
Abstract
Nutrient availability and interspecific competition may affect emergent wetland plant growth and resource allocation in constructed wetland. A glasshouse study was conducted to investigate the influence of nutrient and mixture between Canna indica Linn and Schoenoplectus validus (Vahl) A. Löve & D. Löve on their growth and resource allocation in the wetland microcosms, using simulated secondary-treated municipal wastewater effluent with either low (17.5 mg N and 10 mg P l−1) or high (35 mg N and 20 mg P l−1) nutrient concentrations. After 65 days, the high nutrient treatment stimulated plant growth and resulted in allocation of more resources to the above-ground tissues compared to below-ground ones. The concentrations of N and P in the plant tissues (except P in above-ground tissues) were significantly higher, whereas N and P use efficiencies were significantly lower in the high than the low nutrient treatment. The total biomass for C. indica in mixture increased significantly in the high nutrient treatment, but that for S. validus was significantly lower in mixture than in monoculture. Relative yield (RY) indicated that there was significant interspecific competition between S. validus and C. indica in mixtures, with C. indica being the superior competitor. The growth of S. validus was significantly inhibited by the presence of C. indica in their mixture. Compared with monoculture, S. validus in mixture had significantly higher percentages of root biomass and allocations of N and P to roots, whereas C. indica was not significantly affected by mixture. The results suggested that the growth and resource allocation of C. indica and S. validus could be altered by nutrient availability and interspecific competition in constructed wetlands.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The use of constructed wetlands for tertiary water purification of municipal wastewater has been received an increasing attention in recent years. Constructed wetlands offer effective and reliable treatment of wastewater in a simple and inexpensive manner (Sundaravadivel & Vigneswaran, 2001).
Wetland plants contribute to the function of wetland systems as habitat providers, bioengineers to promote sedimentation and even dispersion of flow, and erosion control and nutrient/pollutant transformers (Kadlec & Knight, 1996). This latter attribute relates to the rapid growth rates of species in resource-rich environments, and ability to concentrate luxury amounts of nutrients in their above- and below-ground biomass. The partitioning of nutrients between shoots and roots/rhizomes varies between species and seasons. In small, lightly loaded wastewater treatment systems, plant uptake can be the principal form of nutrient removal, often accounting for up to 80% of the nutrient pool (Kadlec & Knight 1996). Plants are also the principal nutrient sinks during the initial years of establishment in the wetland. The removal of plant material through a harvest reduces the potential for biologically-assimilated nutrients being remobilised into the wetland system. Harvesting can also encourage large nutrient uptake by the plants during the rapid growth and recovery of the harvested plants. Therefore, the function of plants in nutrient stripping is a dynamic one, and requires an understanding of plant tolerances and nutrient requirements to optimize performance in terms of uptake.
The concentrations of nutrients (N and P) in the effluents of the wastewater and loading rate to the constructed wetlands vary at different times during the year and for different wastewater treatment plants. On the other side, nutrient availability could affect plant growth responses and resource allocations. Plant not only grow at a slow rate at low nutrient supply compared with high nutrient supply, but also increase their biomass allocation to roots (Poorter & Nagel, 2000) and reduce the nutrient concentrations in their biomass (Aerts & Chapin, 2000).
Improvements in plant selection and cultivation might make the constructed wetlands more efficient for nutrient removal from the wastewater. It is not clear if it is desirable to maintain a single plant species, or a mix of plant species, in constructed wetland. But single plant (monoculture) systems are more susceptible to plant death due to predation or disease. Therefore, it is generally assumed that multiple plant (mixture) and native plant systems are more resilient than monocultures (EPA, 2000). It is well known that positive, negative or indifferent interrelationship may occur between plants of different species. The results of such competition might cause the preferential establishment and growth of certain species, and/or the suppression and extinction of other species (Agami & Reddy, 1990). In recent years, several studies have been reported on the interspecific competition for emergent wetland species. For example, Wetzel and van der Valk (1998) found that Phalaris arundinacea is an inherently better competitor than Carex stricta or Typha latifolia. Coleman et al. (2001) observed that Typha latifolia was the superior competitor among the three-species (Juncus effusus, Typha latifolia and Scirpus cyperinus) mixture in small-scale constructed wetlands. In plant mixtures consisting of Carex flava, Centaurea angustifolia, Lycopus europaeus and Selinum carcifolia grown in the sand culture with different N:P supply ratios and different total supplies of N and P, Lycopus europaeus performed best at low and intermediate N:P ratios, and Carex flava at high N:P ratio (Güsewell & Bollens, 2003). However, few studies have investigated the competitive impact between species with different growth forms or significantly different morphologies.
