
Abstract
This study analyzes the interaction of fish assemblages with 14 physicochemical and hydrogeomorphological variables at 31 sampling stations along the watershed of the Meia Ponte River, Upper Paraná Basin, Central Brazil, during low and high water seasons in 2001. This river and its tributaries drain both urban and agricultural areas. Fish were caught with sieves along a 100 m stretch demarcated in every sampling site, where environmental variables were also measured. A total of 3508 individuals belonging to 31 species were collected. Fish abundance and environmental data matrices were submitted to a multivariate analysis of co-inertia. Two axes were retained for interpretation (total variance explained = 63.65%) indicating that pH, water temperature, conductivity, chemical dissolved oxygen, and turbidity, all have an influence on fish assemblage structure. The co-structure found (fish assemblages and physicochemical variables) is correlated in both of the axes considered (r = 0.73 and r = 0.68, respectively), and is statistically significant (Monte Carlo test, P < 0.001). This co-structure is regulated by seasonality, but is influenced by fish habitat preferences, spawning and available food, the extent and effects of anthropogenic activities (domestic sewage, agriculture, ranching, urban areas) and the position of sampling stations along the watershed.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Neotropical aquatic environments have been intensively used by humans in several ways, such as, consumption, irrigation, hydroelectric dams, sewage, and industrial discharge (Pringle et al., 2000). This situation makes the ecological study of any aquatic ecosystem more complex. Therefore, in addition to the biotic and abiotic interactions and the exchanges between aquatic and terrestrial environments (Lowe McConnell, 1999), it is necessary to consider the influence of anthropogenic activities that may modify the conditions of aquatic habitats. Camargo et al. (1996) and Sabater et al. (2000) indicate that land use of a basin modifies physicochemical water characteristics. Leite et al. (2000), Holomuzki & Biggs (2000) and Duarte & Araújo (2001) point out that the amount of solid particles suspended in the water, a condition caused by erosion as a result of anthropogenic activities, modifies turbidity, temperature, hardness, and conductivity.
In turn, alterations of these environmental variables influence fish assemblage structure (Matthews, 1998; Lammert & Allan, 1999; Tejerina-Garro et al., 2005). They may lead to modifications in reproductive patterns, feeding behavior, and growth rates of fish (Böhlke et al., 1978; Gondin-Ferreira, 1993; Araújo, 1996), and predator–prey relationship (Rodríguez & Lewis, 1997; Tejerina-Garro et al., 1998). Fish are considered excellent indicators of the condition or health of ecosystems (Lyons et al., 1995), and are used to evaluate the effects of different types of environmental stressors or pressures on fish assemblages (Karr, 1981), or to evaluate a specific impact using a single species (Schulz & Martins-Junior, 2001).
However, it is necessary to know how the fish-habitat relationship responds to anthropogenic impacts (Barnes, 1998). One approach used to evaluate the effect of environmental modification on fish assemblages (Davis, 1995; Camargo et al., 1996) is to compare the current fish assemblage data with historical data (Paul & Meyer, 2001). But, in many Neotropical rivers, there are no historical fish data available and the fish-habitat relationships are poorly known, although the aquatic environment has already been modified by anthropogenic activities.
The aim of this article is to respond to the following question: which environmental variables among 14 physicochemical and hydrogeomorphological ones structure the fish assemblages in water courses that drain regions under anthropogenic influences in Central Brazil?
Materials and methods
Sampling area
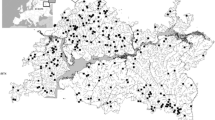
The Meia Ponte River belongs to the upper Paraná River basin. It rises in the Brandões Mountains in the Itauçú Municipality, Goiás State, Central Brazil. The river flows for 480 km, before joining the Paranaíba River, draining an area of 12,350 km2 in 35 municipalities of the State of Goiás (Fig. 1). Some of the rivers in this basin are used for water supply and irrigation (Fialho & Tejerina-Garro, 2004). The area sampled in the Meia Ponte River basin is located between the 16°16’38” and 18°32’53” S parallels, and the 48°46’38” and 49°44’51” W meridians. In this area, 31 stations located in tributaries and one in the main channel of the Meia Ponte River were sampled (Fig. 1).

