
Abstract
Alpha, beta and gamma are three components of species diversity. Knowing these attributes in floodplain lake phytoplankton communities is vital when selecting conservation areas. Species diversity is commonly used with other taxonomic groups, but rarely with phytoplankton. We compared the number of phytoplankton species (alpha diversity) from 21 Middle Araguaia River floodplain lakes in the 2000 and 2001 rainy and dry seasons. From these samples we estimated complete survey species richness (gamma diversity), quantified differences in species composition between lakes (beta diversity) and assessed the influence of abiotic variables on beta diversity. We recorded a total of 577 taxa. The Sjack1 estimator indicated that 62.31% of taxa were sampled in the 2000 rainy and 67.65% dry seasons, and 68.36% in the 2001 rainy and 73.5% dry seasons. In almost all seasons, alpha diversity negatively correlated with latitude. Beta diversity (β-1) was higher in high water periods, especially in 2000. This may have been caused by isolated heavy rainfall, which would have increased environmental heterogeneity and raised beta diversity. DCA showed differences in phytoplankton composition between rainy and dry seasons in 2000 and 2001, reflecting the influence of flood pulses on phytoplankton composition. The Mantel test indicated spatial distribution patterns where geographically more distant lakes had less-similar phytoplankton communities.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
In tropical regions, river floodplain systems exhibit significant physical, chemical, and biological temporal variations. According to the “flood pulse” theory (Junk et al., 1989), such variations are mainly associated with changes in water level.
Species richness is the simplest measurement to quantify and express the complexity of a given area. According to Gaston (1996), the number of species recorded reflects sampling effort. Beta diversity provides a new perspective on analyzing biodiversity in a river-floodplain system (Ward et al., 1999). It is defined as the change in species composition along environmental gradients (Neiff, 1996). In floodplain lakes, high environmental heterogeneity (differences in origin, morphometry, hydrology, and limnology) is expected to cause high beta diversity (Neiff, 1996). Because of the homogenizing effect of flood pulses, which cause increased similarity between lakes (Thomaz et al., 1997), we expected decreased beta diversity during rainy seasons.
The aim of this study was to compare the number of phytoplankton species (alpha diversity) of 21 floodplain lakes of the Middle Araguaia River during periods of high (rainy season) and low (dry season) waters in the years 2000 and 2001. We estimated species richness for the whole survey (gamma diversity), quantified differences in species between lakes (beta diversity), and investigated the influence of abiotic variables on beta diversity.
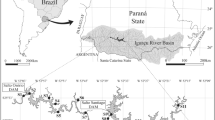
The Araguaia River, located in central Brazil has its headwaters in the Serra dos Caiapós, and has a large number of floodplain lakes. We investigated 21 of these lakes on a 240 km stretch of the river between 13°10′ & 14°50′ S, and 50°35′ & 51°05′ W. Details of the geographical coordinates are presented in Nabout et al. (2006). Field observations demonstrate that the lakes have considerably different geomorphological origins and levels of human impact. Pinheiro (2004) studied 20 of these 21 lakes and reported that lakes 6, 7, 8, 9, 10, 12, 13, 14, 15, 16, 18, 20, and 21 are abandoned channel (oxbow), lakes 2, 3, and 4 are oxbow chain, lake 5 is a meander scroll, and lakes 1, 11, and 17 are blocked valley; lake 19 was not classified.
Subsurface plankton samples were collected in the rainy (February 2000, March 2001) and dry (August 2000, September 2001) seasons. We analyzed both quantitative and qualitative samples to estimate total richness. Subsamples were placed in 100 ml dark bottles and fixed with a modified lugol-acetic solution for quantitative analyses (Vollenweider, 1974) and Transeau solution for qualitative analyses (Bicudo & Menezes, 2005). We adopted the Van den Hoeck et al. (1993) taxonomic classification system.
Limnological features of the Middle Araguaia River floodplain lakes were described by Nabout et al. (2006).