Schoenoplectus validus (Vahl) A. Löve & D. Löve and closely related rush species S. lacustris, S. acutus and S. californicus have been used widely in constructed wetlands around the world (Tanner, 2001). S. validus is a tall, perennial, herbaceous rush. The leaves are reduced to sheaths at the base of the stems. It usually occurs in poorly drained soil, grows better in saline conditions than in fresh water and tolerates a wide range of salinity. Similarly, ornamental species, Canna indica Linn, has been used to treat (i) septic tank effluent in a simulated vertical-flow constructed wetlands (Zhu et al., 2004); (ii) domestic wastewater in a medium-scale vertical/reverse-vertical flow constructed wetland (Yue et al., 2004); and (iii) municipal wastewater in a full-scale subsurface-flow constructed wetland (Shi et al., 2004). C. indica is an upright perennial rhizomatous herb. The leaves are elliptic and rather fleshy. It is a native plant of tropical America, and very popular ornamental plant throughout the tropical and subtropical region around the world, but grows in thickets, crowding out other plants and difficult to remove due to its spread by rhizomes.
Although the two plant species and their closely related species have been selected and planted in the various constructed wetlands for the improvement of water quality and landscape restoration under mono- or mixed-culture conditions, due to their relative high nutrient removal efficiency and aesthetical value (Calheiros et al., 2007; Fu et al., 2006; Grosse et al., 2001; Wu and Ding, 2006; Wu et al., 2006), no research has been done on dealing with interspecific competition between C. indica and S. validus. The objective of this study was to investigate the influence of nutrient availability and mixture between C. indica and S. validus on their growth and resource allocation using simulated secondary-treated municipal wastewater in the wetland microcosms.
Materials and methods
Concentrations of N and P in secondary-treated municipal wastewater effluent
The data on quality of discharge water from secondary-treated municipal wastewater effluent collected at Subiaco Waste Water Treatment Plant in Shenton Park, Perth, Western Australia from July 2002 to November 2004 were obtained from Water Corporation, Western Australia. The yearly ranges of the total N and P concentrations in the wastewater effluent were from 8.2 to 28 mg N l−1and 5.6 to 17 mg P l−1; and the average concentrations between 1 November 2003 to 31 October 2004 were 17 mg N l−1 and 10 mg P L−1.
Wetland microcosms
Thirty-six microcosms were used as experimental units in this study. A wetland microcosm was established in 33-l plastic containers (0.39 × 0.29 × 0.30 m) with a hole fitted with a plastic tube close to the base to facilitate drainage of water. A mesh covering the hole was fastened on the inside of the microcosm to prevent loss of sand during the water drainage. Approximately 25 kg of washed river sand (<4 mm diameter) was added to each container, giving a sand depth of about 0.15 m.
Experimental setup
The experiment was conducted in a phytotron at the University of Western Australia with controlled day/night temperatures of 25/20°C under natural light conditions from the beginning of June to the later September. A complete randomized block factorial design (four plant culture treatments × two nutrient concentration treatments) with three replicates was employed.