Sampling sites (black dots) in the Meia Ponte River basin, Central Brazil. Numbers identify sampling sites. Pentagon = cities with 10–40 inhabitants/km2; dotted line = towns with > 100 inhabitants/km2; dashed line represents the border of the basin
Sampling protocols
In each station, 100 m long stretches were randomly selected to collect fish, during high (March 2001) and low (August 2001) water seasons. As the rivers sampled are small (creeks), fish were caught by sieves (3 mm mesh) over a period of 60 min and fixed in 10% formalin. In the laboratory, the fish were identified, measured, and weighed. For each stretch, several environmental variables were measured (channel width, water depth, water velocity—General Oceanics Model 2030 flow meter, pH—pHmeter F-1002, water temperature—Guterm-180 thermometer, conductivity—F-1000 conductivimeter, turbidity—LaMotta 2020 turbidimeter, and dissolved oxygen—F-1001 oxymeter). Water samples were taken to determine the chemical demand of oxygen (CDO—titrametric method with dichromate of potassium in a Tecnal digestor). Hardness was determined using EDTA 0.01 N titrametric, and both phosphate and nitrite were measured using a DREL HACH-2000 spectrophotometer. Sediment residues were determined using a gravimetric method, whereas fecal coliforms were estimated through use of the multiple tubes method. These measurements were all carried out at the Environmental Agency Laboratory of the State of Goiás.
Data analyses
First, we generated a fish abundance data matrix. To eliminate the effects of rare species on the ordination, only the species with more than 10 individuals captured were considered. This matrix was summarized by a factorial analysis of correspondence (FAC) and environmental variables were summarized by a Principal Components Analysis (PCA). The collinearity between environmental variables resulting from the PCA was tested using a Pearson correlation test (Zar, 1998) following the removal of the variable phosphate from the model. Then, a co-inertia analysis was performed using FAC and PCA results to identify a possible co-structure between species abundance and environmental variables (Gauch, 1982). This analysis is powerful even when few samples are available (Dolédec & Chessel, 1994). The co-structure between species abundance and environmental variables resulting from the co-inertia analysis was tested using a Monte Carlo test (1,000 iterations). All analyses were performed using ADE-4 software (Thioulouse et al., 2001).
Several one-way “t” tests were also performed to compare the averages of each variable between the high and low water seasons. As this approach can increase the probability of Type I error, we used the protected ANOVA protocol (Scheiner & Gurevitch, 1993) considering that an ANOVA is a generalization of the “t” test (two group comparison). In this protocol, a MANOVA is first applied (for the factor water season), and only if it was significant, would a one-way “t” test for each variable be conducted separately. Assumptions of “t” tests were examined by the Shapiro–Wilk (normality) and Levene (homogeneity of variances) tests. The parametric tests (MANOVA and “t” tests) were conducted using Statistica 6.0 software.
Results
A total of 3508 fish individuals of 31 different species were collected. A similar number of fish were sampled in both seasons (high water = 1746; low water = 1762). However, the number of individuals of dominant species, such as Poecilia reticulata, Astyanax eigenmanniorum, Bryconamericus stramineus, Astyanax fasciatus, and Hypostomus ancistroides varied greatly between seasons (Table 1) and among sampling stations (e.g., Poecilia reticulata was absent in sampling station 31, but 181 individuals were collected in sampling station 1).
The co-inertia analysis ordinated fish species and sampling stations according to the environmental variables in two axes (they explained 63.65% of the total variance). Correlation between environmental variables and fish abundance in the sampling stations was significant for both axes (r = 0.73 and r = 0.68, respectively; Monte Carlo test P < 0.001; Table 2).
Taking in account the protected ANOVA protocol, the MANOVA was significant for water season (low and high water) (D. F. = 2.62; R of Rhao/F = 108.39; P = 0.000) indicating the appropriateness of applying one-way “t” test” for each variable. The “t” test indicated significant differences for seven variables (Table 3) that met the assumptions of the “t” test (Shapiro–Wilk > 0.05; Levene > 0.05).
On axis 1, the co-structure of fish species and sampling stations was influenced by pH and water temperature, but regulated by seasonality (all the black circles, indicating high water, are located on the left side of the ordination; Fig. 2). In the low water season, the armored catfish Hypostomus sp. B, the characins Astyanax scabripinis, Bryconamericus stramineus, the catfish Phenacorhamdia tenebrosa and Rhamdia quelen, and the guppy Poecilia reticulata are associated with alkaline water (mean pH = 7.68) and low water temperature (mean = 18.40°C). However, in the high water season, the characin Bryconamericus sp. is related to acidic water (average = 6.0) and high water temperature (average = 24.16°C) (Fig. 2). Both, pH and water temperature showed significant differences between low and high water (Table 3).