To estimate phytoplankton species richness of the complete survey area (gamma diversity), we used the nonparametric extrapolating index. This index is based on incidence (presence/absence) data (Chazdon et al., 1998). We used the first-order Jackknife estimator (Sjack1; Burnham & Overton, 1978) and calculations were made using the EstimateS program (Colwell, 1997).
Beta diversity indices were also applied to quantify species composition turnover. We used β-1 index (Harrinson et al., 1982) for groups of the 21 lakes along the Middle Araguaia River.
ANOVA (significance level 0.05) was used to test for limnological differences between the 2000 and 2001 rainy and dry seasons, and to determine species richness difference (alpha diversity) between the different types of lakes (abandoned channel or oxbow, oxbow chain, blocked valley). The meander scroll lake (lake 5) was excluded from the analysis, because it was sampled once only; lake 19 was also excluded because it was not classified. Linear regression analysis (significance level 0.05) was used to model the relationships between alpha diversity (richness of each lake) and latitude. Variables were log-transformed when necessary to meet linear regression assumptions (Sokal & Rohlf, 1981).
Detrended correspondence analysis (DCA; Hill & Gauch, 1980) a multivariate approach was also applied to evaluate the similarity among the species composition of the lakes in different seasons. DCA was used with the option of downweighting rare species, and calculations were made using the PC-ORD program (McCune & Mefford, 1997). We tested spatial autocorrelation by the Mantel test (Mantel, 1967), based on 10,000 randomizations. The first matrix was composed of the Jaccard dissimilarity (1-Jaccard similarity) between lakes. The second matrix expressed geographical distances between the lakes.
ANOVA indicated significant differences between the rainy and dry season in 2000 for pH, water temperature, oxygen saturation, transparency and depth; and for, nitrogen, total phosphorus, water temperature, oxygen saturation, transparency and depth in 2001 (Table 1).
The lakes showed limnological differences between the 2000 and 2001 rainy and dry seasons; there were also between-year differences. In the 2000 rainy season the lakes were deeper; in the dry season, they had higher transparency, conductivity, oxygen saturation, and water temperature. In 2001, they were also deeper and had higher transparency and water temperature; in the dry season the lakes had higher total nitrogen and total phosphorus concentrations (Nabout et al., 2006).
We recorded 577 species in 2000 and 2001. In 2000, species richness was higher in the low-water period than the high-water period (Fig. 1a) in most lakes; this pattern was similar in 2001 (Fig. 1b). Chlorophyceae had the highest number of species in all hydrologic periods, followed by Cyanophyceae in the 2000 rainy and dry seasons, and Zygnemaphyceae in the 2001 rainy and dry seasons.

Number of species for lakes (α diversity). In (a) the lakes in the rainy and dry season of 2000 and in (b) lakes in the rainy and dry seasons of 2001. The letters indicate the classifications of the lakes: A—abandoned channel; B—oxbow chains; C—meander scroll and D—blocked valley. Lake 19 was not classified
There was no single taxon present in all lakes surveyed. However, species such as Chroococcus minor (Kütz) Näg., Cryptomonas erosa Ehr., C. marsonii Skuja, C. obovata Skuja, Trachelomonas volvocinopsis Sw. and Scenedesmus ellipticus Corda were the most frequent.
The linear regression analysis demonstrated that the species richness of each lake (alpha diversity) was significantly related to latitude in the rainy season of 2000 (r = −0.40, P = 0.08 and n = 20), rainy season of 2001 (r = −0.77, P = 0.0001 and n = 19) and dry season of 2001 (r = −0.36, P = 0.1 and n = 20), although not in the dry season of 2000 (r = −0.18, P = 0.45 and n = 20). Species richness increased towards lower latitudes. Alpha diversity seemed to increase downstream (in this case from south to north). The determinants of species richness are the environmental variables correlating with latitude and not latitude itself. Latitudinal gradients of algal biodiversity in fluvial environments have received little attention therefore no comparable data exist. Freshwater fish and macroinvertebrates show greater diversity in the Tropics (Rohde, 1998).