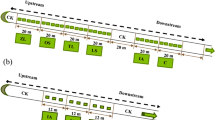
At the establishment of plant treatment, the four plant culture treatments with nine replicates were: (A) control (with no plants added); (B) monoculture of C. indica; (C) monoculture of S. validus; and (D) mixture between C. indica and S. validus. The seedlings of C. indica and S. validus were purchased from a local nursery. The healthy plants of relatively similar size were transplanted into the microcosms. Each microcosm contained six plants in two row of three: six C. indica, six S. validus or three of each species (Fig. 1). At the planting time, seedlings of C. indica were approximately 8 cm tall with 1–2 leaves, and S. validus were approximately 10 cm tall with 4–6 ramifications.

The vertical view of the design plant treatment in the microcosms. (A) control (with no plants added); (B) monoculture of C. indica; (C) monoculture of S. validus; and (D) mixture between C. indica and S. validus
In order to minimize variability in the experiment, a simulated nutrient solution was used in the microcosms at the average concentrations of N and P similar to the secondary-treated wastewater at Subiaco Waste Water Treatment Plant as described above. The solution contained 17.5 mg N l−1 (1:1 NH4–N and NO3–N) and 10 mg P l−1, and other macro- and micronutrients (mg l−1): 13 K, 10 Ca, 5 Mg, 7 S, 0.04 Zn, 0.01 Cu, 0.34 Fe, 0.05 Mn, 0.26 B and 0.05 Mo. Microcosms were filled with the nutrient solution to achieve the water depth of 0.02 m above the sand surface and renewed weekly.
After plants were grown in the microcosms for 50 days, three replicates of each plant culture treatment (including control) were harvested, and plant (shoot, rhizome and root) and substrate samples were taken. The remaining microcosms were initialized with two nutrient treatments each in three replicates: (1) low level containing nutrients as in the starting solution described above, and (2) high level with double concentrations of N and P. The concentrations of other nutrients were kept the same as in the starting solution described above, except double K. Microcosms were filled with 10 l either low or high nutrient solution to achieve the water depth of about 0.08 m above the sand surface. Each microcosm was manually drained through tube positioned at the bottom of each container and refilled every 7 days.
Sampling and measurements
Total shoot numbers and shoot heights in each microcosm were measured at approximately weekly intervals from the day after imposing nutrient treatments until the plants were harvested at the end of the experiment (65 days later). The shoot height was measured from the base of the plants to the top of the longest leaves for C. indica or the top of culms for S. validus.
To estimate above-ground biomass of plants prior to the destructive sampling, for S. validus, regression relationship between total shoot length and dry above-ground biomass were developed at the end of the experiment. For C. indica, no significant regression between shoot number and above-ground biomass was detected, so that shoot number was not included in the regression. These equations were used to estimate above-ground biomass from weekly shoot height and number measurements. The regression equation for each species was:
S. validus
C. indica
The plants were harvested after 65 days of nutrient and plant culture treatments. The shoots were cut at the sand surface, and their bases were washed to remove any adhering sediments. Each microcosm was then excavated and hand sorted into above-ground plant parts (stems and leaves, inflorescences and flowering stems), rhizomes (including stem base) and roots. The plants that had grown in mixture were separated to species. The roots were separated from the sand by washing away the sand using tap water and collecting roots onto the mesh. All plant samples were dried to constant weight at 70°C for 5 days in a forced-air oven, weighed and ground to pass a 0.75-mm mesh.
Total nitrogen in the samples of plant tissues were determined by the Dumas combustion method using an automated CN analyzer (LECO CHN-1000, LECO Company, St Joseph, Michigan, USA). Total phosphorus in plant was determined by colorimetric with spectrophotometer HITACHI U-1100 using the vanado-molybdate method after digesting material in mixture of concentrated nitric and perchloric acids (Bassett et al., 1978).
