats is a complex process, as facilitating recruitment and rehabilitating vascular plant communities is likely to be affected by a wide range of synergistically interacting environmental factors, such as water level, water-column salinity and soil salinity. Middleton (2000) reported a similar complexity for Taxodium-dominated wetlands in Illinois (USA), where a specific set of environmental conditions (flood pulse of high water in winter to maximise seed dispersal, followed by low water levels in summer to facilitate seed germination and seedling recruitment) were needed to rehabilitate degraded wetland sites. In Dowd Morass, however, it is apparent that simply restoring the hydrological regime to one that reflects more accurately pre-1970s conditions is not straightforward nor can it be assumed, by itself, to result in improvements in vegetation condition or successful recruitment.
This study is one of the first to demonstrate the critical importance of aerial seed banks to sexual recruitment in woody wetland plants and indicates that canopy-held seeds should be considered along with soil seed banks in the establishment and long-term survival of plants in wetlands, as well as in future strategies designed to rehabilitate such high-value wetlands.
References
Abernethy, V. J. & N. J. Willby, 1999. Changes along a disturbance gradient in the density and composition of propagule banks in floodplain aquatic habitats. Plant Ecology 140: 177–190.
Baldwin, A. H., K. L. McKee & I. A. Mendelssohn, 1996. The influence of vegetation, salinity and inundation on seed banks of oligohaline coastal marshes. American Journal of Botany 83: 470–479.
Barrett, L. G., T. H. He, B. B. Lamont & S. L. Krauss, 2005. Temporal patterns of genetic variation across a 9-year-old aerial seed bank of the shrub Banksia hookeriana (Proteaceae). Molecular Ecology 14: 4169–4179.
Bird, E. C. F., 1962. The Swamp Paper-bark. The Victorian Naturalist 79: 72–81.
Boedeltje, G., J. P. Bakker & G. N. J. Ter Heerdt, 2003. Potential role of seed banks in the development of aquatic vegetation in backwaters along navigation canals. Aquatic Botany 77: 53–69.
Britton, D. L. & M. A. Brock, 1994. Seasonal germination from wetland seed banks. Australian Journal of Marine and Freshwater Research 45: 1445–1457.
Brock, M. A. & D. L. Britton, 1995. The role of seed banks in the revegetation of Australian temporary wetlands. In Wheeler, B., S. Shaw, W. Fojt & A. Robertson (eds), The Restoration of Temperate Wetlands. Wiley, Chichester: 183–188.
Brown, D., 1992. Estimating the composition of a forest seed bank: a comparison of the seed extraction and seedling emergence methods. Canadian Journal of Botany 70: 1603–1612.
Capon, S. J. & M. A. Brock, 2006. Flooding, soil seed bank dynamics and vegetation resilience of a hydrologically variable desert floodplain. Freshwater Biology 51: 206–223.
Cremer, K. W., 1965. How eucalypt fruits release their seed. Australian Journal of Botany 13: 11–16.
de Jong, M., 1997. Register of Wetland Restoration Projects in Australia and New Zealand. Wetlands Management Program, Department of Environment and Natural Resources, Adelaide.
Denton, M. & G. G. Ganf, 1994. Response of juvenile Melaleuca halmaturorum to flooding: management implications for a seasonal wetland, Bool Lagoon, South Australia. Australian Journal of Marine and Freshwater Research 45: 1395–1408.
Doran, J. C. & B. V. Gunn, 1994. Exploring the genetic resources of tropical melaleucas. Forest Genetic Resources 22: 12–24.
Enright, N. J. & D. Goldblum, 1999. Demography of a non-sprouting & resprouting Hakea species (Proteaceae) in fire-prone Eucalyptus woodlands of southeastern Australia in relation to stand age, drought and disease. Plant Ecology 144: 71–82.
Fowler, N. L., 1988. What is a safe site?: Neighbor, litter, germination date, and patch effects. Ecology 69: 947–961.
Goubitz, S., M. J. A. Werger & G. Ne’eman, 2003. Germination response to fire-related factors of seeds from non-serotinous and serotinous cones. Plant Ecology 169: 195–204.
Harris, W., 2002. Variation of inherent seed capsule splitting in populations of Leptospermum scoparium (Myrtaceae) in New Zealand. New Zealand Journal of Botany 40: 405–417.
Kollmann, J. & D. Goetze, 1998. Notes on seed traps in terrestrial plant communities. Flora 193: 31–40.
Kozlowski, T. T., 1997. Responses of woody plants to flooding and salinity. Tree Physiology Monograph 1: 1–29.