Discrimination of fish species and sampling sites (A) according to environmental variables (B). Only species and environmental variables with significant contributions are displayed. Black circles = sampling in high water; crosses = sampling in low water. Numbers correspond to sampling sites. Species codes correspond to those mentioned in Table 1. Code of environmental variables corresponds to those presented in Table 3. Small boxes indicate the graphic scale
On axis 2, ordination revealed two groups, one formed by fish assemblages at sampling stations 2–6, 8, 21 and 25 in both seasons. The other group was represented by sampling stations 10–20, 22–24, and 26–31, in both seasons, but more evident during the low water season (Fig. 2). In the first group, characins Astyanax altiparanae, A. eigenmanniorum and Cyphocharax modestus; catfish Rhamdia quelen and Cetopsorhamdia iheringi; armored catfish Hypostomus sp. C and H. ancistroides were associated to high values of conductivity, CDO, and turbidity in both low (average of 121.20 μS/cm; 9.83 mg/l of O2; 13.57 NTU, respectively) and high (average of 101.34 μS/cm; 17.46 mg/l of O2; 72.71 NTU, respectively) water seasons. In the second group, during low water season, the characins Hyphessobrycon sp. was related to low conductivity (average = 42.47 μS/cm), CDO (average = 6.08 mg/l of O2), and turbidity (average = 11.75 NTU).
Discussion
The influence of environmental variables on fish assemblage structure is one approach to understand fish-habitat relationships. Rivers are highly variable environments, and are periodically subjected to unpredictable oscillations of their physical and chemical habitat (e.g., flow, temperature, dissolved oxygen, pH, and conductivity), and these fluctuations have been shown to affect the richness and structure of river fish assemblages (Tejerina-Garro et al., 2005). However, in the Neotropical regions alternating high and low water seasons influence habitat (Poff et al., 1997) and fish assemblages (Mérona & Gascuel, 1993) inclusive of the Meia Ponte River tributaries (Fialho et al., 2007). In this case, seasonality can be seen as a key factor switching on/off other environmental variables and increasing or inhibiting environmental modifications (Agostinho & Zalewski, 1995). This seems to be the case in this study, where seasonality regulates the fish-habitat relationship, especially through pH and water temperature.
Despite seasonality the fish-habitat relationship is also influenced by anthropogenic impacts (e. g., Balon, 1993; Penczak et al., 1994; Moyle, 1995; Camargo et al., 1996; Sabater et al., 2000; Wiens, 2002). Agriculture and urbanization are the human activities that threaten aquatic environments in Europe, North America (Paul & Meyer, 2001), and South America. This is the case of the Meia Ponte River basin, where 32.7% of the population of Goiás State (about 1.6 million inhabitants) is concentrated, and the main economic activities are agriculture and cattle raising (Galinkin, 2003). These anthropogenic activities influence the physical (hydrology, geomorphology, and water temperature), chemical (nutrients, metals, and organic compounds), and biological (fauna and flora) characteristics of water courses (Camargo et al., 1996; Paul & Meyer, 2001; Giller & Malmqvist, 2000).
The pH of running waters is influenced by regional seasonality (Soulsby et al., 2001), for example, during the rainy season low pH water conditions can be predominant because of leaching iron by rain from the watershed surfaces (Giller & Malmqvist, 2000). This situation can be found in Central Brazil soils, where the Meia Ponte River watershed is located, because of the predominance of oxisols (ferralitic soils) (Ratter et al., 1997). This can explain the observed significant differences in pH between high (average = 6.0) and low water seasons (average = 7.68). However, water pH also can be influenced by industrial and domestic sewage (Schulz & Martins-Junior, 2001), and by calcium and magnesium washed from urban areas (Ometo et al., 2000; Araújo & Pfrimer, 2005).
The water pH influences fish in key ways. Acidic pH produces osmoregulatory disturbances in fish resulting in significant ion (Na, Cl, K, and Ca) loss (Shuter et al., 1989). Water pH determines habitat preferences for the reproduction of some species (Dei Tos et al., 2002), and affects fish development and growth (Ferreira et al., 2001). It also controls the changes between the harmless ammonium (NH +4 ) and the toxic undissociated ammonia (NH3) (Lampert & Sommer, 1997).