Calculations and statistical analysis
All data for calculation and analysis were obtained from the beginning of imposing nutrient treatments to the end of the experiment (65 days of plant growth). Dry biomass production was estimated from the total biomass of each microcosm divided by the area of the microcosm. The biomass allocation was characterized using the ratio of root-supported tissue (above-ground, rhizomes) to root biomass (S/R) and the ratio of above-ground to below-ground biomass (A/B) (Lorenzen et al., 2001). Resource allocation ratio was defined as the ratio of a certain tissue biomass or N, P content to total plant biomass or total plant N, P content. The allocation into above-ground, rhizomes and roots were calculated as the ratio between the biomass or nutrient content of the fraction and the total biomass or nutrient content of the plants. Nutrient use efficiency was calculated as the total dry biomass divided by total N or P content. The following biometric characteristics were estimated: relative shoot growth rate = (Ln final shoot length−Ln initial shoot length)/days, and relative increase in shoot number = (Ln final shoot number−Ln initial shoot number)/days (Tylova-Munzarova et al., 2005).
The competitive effects of species were examined by calculating the relative yield of individuals of a species when grown with another species (interspecific competition) compared to the relative yield of the species grown alone (intraspecific competition) while maintaining the same overall density. Relative yield (RY) of above-ground biomass was calculated as:
RY of species A = (yield of species A in mixture)/(yield of species A in monoculture) (Harper, 1977).
All statistical tests were performed using SPSS version 10 for windows. A set of two-way ANOVA was used to determine significance of nutrient and plant treatment effects on plant biomass and characteristics, plant concentrations of N and P, and resource allocation (the percentage after log transformation). Least significant difference (LSD) was applied to test for significance between treatment means.
Results
Plant biomass
Significant differences in the aboveground and total biomass were observed, but not in the belowground, rhizome and root biomass between the nutrient treatments and significant differences in the biomass of various plant parts were observed among the plant treatments. The significant interactions between nutrient and plant treatments were observed in the aboveground and total biomass (Table 1). After 65 days of nutrient and plant treatments, the total biomass for C. indica in mixture was significantly higher in the high nutrient treatment than in the low nutrient treatment, but that for S. validus was not significantly affected by the nutrient treatments. The above-ground biomass for C. indica in both monoculture and mixture, and that for S. validus in monoculture were significantly increased by the high nutrient treatments. The below-ground biomass for C. indica was significantly higher, whereas that for S. validus was significantly lower in mixture than in monoculture treatment (Table 1).
Species mixture remarkably influenced above-ground plant growth before imposing nutrient treatments. For both species, these early effects followed the same trends as those at harvesting time. After imposing nutrient treatments, the relative growth of C. indica in mixture increased in the high nutrient treatment, but the effect disappeared after day 40, whereas the relative growth of S. validus in mixture was not enhanced by the high nutrient treatment. Those effects in 65 days of plant growth were shown using the relative yield (RY) of above-ground biomass for C. indica and S. validus under the low and high nutrient treatments (Fig. 2). The relative yield (RY) indicated that there was significant interspecific competition between S. validus and C. indica in mixture, with C. indica being the superior competitor.

Relative yield (RY) of above-ground biomass for C. indica and S. validus under the low and high nutrient treatments after imposing the nutrient treatments. The high nutrient treatment was imposed after transplanting plants of 50 days. A RY of 1 indicates that plants were the same size in mixture as in monoculture. RY <1 indicates that plants were smaller in mixture than in monoculture and RY >1 indicates that plants were larger in mixture than in monoculture
Biomass allocation and biometric characteristics
Biomass allocation and plant characteristics (except relative rate of shoot growth among the plant treatments) were significantly affected by the nutrient and plant treatments, but no significant interaction between nutrient and plant treatments was detected (Table 2). Plants had significantly higher percentage of above-ground biomass, relative rate of shoot growth and relative increase in numbers of shoots in the high than the low nutrient treatment. Compared with monoculture, S. validus in mixture had significantly lower A/B ratio, percentage of above-ground biomass and relative increase in numbers of shoots under the low nutrient treatment, and S/R ratio and relative increase in numbers of shoots under the high nutrient treatment, whereas C. indica was not significantly affected by mixtures. Compared with C. indica, S. validus had more biomass allocated into above-ground tissues (Table 3).