Ladiges, P. Y., P. C. Foord & R. J. Willis, 1981. Salinity and waterlogging tolerance of some populations of Melaleuca ericifolia Smith. Australian Journal of Ecology 6: 203–215.
Liu, G. H., W. Li, E. H. Li, L. Y. Yuan & A. J. Davy, 2006. Landscape-scale variation in the seed banks of floodplain wetlands with contrasting hydrology in China. Freshwater Biology 51: 1862–1878.
Middleton, B. A., 2000. Hydrochory, seed banks, and regeneration dynamics along the landscape boundaries of a forested wetland. Plant Ecology 146: 169–184.
Middleton, B. A., 2003. Soil seed banks and the potential restoration of forested wetlands after farming. Journal of Applied Ecology 40: 1025–1034.
Nicol, J. M., G. G. Ganf & G. A. Pelton, 2003. Seed banks of a southern Australian wetland: the influence of water regime on the final floristic composition. Plant Ecology 168: 191–205.
Page, M. J., L. Newlands & J. Eales, 2002. Effectiveness of three seed-trap designs. Australian Journal of Botany 50: 587–594.
Parker, V. T. & M. A. Leck, 1985. Relationships of seed banks to plant distribution patterns in a freshwater tidal wetland. American Journal of Botany 72: 161–174.
Pettit, N. E. & R. H. Froend, 2001. Variability in flood disturbance and the impact on riparian tree recruitment in two contrasting river systems. Wetlands Ecology and Management 9: 13–25.
Quinn, G. P. & M. J. Keogh, 2002. Experimental Design and Data Analysis for Biologists. Cambridge University Press, Cambridge.
Raulings, E. J., P. I. Boon, P. C. Bailey, M. C. Roache, K. Morris & R. Robinson, 2007. Rehabilitation of Swamp Paperbark (Melaleuca ericifolia) wetlands in south-eastern Australia: effects of hydrology, microtopography, plant age and planting technique on the success of community-based revegetation trials. Wetlands Ecology and Management 15: 175–188.
Rayamajhi, M. B., T. K. Van, T. D. Center, J. A. Goolsby, P. D. Pratt & A. Racelis, 2002. Biological attributes of the canopy-held melaleuca seeds in Australia and Florida. U.S. Journal of Aquatic Plant Management 40: 87–91.
Roberts, H. A., 1981. Seed banks in soils. In Cóaker, T. H. (ed.), Advances in Applied Biology. Academic Press, London: 1–55.
Russell-Smith, J. & S. A. Setterfield, 2006. Monsoon rain forest seedling dynamics, northern Australia: contrasts with regeneration in eucalypt-dominated savannas. Journal of Biogeography 33: 1597–1614.
Salter, J., K. Morris, P. C. E. Bailey & P. I. Boon, 2007. Interactive effects of salinity and water depth on the growth of Melaleuca ericifolia Sm. (Swamp paperbark) seedlings. Aquatic Botany 86: 213–222.
Salter, J., K. Morris & P. I. Boon, 2008. Does salinity reduce the tolerance of two contrasting wetland plants, the submerged monocot Vallisneria australis and the woody shrub Melaleuca ericifolia, to wetting and drying? Marine and Freshwater Research 59: 291–303.
Schneider, R. L. & R. R. Sharitz, 1988. Hydrochory and regeneration in a bald cypress-water tupelo swamp forest. Ecology 69: 1055–1063.
Schwilk, D. W. & D. D. Ackerly, 2001. Flammability and serotiny as strategies: correlated evolution in pines. Oikos 94: 326–336.
Tapias, R., J. Climent, J. A. Pardos & L. Gil, 2004. Life histories of Mediterranean pines. Plant Ecology 171: 53–68.
van der Valk, A. G. & C. B. Davis, 1978. The role of seed banks in the vegetation dynamics of prairie glacial marshes. Ecology 59: 322–335.
Woodall, S. L., 1982. Seed dispersal in Melaleuca quinquenervia. Florida Scientist 45: 81–93.
Acknowledgements
We are grateful to John Topp from Gippsland Indigenous Plants for his assistance in collecting the seed trap material. We thank Jacqueline Salter, Elizabeth Wallis, Shauna Tupper and Dr Ni Watiniasih for access to their data. We also thank Dr Ross Thompson and the two anonymous reviewers for their invaluable comments and suggestions on earlier drafts of this paper.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Luis Mauricio Bini
Rights and permissions
About this article
Cite this article
Hamilton-Brown, S., Boon, P.I., Raulings, E. et al. Aerial seed storage in Melaleuca ericifolia Sm. (Swamp Paperbark): environmental triggers for seed release. Hydrobiologia 620, 121–133 (2009). https://doi.org/10.1007/s10750-008-9623-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-008-9623-6