In the Meia Ponte River, characins (e. g., Bryconamericus stramineus), catfish (e. g., Phenacorhamdia tenebrosa), an armored catfish (Hypostomus sp. B), and an introduced guppy (Poecilia reticulata) were associated with alkaline water. Froese & Pauly (2006) describe similar pH preferences for Bryconamericus stramineus (5.5–7.2) and Poecilia reticulata (7–8). However, the association we found, could also be related to anthropogenic impacts such as domestic sewage pumped into rivers, which increases the abundance of Chironomids (flies) (deBruyn et al., 2003), one of the food items consumed by Bryconamericus stramineus (Casatti & Castro, 1998). On the other hand, the multiple spawning per year displayed by Poecilia reticulata (Froese & Pauly, 2006), associated with some habitat characteristics such as low water volume and velocity could also explain the presence of this species during low water.
Water temperature is another environmental variable that influenced fish assemblages in the Meia Ponte River basin (average high water temperature = 23.92°C; average low water temperature = 18.73°C). Although water temperature changes are associated with seasonality in tropical and temperate regions (Tejerina-Garro et al., 2005), other factors can also influence this variable. In water courses with reduced flow, like those sampled in this study, energy exchanges between the water mass, solar radiation, and air conductivity result in rapid oscillations of water temperature (Walling & Webb, 1992) in high and low water seasons. This process can be amplified by the absence of riparian vegetation (Castro & Casatti, 1997), similar to that observed in various water courses sampled. The characins Bryconamericus sp. and the armored catfish Hypostomus ancistroides and Hypostomus sp. C were associated with warm waters during the high water season (average = 23.92°C). However, the presence of bottom-dwelling fish species like the armored catfish of the genus Hypostomus (Moyle & Cech, 1996; Santos et al., 2004) could also be related to changes in benthic habitat conditions, because of an increase in water velocity and volume during the high water season. These species are sensitive to benthic habitat changes because they have specific reproduction and feeding requirements (Casatti, 2004).
Sampling sites along the watershed were separated into two groups that were most evident during low water. In both seasons, fish assemblage was related to high values of water conductivity, CDO, and turbidity registered in sampling stations 2–6, 8, 21, and 25.
Increased water volume during high water season favored the redistribution and dispersion of adult and young individuals, larvae, and eggs between sections and compartments (e. g., lake and main channel) of an aquatic ecosystem (Rodríguez & Lewis, 1994), and homogenized water physicochemical characteristics, even in areas subjected to distinct levels of anthropogenic impacts and at different spatial extents (Thomaz et al., 2007). This seems to explain the similar co-structure of fish assemblages and environmental variables observed between sampling sites located in the upper (1–8), middle (19–22) and lower (29) sections of the watershed during the high water season.
However, during low water, differences between sections can be accentuated, not only by natural phenomena such as water volume reduction, but also by the effects of anthropogenic activities. In creeks of the upper Paraná River basin not impacted by anthropogenic activities such as sewage or agriculture, conductivity diminishes from low to high water season, whereas turbidity displays the opposite trend (Carmo et al., 2005). But, this was not the case for conductivity in sampling stations 2–6, 21 and 25 of this study. In the upper section of the Meia Ponte River basin, where sampling stations 2–6 are located, an intensive process of erosion is occurring, associated with the pluvial water waste drainage system, agriculture, ranching, and deforestation (Rubin et al., 2005). These anthropogenic activities may contribute during both seasons to increase conductivity and CDO because of input of organic matter, and its later decomposition by heterotrophic organisms (Matthews, 1998). These environmental conditions appeared to favor the presence of bottom-dwelling (e.g., armored catfish Hypostomus ancistroides and catfish Cetopsorhamdia iheringi), and benthopelagic species (e. g., characins Astyanax altiparanae and catfish Rhamdia quelen) (Froese & Pauly, 2006). However, the presence of the two potamodromous characins Astyanax eigenmanniorum and Cyphocharax modestus (Froese & Pauly, 2006) could be related to migratory patterns.
Conclusion
Among the 14 environmental variables considered in this study, pH, water temperature, conductivity, CDO, and turbidity played important roles in structuring fish assemblages. Contrary to what was expected, this co-structure was regulated by seasonality (low and high water seasons) but influenced by biological patterns (fish habitat preferences, feeding, and spawning) and by anthropogenic activities (domestic sewage, urban areas, ranching,and agriculture). This situation is demonstrated not only by the difference in co-structures (fish assemblage and physicochemical variables) observed during low and high water, but also in the variation among sampling stations along the watershed. The latter suggests that the degree of influence of environmental impacts on fish-habitat relationship depends on duration and position within a watershed.