Concentrations of N and P in plant tissues
The concentrations of N and P in the plant tissues were significantly influenced by nutrient and plant treatments after 65 days of plant growth. The concentrations of N and P (except P in above-ground tissues) were significantly higher in the high nutrient than in the low nutrient treatment. The concentrations of N and P in plant tissues (except rhizome) were significantly different in plant treatments. The concentrations of N and P in the roots of S. validus were significantly lower in mixed-culture than in monoculture in the high nutrient treatment (Table 4).
Nutrient allocation
Plant nutrient (N and P) allocations and N/P ratios (except N/P ratios in roots between nutrient treatments) were significantly affected by nutrient and plant treatments, and significant interactions between nutrient and plant treatments were detected in N allocation to rhizome and P allocation to above-ground organ and rhizome (Table 5). Plants had significantly higher relative allocation of N and P to aboveground organs in the high than the low nutrient treatment. Compared with monoculture, S. validus in mixture had significantly higher relative allocation of N and P to roots (except P allocation to root under the high nutrient treatment) and significantly lower N/P ratios in above-ground tissues and rhizomes, whereas C. indica was not significantly affected by mixtures (Table 6).
Nutrient use efficiency
N and P use efficiency of C. indica and S. validus were significantly affected by the nutrient treatments, but not by the plant treatments. No significant interaction was detected between nutrient and plant treatments (Table 7). N and P use efficiency were significantly higher in the low nutrient treatment (86 g dry weight g−1 N and 474 g dry weight g−1 P, averaged over the plant treatments) than in the high nutrient treatment (61 g dry weight g−1 N and 404 g dry weight g−1 P, averaged over the plant treatments).
Discussion
Resource allocation
Aquatic plants can take up large quantities of nutrients, and even assimilate them luxuriously (Cronk & Fennessy, 2001). The present results showed the plants were capable of taking up more N and P (Table 4) and producing more biomass (Table 1) in the high than the low nutrient treatment. Differences between species in biomass accumulation, and tissue N and P concentrations are likely to reflect species and developmental stage differences in efficiency of nutrient uptake and use (Tanner, 1996; Güsewell & Bollens, 2003). In the present study, significant differences in the nutrient use efficiency were not detected among the plant treatments (Table 7). Therefore, the differences in biomass and nutrient concentration might be related to the nutrient uptake efficiency. Although the below-ground biomass was relatively high in the high nutrient treatment compared with the low nutrient treatment, significant difference in the below-ground biomass was not observed between the nutrient treatments. This might be due to the relatively high plant density and small size of the microcosms, which limited below-ground plant growth in the high nutrient treatment. Güsewell and Bollens (2003) also pointed out that the total below-ground biomass was less responsive to nutrient treatments than the above-ground biomass, with an inconsistent effect of the N/P supply ratio in the pot experiments. The total below-ground biomass was higher in the intermediate nutrient supply than the high nutrient supply at 15 N/P supply ratio.
Plant productivity and resource allocation varied widely between C. indica and S. validus (Tables 1, 3 and 6). This variation is likely to arise from relative differences in initial propagule vigor, as well as from intrinsic species and possibly ecotype growth characteristics (Daniels, 1991). However, factors such as the physiological and developmental state of the propagules are likely to have been of more importance than the biomass of the propagules per se (Tanner, 1996).
The A/B ratios of the species in the present study ranged from 0.6 to 1.7. These values were in the ranges of other aquatic plants (Hogetu, 1984). Plants alter their resource allocation to above-ground and below-ground tissues along environmental gradients of disturbance and resource availability (Kirkman & Sharitz, 1993; Stuart et al., 1999). In the present study, the resource allocations were altered by nutrient availability, and also changed for S. validus by the mixture. The growth of S. validus in mixture was strongly inhibited by the presence of C. indica, and allocated more percentage biomass to its below-ground tissues. Wu & Yu (2004) found that Nymphoides peltata decreased the ratios of above-ground to below-ground biomass with increasing density of Zizania latifolia in mixture.