Total species richness (gamma diversity) was high in all seasons, exceeding that of other floodplain lakes; the 2000 dry season had the highest (315 taxa) and the 2001 dry season the lowest (197 taxa; Table 2). Values obtained from the estimator indicated relatively thorough sampling in all seasons. In the Curuaí várzea (Nogueira, I.S.: unpublished data from the Curuaí floodplain in the Amazon Basin) using the same richness estimator as in this work, 60% and 62% (for 2002 and 2003 respectively) of the phytoplankton community was inventoried. Thus species richness found in the Middle Araguaia River várzea was similar to the Curuaí floodplain.
According to β-1 values, species composition in 2000 was more dissimilar in the high-water period than the low-water period; in 2001 it was similar in both periods (Table 2). This may have been because the flood pulse was less intense in 2000 (less rain; Nabout et al., 2006). The effect of environmental variability is measured by the sum of the coefficient of variation of limnological variables (total nitrogen, pH, conductivity, and transparency). Pearson correlation (P < 0.05) between the sum of coefficient of variation (environmental variables) and β-1 was significantly positive (r = 0.93; P = 0.05 and n = 4). According to Harrinson et al. (1982), this can be generally expected and has been found in other studies using different taxonomic groups.
Beta diversity indicated that phytoplankton composition was more dissimilar in the 2000 rainy season, whereas in 2001 levels in both seasons were equally dissimilar. According to Thomaz et al. (1997), flood pulses have a homogenizing effect, increasing similarity between lakes. However, this was not occurred for the phytoplankton community in the Middle Araguaia lakes, because rainy season beta diversity was higher than in the dry season. This might have occurred because the sampling area is extensive and is located in the Cerrado (Brazilian savanna). This area is subject to isolated and intense rains (precipitation over 200 mm). Isolated heavy rainfall events over such a large area lead to increased environmental heterogeneity which elevates beta diversity. Pearson correlation analysis demonstrated that β-1 was positively correlated with monthly precipitation level (r = 0.72, P = 0.02 and n = 4).
The 21 Middle Araguaia River lakes are distributed along a wide latitudinal gradient (240 km) and they vary in morphology, geology, limnology, and degree of human impact. These diverse characteristics lead to high species richness and high beta diversity. Pinheiro (2004) evaluating the geomorphological characteristics of these lakes found that they have different origins and consequently have different degrees of connectivity to the river: the abandoned channel lakes have the highest degree of connectivity, and the blocked valley lakes the lowest. According to Bini et al. (2003), connectivity has important implications for biodiversity dynamics because it influences the interchange of energy and matter (organisms) between the river and floodplain lakes. ANOVA indicated that in 2000, species richness was not significantly different in the rainy and dry seasons (rainy, F = 0.701 and P = 0.51; dry, F = 1.88 and P = 0.18) for the three different types of Middle Araguaia River lakes (abandoned channel or oxbow, oxbow chain, and blocked valley), while in 2001 only in the rainy season did the lakes show significant differences (rainy, F = 5.16 and P = 0.018; dry, F = 0.66 and P = 0.52). This lack of species richness difference between lake types in most seasons was probably due to a latitudinal pattern; in other words, the lakes at lower latitudes had higher species richness independent of their geomorphologic characteristics.
Phytoplankton composition in these different types of lake (oxbow, oxbow chain, meander scroll and blocked valley) also did not show substantial difference: their DCA scores did not separate in the graph (Fig. 2). DCA (the first two axes explained 34.1% of total data variability) also indicated that the phytoplankton composition differed between the rainy and dry seasons in both years, indicating a temporary difference and suggesting that flood pulse influences phytoplankton composition. Flood pulses are an important regulator of phytoplankton community changes (Thomaz et al., 1997; Train & Rodrigues, 2004; Melo & Huszar, 2000; Oliveira & Calheiros, 2000; Nabout et al., 2006). In addition to differences between the rainy and dry seasons, DCA also indicated a difference in phytoplankton composition between 2000 and 2001, suggesting that interannual variability influenced phytoplankton composition. Nabout et al. (2006), studying the same lakes, also found limnological differences between 2000 and 2001.