The findings of this study may be useful in approaches aiming the quantification of the quality of habitats and monitoring programs involving fish fauna. Consequently, they could serve as a support for management and restoration efforts in Neotropical regions.
References
Agostinho, A. A. & M. Zalewski, 1995. The dependence of fish community structure and dynamics on floodplain and riparian ecotone zone in Paraná River, Brazil. Hydrobiologia 303: 141–148.
Araújo, E. S. & I. A. H. Pfrimer, 2005. Hidrogeoquímica no entorno de Goiânia (Rios Meia Ponte, João Leite e Santo Antônio): um estudo comparativo da qualidade da água. Estudos 32: 103–113.
Araújo, F. G., 1996. Composição e estrutura da comunidade de peixes do médio e baixo rio Paraíba do Sul. Revista Brasileira de Biologia 56: 111–126.
Balon, E. K., 1993. Dynamics of biodiversity and mechanisms of change: A plea for balanced attention to form creation and extinction. Biological Conservation 66: 5–16.
Barnes, N. J., 1998. Chronic pollution in freshwaters, evaluation of ecological impacts and implications for water resource management. Environmental Management and Health 9: 49–53.
Böhlke, J. E., S. H. Weitzman & N. A. Menezes, 1978. Estado atual da sistemática de peixes de água doce da América do Sul. Acta Amazônica 8: 657–677.
Camargo, A. F. M., R. A. R. Ferreira, A. Schiavetti & L. M. Bini, 1996. Influence of physiograph and human activity on limnological characteristics of lotic ecosystems of the south coast of São Paulo, Brazil. Acta Limnologica Brasiliensis 8: 231–243.
do Carmo, M. S., G. R. Boaventura & E. C. Oliveira, 2005. Geoquímica das águas da bacia hidrográfica do Rio Descoberto, Brasília/DF – Brasil. Química Nova 28: 565–574.
Casatti, L. & R. M. C. Castro, 1998. Fish community of the São Francisco River headwaters riffles, southeastern Brazil. Ichthyological Exploration of Freshwaters 9: 229–242.
Casatti, L., 2004. Ichthyofauna of two streams (silted and reference) in the Upper Paraná River basin, Southeastern Brazil. Brazilian Journal of Biology 64: 757–765.
Castro, R. M. C. & L. Casatti, 1997. The fish fauna from a small forest stream of the upper Paraná river basin, southern Brazil. Ichthyological Exploration of Freshwaters 7: 337–352.
Davis W. S., 1995. Biological Assessment and Criteria: Building on the Past. In Davis, W. S. & T. P. Simon (eds), Biological Assessment And Criteria: Tools For Water Resource Planning And Decision Making. CRC Press, Boca Raton, Florida: 15–30.
de Bruyn, A. M. H., D. J. Marcogliese & J. B. Rasmussen, 2003. The role of sewage in a large river food web. Canadian Journal of Fisheries and Aquatic Sciences 60: 1332–1344.
Dei Tos, C., G. Barbieri, A. A. Agostinho, L. C. Gomes & H. I. Suzuki, 2002. Ecology of Pimelodus maculatus (Siluriformes) in the Corumbá Resevoir, Brazil. Cybium 26: 275–282.
Dolédec, S. & D. Chessel, 1994. Co-inertia analysis: An alternative method for studying species-environment relationships. Freshwater Biology 31: 277–294.
Duarte, S., F. G. Araújo, 2001. Abundância relativa e distribuição de Loricariichthys spixii (Steindachner) (Siluriformes, Loricariidade) no reservatório de Lajes, Estado do Rio de Janeiro, Brasil. Revista Brasileira de Zoologia 18: 465–477.
Ferreira, A. A., A. P. O. de Nuñer & J. R. Esquivel, 2001. Influência do pH sobre ovos e larvas de jundiá, Rhamdia quelen (Osteichthyes, Siluriformes). Acta Scientiarum 23: 477–481.
Fialho, A. P. & F. L. Tejerina-Garro, 2004. Peixes da bacia do Rio Meia Ponte, GO. Editora da Universidade Católica de Goiás, Goiânia.
Fialho, A. P., L. G. Oliveira, F. L. Tejerina-Garro & L. C. Gomes, 2007. Fish assemblage structure in tributaries of the Meia Ponte River, Goiás, Brazil. Neotropical Ichthyology 5: 53–60.