Resource allocation is known to change during the growing season for most plant species (Aerts et al., 1992). The resource allocation patterns observed in the present experiment may only reflect the short length of the experiment. It is important to note that while a microcosm trial enables more control over experimental conditions than field trials, there are significant differences in the temporal and spatial aspects of such studies. Thus the present results of the species combination, and the relatively short duration of the growth trials in relation to the life cycles of these clonal species, care must be exercised, in attempting to generalize the results from the microcosm to field-scale constructed wetland.
Competition in mixture
Grime and Hodgson (1987) listed characteristics of species with high competitive ability: (1) a robust perennial life form with a strong capacity to ramify vegetatively, (2) the rapid commitment of captured resources to the construction of new leaves and roots, (3) high morphological plasticity during the differentiation of leaves and roots, and (4) short life spans of individual leaves and roots. Both C. indica and S. validus are robust perennials, which rapidly produce ramets and have high growth rates. In addition, C. indica produces large storage rhizomes and high growth rates have been measured (Zhao et al., 2003).
Wetzel and van der Valk (1998) pointed out that rapid growth was not the only factor, and suggested that plant architecture played a significant role in competition between Carex stricta, Phalaris arundinacea and Typha latifolia. Morphological characteristic of a plant affecting competition for light have been reported in agricultural and woody plants (McLachlan et al., 1993; Sipe & Bazzaz, 1994; Webster et al., 1994). Species with different morphologies showed large differences in canopy structure. A grass, having a more open canopy, was consistently a weak competitor when grown with forbs (Tremmel & Bazzaz, 1993). The morphological characteristics, such as tall shoot, leaf shape and large canopy diameter, were significantly correlated with increased competitive ability in wetland plants (Gaudet & Keddy, 1988). Changes of water levels, and the presence/absence of competitor species, produced significant morphological responses in mature individuals of five freshwater wetland plant species: Agrostis stolonifera L., Carex rostrata Stokes, Deschampsia cespitosa (L.) Beauv., Filipendula ulmaria (L.), Phalaris arundinacea (L.). These responses provide evidence for potential advantages in survival and ability to spread vegetatively (Kennedy et al., 2003). In the present study, C. indica and S. validus differ substantially in their growth rate, morphology, physiology and size. It is possible that C. indica, having large leaf areas and canopy diameter, maximized the capture of light and nutrient resources by maximizing vegetative growth under both nutrient availabilities and out-competed S. validus in mixture.
Interspecific competition is often regarded as being caused by mutual exploitation of limiting resources (resource consumption, including light interception by plants and space occupancy by space-limited sessile organisms), by the production of toxins, and by various combinations of these mechanisms (Tilman, 1987). In the present results, however, it is unknown whether interspecific competition between C. indica and S. validus was caused by single mechanism or the combinations of above mentioned mechanisms. It is worth mentioning that there were significant differences in pH of the effluents among the plant treatments. The lowest effluent pH was detected in microcosms with monoculture of C. indica, followed by mixture, but no significant difference was found between monoculture of S. validus and the treatment without plants (see Zhang et al., 2007).
Conclusions
High nutrient availability improved plant growth and resulted in allocation of more resources to the above-ground tissues compared to below-ground ones. Due to interspecific competition, the growth and resource allocation of S. validus were significantly influenced by mixture, but C. indica was less affected. The results suggested that the plant growth and resource allocation of C. indica and S. validus could be altered by nutrient availability and interspecific competition in the constructed wetlands. To enhance the aesthetic appeal of constructed wetlands, but avoid the interspecific competition, the intensive studies on nutrient uptake for each plant species and their mixtures at various nutrient concentrations and at various planting densities are needed in both laboratory and field conditions.
References
Aerts, R. & F. S. Chapin, 2000. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research 30: 1–67.
Aerts, R., H. de Caluwe & H. Konings, 1992. Seasonal allocation of biomass and nitrogen in four Carex species from mesotrophic and eutrophic fens as affected by nitrogen supply. Journal of Ecology 80: 653–664.
Agami, M. & K. R. Reddy, 1990. Competition for space between Eichhornia crassipes (Mart.) Solms and Pistia stratiotes L. cultured in nutrient-enriched water Aquatic Botany 38: 195–208.