DCA scores for presence/absence data of species in all seasons. The lakes are indicated by points
The spatial distribution patterns of the phytoplankton community were assessed by the Mantel test applied to the sampling periods. The Mantel test indicated a positive correlation between geographic distance and floristic dissimilarity (Index 1-Jaccard). In the 2000 rainy season r = 0.24 and P = 0.0027, and dry season r = 0.14 and P = 0.037; in the 2001 rainy season r = 0.41 and P = 0.000001, and dry season r = 0.098 and P = 0.11, which was not significant. This suggests that geographically proximate lakes had similar floristic compositions, even with different geomorphological characteristics and degrees of human impact. According to Legendre & Fortin (1989), spatially proximate individuals are influenced by the same local processes.
Amoros & Bornette (2002) observed that floodplain systems are characterized by high alpha, beta, and gamma diversity, which result from the combination of complex habitat condition and hydrological connectivity gradients. Recently, Agostinho et al. (2005) noted that conservation of definite stretches of main rivers and their floodplains together with maintaining the region’s hydrological integrity are fundamental in preserving the biodiversity of Brazilian continental waters and the consequent maintenance of aquatic resources. In this context, and based on the information obtained from phytoplankton, the Middle Araguaia River floodplain should be given conservation priority.
References
Agostinho A. A., S. M. Thomaz, & L. C. Gomes, 2005. Conservation of the Biodiversity of Brazil’s Inland Waters. Conservation Biology 19: 646–652.
Amoros C. & G. Bornette, 2002. Connectivity and biocomplexity in waterbodies of riverine floodplains. Freshwater Biology 47: 761–776.
Bicudo C. E. M. & M. Menezes, 2005. Gêneros de Algas de Águas Continentais do Brasil – Chave para Identificação e Descrições [Algae Continental water in Brazil – identification Key and Descriptions]. Rima, São Paulo.
Bini L. M., L. F. M. Velho & F. A. Lansac-Tôha, 2003. The effect of connectivity on the relationship between local and regional species richness of testate amoebae (Protozoa, Rhizopoda) in floodplain lagoons of the upper Paraná River, Brazil. Acta Oecologica 24:145–151.
Burnham K. P. & W. S. Overton, 1978. Estimation of the size of a closed population when capture probabilities vary among animals. Biometrika 65: 625–633.
Chazdon R. L., R. K. Colwell, J. S. Denslow & M. R. Guariguata, 1998. Statistical methods for estimating species richness of woody regeneration in primary and secondary rain forests of Northeastern Costa Rica. In Dallmeier F. & J. A. Comiskey (eds). Forest biodiversity research. monitoring and modeling. The Parthenon Publishing Group. Washington, 285–309.
Colwell. R. K., 1997. Estimates 5: Statistical estimation of species richness and shared species from Samples. User Guide 22 p.Web site: www.viceroy.eeb.uconn.edu/estimates.
Gaston K. J., 1996. Biodiversity. A biology of number of difference. Blackwell, Oxford.
Harrinson S., S. J. Ross, J. H. Lawton, 1982. Beta diversity on geographic gradients in Britain. Journal Animal Ecology 62: 151–158.
Hill. M. O & H. G. Gauch, 1980. Detrend correspondence analysis, an improved ordination technique. Vegetatio 42: 47–58.
Junk W. J., P. B. Bailey & R. E. Sparks, 1989. The flood pulse concept in river-floodplain systems. In Dodge D. P. (ed), Proceedings of the International Large River Symposium. Canadian Special Publication of Fisheries and Aquatic Science 106: 110–127.
Legendre P. & M. J. Fortin, 1989. Spatial pattern and ecological analysis. Vegetatio 80: 107–138.