Froese, R. & D. Pauly, 2006. FishBase. World Wide Web electronic publication. http://www.fishbase.org.version (12/2006).
Galinkin, M., 2003. GeoGoiás 2002. Galinkin, M. (ed.). Agência Ambiental do Estado de Goiás, Fundação CEBRAC, PNUMA, SEMARH, Brasília.
Gauch Jr. H. G., 1982. Mutivariate analysis in comunity ecology. Cambridge University, London.
Giller, P. S. & B. Malmqvist, 2000. The Biology of Streams and Rivers. Oxford University Press, New York.
Gondin-Ferreira, E. J., 1993. Composição, distribuição e aspectos ecológicos da ictiofauna de um trecho do rio Trombetas, na área de influência da futura UHE Cachoeira Porteira, Estado do Pará, Brasil. Acta Amazônica 23: 1–89.
Holomuzki, J. R. & B. J. F. Biggs, 2000. Taxon-specific responses to high-flow disturbance in streams: implications for population persistence. Journal of the North American Benthological Society 19: 670–679.
Karr, J. R., 1981. Assessment of biotic integrity using fish communities. Fisheires 6: 21–27.
Lammert, M. & J. D. Allan, 1999. Assessing biotic integrity of streams: effects of scale in measuring the influence of land use/cover and habitat structure on fish and macroinvertebrates. Environmental Management 23: 257–270.
Lampert, W. & U. Sommer, 1997. Limnoecology: The Ecology of Lakes and Streams. Oxford University Press, Oxford.
Leite, M. A., E. L. G. Espínola & M. C. Calijuri, 2000. Tripton sedimentation rates in the Salto Grande reservoir (Americana, SP, Brasil): a methodological evaluation. Acta Limnologica Brasiliensis 12: 63–68.
Lowe-McConnell, R. H., 1999. Estudos ecológicos de comunidades de peixes tropicais. Editora da Universidade de São Paulo, São Paulo.
Lyons, J., S. Navarro-Pérez, P. A. Cochran, E. Santana & M. Guzman-Arroyo, 1995. Index of biotic integrity based on fish assemblages for the conservation of streams and rivers in west-central Mexico. Conservation Biology 9: 569–584.
Matthews, W. J., 1998. Patterns in Freshwater Fisher Ecology. International Thomson Editors, New York.
de Mérona, B. & D. Gascuel, 1993. The effects of flood regime and fishing effort on the overall abundance of an exploited fish community in the Amazon floodplain. Aquatic Living Resources 6: 97–108.
Moyle, P. B. & J. J. Cech Jr., 1996. Fishes: An Introduction to Ichthyology. Prentice Hall, New Jersey.
Moyle, P. B., 1995. Conservation of native freshwater fishes in the Mediterranean-type climate of California, USA: A review. Biological Conservation 72: 271–279.
Ometo, J. P. H. B., L. A. Martinelli, M. V. Ballester, A. Gessner, A. V. Krusche, R. L. Victoria & M. Williams, 2000. Effects of land use on water chemistry and macroinvertebrates in two streams of the Piracicaba River Basin, southeast Brazil. Freshwater Biology 44: 327–337.
Paul, J. M. & J. L. Meyer, 2001. Streams in the urban landscape. Annual Review of Ecology and Systematics 32: 333–365.
Penczak, T., A. A. Agostinho & E. K. Okada, 1994. Fish diversity and community structure in two tributaries of the Paraná River, Paraná state, Brazil. Hydrobiologia 294: 243–251.
Poff, N. L., 1997. Landscape filters and species traits: Towards mechanistic understanding and prediction in stream ecology. Journal of the North American Benthological Society 16: 391–409.
Pringle, C. M., F. N. Scatena, P. Paaby-Hansen & M. Núñez-Ferrera, 2000. River Conservation in Latin America and the Caribbean. In Boon P. J., B. R. Davies & G. E. Petts (eds.), Global Perspectives on River Conservation: Science, Policy and Practice. John Wiley & Sons, Chichester: 41–78.
Ratter, J. A., J. F. Ribeiro & S. Bridgewater, 1997. The Brazilian Cerrado: Vegetation and Threats to its Biodiversity. Annals of Botany 80: 223–230.
Rodríguez, M. A. & W. M. Lewis Jr., 1994. Regulation and stability in fish assemblages of Neotropical floodplain lakes. Oecologia 99: 166–180.