Bassett, J., R. C. Denney, G. H. Jeffery & J. Mendham, 1978. Vogel’s Textbook of Quantitative Inorganic Analysis Including Elementary Instrumental Analysis. 4th edition. Longman, London, New York.
Calheiros, C. S. C., A. O. S. S. Rangel & P. M. L. Castro, 2007. Constructed wetland systems vegetated with different plants applied to the treatment of tannery wastewater. Water Research doi:10.1016/j.watres.2007.01.012.
Coleman, J., K. Hench, K. Garbutt, A. Sextone, G. Bissonnette & J. Skousen, 2001. Treatment of domestic wastewater by three wetland plant species in constructed wetlands. Water, Air and Soil Pollution 128: 283–295.
Cronk, J. K. & M. S. Fennessy, 2001. Wetland plants: Biology and Ecology. Lewis Publisher, Boca Raton, FL, USA: 462.
Daniels, R. E., 1991. Variation in the performance of Phragmites australis in experimental culture. Aquatic Botany 42: 41–48.
EPA, 2000. U.S. Environmental Protection Agency, Manual, Constructed Wetlands Treatment of Municipal Wastewaters. EPA/625/R-99/010. Cincinnati, OH.
Fu, C., Y. Tang, X. Chen, Y. Xu & G. Xing, 2006. Effects of purification highly salty reuse water quality by three plants in TEDA landscape river. Journal of Chongqing University (Natural Science Edition) 29: 118–121 (in Chinese, with English abstract).
Gaudet, C. L. & P. A. Keddy, 1988. A comparative approach to predicting competitive ability from plant traits. Nature 334: 242–243.
Grime, J. P. & J. G. Hodgson, 1987. Botanical contributions to contemporary ecological theory. New Phytologist 106(Suppl.), 283–295.
Grosse, W., F.W. Wissing, R. Perfler, Z. Wu, J. Chang & Z. Lei, 2001. Water Quality Improvement in Tropical and Subtropical Areas for Reuse and Rehabilitation of Aquatic Ecosystem. In Gawlik B.M., B. Platzer & H. Muntau (eds), Freshwater Contamination in China. Office for official publications of the European Communities, European Communities: 19–32.
Güsewell, S. & U. Bollens, 2003. Composition of plant species mixtures grown at various N:P ratios and levels of nutrient supply. Basic and Applied Ecology 4: 453–466.
Harper, J. L., 1977. Population Biology of Plants. Academic Press, New York.
Hogetu, K., 1984. Bioeconomics. Syokabo Press. Tokyo: 245.
Kadlec, R. H. & R. L. Knight, 1996. Treatment Wetlands. Lewis Publisher, Boca Raton, FL, USA: 893.
Kennedy, M. P., J. M. Milne & K. J. Murphy, 2003. Growth responses to groundwater level variation and competition in freshwater wetland plant species. Wetlands Ecology and Management 11: 383–396.
Kirkman, L. K. & R. R. Sharitz, 1993. Growth in controlled water regimes of three grasses common in freshwater wetlands of the southeastern USA. Aquatic Botany 44: 345–359.
Lorenzen, B., H. Brix, I. A. Mendelssohn, K. L. McKee & S. L. Miao, 2001. Growth, biomass allocation and nutrient use efficiency in Cladium jamaicense and Typha domingensis as affected by phosphorus and oxygen availability. Aquatic Botany 70: 117–133.
McLachlan, S. M., M. Tollenaar, C. J. Swanton & S. F. Weise, 1993. Effect of corn-induced shading on dry matter accumulation, distribution, and architecture of redroot pigweed (Amaranthus retroflexus). Weed Science 41: 568–573.
Poorter, H. & O. Nagel, 2000. The role of biomass allocation in the growth response of plants to different levels of light, CO2, Nutrients and water: a quantitative review. Australian Journal of Plant Physiology 27: 595–607. .
Shi, L., B. Wang, X. Cao, J. Wang, Z. Lei, Z. Wang, Z. Liu & B. Lu, 2004. Performance of a subsurface-flow constructed wetland in southern China. Journal of Environmental Sciences 16: 476–481.