Mantel N., 1967. The detection of disease clustering and a generalized regression approach. Cancer Research 27: 209–220.
McCune B. & M. J Mefford, 1997. PC-ORD for Windows. Multivariate Analysis of Ecological Data Version 3.0. MjM Sotware. Gleneden Beach, Oregon. USA.
Melo S. & V. L. M. Huszar, 2000. Phytoplankton in Amazonian flood-plain lake (Lago Batata. Brasil): Diel variation and species strategies. Journal of Plankton Research 22: 63–76.
Nabout J. C., I. S. Nogueira & L. G. Oliveira, 2006. Phytoplankton community of floodplain lakes of the Araguaia River, Brazil, in the rainy and dry seasons. Journal of Plankton Research 28: 181–193.
Neiff J. J., 1996. Large rivers of South America: toward the new approach. Verhandlungen der internationalen Vereinigung für theorische und angewandte Limnologie 26:167–180.
Oliveira M. D. & D. F. Calheiros, 2000. Flood pulse influence on phytoplankton communities of the south Pantanal floodplain, Brazil. Hydrobiologia 427: 101–112.
Pinheiro, R. C. D., 2004. Lagoas de Inundação do médio Rio Goiás/Mato Grosso – Características limnológicas. Araguaia [Floodplain lakes of the middle Araguaia River, Goiás/Mato Grosso – Limnological characteristics]. Goiânia: Instituto de Ciências Biológicas/Universidade Federal de Goiás 107 p.Master Dissertation. [Dissertação de Mestrado].
Rohde K., 1998. Latitudinal gradients in species diversity: area matters, but how much? Oikos 82: 184–190.
Sokal R. R. & F. J. Rohlf, 1981. Biometry. 2nd. Ed. W.H. Freeman and Company, New York.
Thomaz S. M., M. C. Roberto & L. M. Bini, 1997. Caracterização limnológica dos ambientes aquáticos e influência dos níveis fluviométricos. [Limnological characterization of aquatic environments and the effect of water level]. In Vazzoler A. E. A. M., A. A. Agostinho & N. S. Hahn (eds) A Planície de Inundação do Alto Rio Paraná. Aspectos físicos, biológicos e socioeconômicos, EDUEM UEM-NUPELIA, 73–102.
Train S. & L. C. Rodrigues, 2004. Phytoplanktonic assemblages. In S. M. Thomaz A. A. Agostinho & N. S. Hahn (eds) The Upper Paraná River and its floodplain: physical aspects, ecology and conservation. Backhuys. Leiden, 103–124.
Van den Hoeck C., D. G. Mann & H. M. Jahns, 1993. Algae: an introduction to phycology. Cambridge University Press, Cambridge.
Vollenweider R.A., 1974 A Manual on Methods for Measuring Primary Production in Aquatic Environments. IBP. N 12, 2nd. Blackwell Sci. Publ., Oxford.
Ward J. V., K. Tockner & F. Schiemer, 1999. Biodiversity of floodplain river ecosystems: ecotones and connectivity. Regulated River: Research and Management 11: 105–119.
Acknowledgements
We are thankful to SECTEC for the financial support of the project, to the Environmental Agency of Goiás State for the aid in the field work, and to the NUPELIA-UEM for processing of the physical-chemical analysis. We also thank Dr. Luis M. Bini (Universidade Federal de Goiás) and Dra. Vera L. M. Huszar (Museu Nacional/ UFRJ) for critical comments on our manuscript and Dra. Judit Padisák and anonymous reviewer for corrections and suggestions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: J. Padisak
Rights and permissions
About this article
Cite this article
Nabout, J.C., de Nogueira, I.S., de Oliveira, L.G. et al. Phytoplankton diversity (alpha, beta, and gamma) from the Araguaia River tropical floodplain lakes (central Brazil). Hydrobiologia 575, 455–461 (2007). https://doi.org/10.1007/s10750-006-0393-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-006-0393-8