Rodríguez, M. A. & W. M. Lewis Jr., 1997. Structure of fish assemblages along environmental gradients in floodplain lakes of the Orinoco river. Ecological Monographs 67: 109–128.
de Rubin, J. C. R., M. Barberi, L. D. Medrado & G. T. Carneiro, 2005. Condicionantes naturais e antrópicos dos processos erosivos no alto Rio Meia Ponte (Goiás), Brasil. Estudos 32: 41–56.
Sabater, F., B. Andréa, M. Eugênia, M. Isabel, R. Anna, W. Joanne & S. Sergi, 2000. Effects of riparian vegetation removal on nutrient retention in a Mediterranean stream. Journal of the North American Benthological Society 19: 609–620.
dos Santos, G. M., B. de Mérona, A. A. Juras & M. Jégu, 2004. Peixes do baixo rio Tocantins: 20 anos depois da Usina HidrelétricaTucuruí. Eletronorte, Brasília.
Scheiner, S. M. & J. Gurevitch, 1993. Design and Analysis of Ecological Experiments. Chapman and Hall, New York.
Schulz, U. H. & H. Martins-Junior, 2001. Astyanax fasciatus as bioindicator of water pollution of Rio dos Sinos, RS, Brazil. Brazilian Journal of Biology 61: 615–622.
Shuter, B. J., P. E. Ihssen, D. L Wales & E. J. Snucins, 1989. The effects of temperature, pH and water hardness on winter starvation of young-of-the-year smallmouth bass, Micropterus dolomieui Lacepede. Journal of Fish Biology 35: 765–780.
Soulsby, C., R. Malcolm, C. Gibbins & C. Dilks, 2001. Seasonality, water quality trends and biological responses in four streams in the Cairngorm Mountains, Scotland. Hydrology and Earth System Sciences 5: 433–450.
Tejerina-Garro, F. L. T., R. Fortín & M. A. Rodríguez, 1998. Fish community structure in relation to environmental variation in floodplain lakes of the Araguaia river, Amazon Basin. Environment Biology of Fishes. 51: 399–410.
Tejerina-Garro, F. L., M. Maldonado, C. Ibanez, D. Pont, N. Roset & T. Oberdorff, 2005. Effects of natural and anthropogenic environmental changes on riverine fish assemblages: A framework for ecological assessment of rivers. Brazilian Archives of Biology and Technology 48: 91–108.
Thioulouse, J., D. Chessel, S. Dolédec, J-M. Olivier, F. Goreaud & R. Pelessier, 2001. Ecological data analysis: Exploratory and Euclidean methods in Environmental Sciences. Version 2001. CNRS 1995–2001.
Thomaz, S. M., L. M. Bini & R. L. Bozelli, 2007. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 579: 1–13.
Walling, D. E. & B. W. Webb, 1992. Channel Morphology and Typology. In: Calow P., & G. E. Petts (eds.). The Rivers Handbook: Hydrological and Ecological Principles. Blackwell Sciences, Massachusetts.
Wiens, J. A., 2002. Riverine landscape: Taking landscape ecology into the water. Freshwater Biology 47: 501–515.
Zar, J. H., 1998. Biostatistical Analysis (4th ed.). Prentice-Hall Inc., Englewood Cliffs, New Jersey.
Acknowledgements
The authors are grateful to the Agência Ambiental de Goiás, and in particular to Rodrigo Porfírio, for their collaboration and support in both field and laboratory, undergraduate Biology students from the Catholic University of Goiás (UCG), the technical staff of the Centro de Biologia Aquática, particularly Waldeir Francisco de Menezes, Mônica Mazini, and in particular Patrick John O’Sullivan (UCG) and Anne Mary Staunton for their revision and suggestions for English style and grammar of the first draft and Dr. José Alexandre Felizola Diniz Filho (Federal University of Goiás) and two anonymous reviewers for their valuable suggestions, and Dr. Robert Hughes (Oregon State University) for his critical reading and English correction. LGO is supported by a CNPq productivity fellowship.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: S. M. Thomaz
Rights and permissions
About this article
Cite this article
Fialho, A.P., Oliveira, L.G., Tejerina-Garro, F.L. et al. Fish-habitat relationship in a tropical river under anthropogenic influences. Hydrobiologia 598, 315–324 (2008). https://doi.org/10.1007/s10750-007-9165-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-007-9165-3