Sipe, T. W. & F. A. Bazzaz, 1994. Gap partitioning among Maples (Acer) in central New England: shoot architecture and photosynthesis. Ecology 75: 2318–2332.
Stuart, J. B., G. G. Georage & K. F. Walker, 1999. Growth and resource allocation in response to flooding in the emerged sedge Bolboschoenus medianus. Aquatic Botany 63: 145–160.
Sundaravadivel, M. & S. Vigneswaran, 2001. Constructed wetlands for wastewater treatment. CRC Critical Reviews in Environmental Science and Technology 31: 351–409.
Tanner, C. C., 1996. Plants for constructed wetland treatment systems–a comparison of the growth and nutrient uptake of eight emergent species. Ecology Engineering 7: 59–83.
Tanner, C. C., 2001. Growth and nutrient dynamics of soft-stem bulrush in constructed wetland treating nutrient-rich wastewaters. Wetland Ecology and Management 9: 49–73.
Tilman, D., 1987. The importance of the mechanisms of interspecific competition. The American Naturalist 129: 769–774.
Tremmel, D. C. & F. A. Bazzaz, 1993. How neighbour canopy architecture affects target plant performance. Ecology 74: 2114–2124.
Tylova-Munzarova, E., B. Lorenzen, H. Brix & O. Votrubova, 2005. The effects of NH +4 and NO −3 on growth, resource allocation and nitrogen uptake kinetics of Phragmites australis and Glyceria maxima. Aquatic Botany 81: 326–342.
Webster, T. M., M. M. Loux, E. E. Regnier & S. K. Harrison, 1994. Giant ragweed (Ambrosia trifida) canopy architecture and interference studies in Soybean (Glycine max). Weed Technology 8: 559–564.
Wetzel, P. R. & A. G. van der Valk, 1998. Effect of nutrient and soil moisture on competition between Phalaris arundinacea, Carex stricta and Typha latifolia. Plant Ecology 138: 179–190.
Wu, J. & L. Ding, 2006. Study on treatment of polluted river water using pilot-scale surface flow constructed wetlands system. Environmental Pollution and Control 28: 432–434 (in Chinese, with English abstract).
Wu, J., S. Huang, X. Ruan & L. Ding, 2006. Treatment of polluted river water using surface flow constructed wetlands in Xinyi river floodplain, Jiangsu Province. Journal of Lake Sciences 18: 238–242 (in Chinese, with English abstract).
Wu, Z. & D. Yu, 2004. The effects of competition on growth and biomass allocation in Nymphoides peltata (Gmel.) O. Kuntze growing in microcosm. Hydrobiologia 527: 241–250.
Yue, C. L., J. Chang, Y. Ge, & Y. M. Zhu, 2004. Treatment efficiency of domestic wastewater by vertical/reverse-vertical flow constructed wetlands. Fresenius Environmental Bulletin 13: 505–507.
Zhang, Z., Z. Rengel & K. Meney, 2007. Nutrient removal from simulated wastewater using Canna indica and Schoenoplectus validus in mono- and mixed-culture in wetland microcosms. Water, Air and Soil Pollution DOI 10.1007/s11270–007–9359–3.
Zhao, J., Q. Yang, Z. Chen & Z. Huang, 2003. Studies on root system biomass of the plants in several kinds of wetland. China Environmental Science 23: 290–294 (in Chinese, with English abstract).
Zhu, X., L. Cui, W. Liu & Y. Liu, 2004. Removal efficiencies of septic tank effluent by simulating vertical-flow constructed Canna indica Linn wetlands. Journal of Agro-Environment Science 23: 761–765 (in Chinese, with English abstract).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zhang, Z., Rengel, Z. & Meney, K. Growth and resource allocation of Canna indica and Schoenoplectus validus as affected by interspecific competition and nutrient availability. Hydrobiologia 589, 235–248 (2007). https://doi.org/10.1007/s10750-007-0733-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-007-0733-3