
Abstract
Negative gravitropic bending and its possible mediator in etiolated Alaska pea seedlings were intensively studied in comparison with seedlings of an agravitropic mutant, ageotropum. When 3.5-day-old etiolated Alaska seedlings were horizontally placed, the growth suppression at the upper side of the epicotyls began 10 min after the onset of the gravitropic stimulation, whereas the growth acceleration at the lower side began at 30 min, resulting in negative gravitropic bending. In contrast, no gravitropic bending was observed in the etiolated ageotropum seedlings, for which the epicotyls show an automorphogenesis-like growth. Strenuous efforts to identify a possible mediator that induces the gravitropic bending resulted in successfully identifying β-(isoxazolin-5-on-2yl)-alanine (βIA). The unilateral application of βIA to the etiolated Alaska epicotyls substantially induced epicotyl bending toward the application site, indicating that βIA could act as a growth inhibitor. Analyses of the distribution of βIA in the upper and lower flanks of the etiolated Alaska epicotyls revealed that its content rapidly increased twice in the upper flanks compared with that in the lower ones in response to gravitropic stimulation, whereas its content in the lower flanks was almost equal to that in the vertical control. In etiolated ageotropum epicotyls, an almost equal amount of βIA was distributed in the upper and lower flanks of epicotyls. These results suggest that a gravitropic stimulation increases βIA in the upper flank, resulting in the negative gravitropic bending of epicotyls via the suppression of the growth rate at the upper side of epicotyls in the etiolated Alaska pea seedlings.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Gravitropism, the developing curvature of a growing organ in response to some asymmetric external gravity, is easily observed, and thus, is open to obvious experiments. Not surprisingly, gravitropism has been a subject of investigation since the earliest times of plant physiology, as has as phototropism. The mechanism of phototropism can be explained in terms of perception of a stimulus vector, signal transduction, and the lateral distribution of a plant growth substance. This growth-promoting substance distributed somehow asymmetrically, and asymmetric growth causing a curvature in the elongating tissue was observed. The study of phototropism led to the discovery of auxin. In gravitropism of coleoptiles that were horizontally placed, it has been generally believed that the negative (upward) gravitropic bending is caused by gravity perception in the tip, thus leading to auxin transport to the lower side, followed by basipetal auxin translocation to the elongation zone below the tip where the differential flank growth results in upward growth (Cholodny–Went theory; see Went and Thimann 1937).
However, the growth-regulating mechanism underlying the negative gravitropic bending in dicotyledonous shoots remains unclear. If the Cholodny–Went theory is correct, gravity may cause negative gravitropic bending in a horizontal shoot by increasing the rate of cell elongation at the lower side of the shoot, accompanied by a decreasing the rate of elongation of cells at its upper side (Firn et al. 2000). A redistribution of growth such that the lower side starts elongating while the upper side simultaneously suppressed following applied gravitropic stimulation has been observed in both etiolated sunflower hypocotyls (Firn et al. 1978) and etiolated cucumber hypocotyls (Cosgrove 1990), thus lending support to the main prediction of the Cholodny–Went theory. Digby and Firn (1979) also reported that the suppression of growth on the upper side of etiolated maize coleoptiles, de-etiolated sunflower hypocotyls, and etiolated cucumber hypocotyls is a major factor bringing about gravitropism. Considering these facts alongside the results of phototropism, Bruinsma and Hasegawa (1990) proposed a new theory of phototropism, it being the consequence of the regulation by a light-induced gradient of auxin-inhibiting substances (Bruinsma–Hasegawa theory). Tokiwa et al. (2006) also demonstrated that a major factor in gravitropism in radish hypocotyls is the suppression of growth at the upper side of the hypocotyls, and the possible involvement of 4-methylthio-3-butenyl isothiocyanate. The apparent discrepancies among these theories suggest the necessity to obtain a clearer picture of the changes in growth rate across a range of species before attempting to set up a detailed general model of how the process of gravitropism is controlled in plants.
In the present study, we report on detailed analyzes of the lengths of the upper and lower sides of etiolated Alaska and an agravitropic mutant, ageotropum, pea epicotyls just prior to and following their gravitropic stimulation (studied with an infrared-imaging system). In addition, the successful identification of β-(isoxazolin-5-on-2yl)-alanine (βIA, Fig. 1) as a possible mediator of gravitropism increasing quantitatively in response to the gravitropic stimulation in etiolated Alaska seedlings is also described. Possible mechanisms of gravitropism as mediated by the compound βIA are also briefly discussed.

β-(Isoxazolin-5-on-2yl)-alanine, a gravitropism-regulating substance, in the etiolated pea epicotyls
Materials and methods
Plant materials
Two kinds of pea plants (Pisum sativum L.), cv. Alaska and an agravitropic mutant, ageotropum, were used in our study. The Alaska seeds were purchased from Watanabe Seeds Co., Ltd. (Miyagi, Japan), and the ageotropum seeds were propagated in the experimental field of the laboratory at Osaka Prefecture University (these seeds were kindly supplied by Prof. Hideyuki Takahashi, Tohoku University, Japan). The ageotropum mutant, an X-ray radiation-induced mutant of the pea cv. Weibull’s Witor (Blixt et al. 1958), is known to display agravitropic behavior in both its shoots and roots (Schöldéen and Burström 1960; Olsen and Iversen 1980a, b; Takahashi and Suge 1991; Takahashi et al. 1991), and to show growth that is similar to the automorphogenesis observed in space and under simulated weightlessness conditions in a 3-dimensional clinostat (Ueda et al. 1999, 2014; Miyamoto et al. 2011, 2014).
Plant culture
The pea seeds were imbibed at 23 °C for 8 h in running tap water, then sown in moistened vermiculite in a plastic tray, where they were allowed to germinate and grow at 23 °C for 3.5 days in the dark. To measure the growth rates at the upper and the lower sides, and of the right and the left sides of the epicotyl in seedlings kept horizontally and vertically, respectively, ion exchange resin beads (Amberlite XAD-2) with lanolin were positioned on the proximal and distal sides to the cotyledons ca. 2 and 12 mm below the hook of 12 etiolated seedlings. These seedlings were kept in a vertical position for 30 min to determine the growth rate prior to the gravitropic stimulation. They were then placed horizontally for 100 min for gravitropic stimulation, or left vertically as the control.
The epicotyl bending and the distance between the beads at the both sides were measured at 5-min intervals using an infrared (IR)-imaging system. This system consisted of two separated stations: one is a recording station that used an IR-sensitive MiniDV Handycam (DCR-TRV9 NTSC, Sony, Japan), and the other is a monitoring station that included a digital video application (DVBK-W2000, Sony, Japan) and Photoshop 5.5 software (Adobe Systems Incorp., USA). Images of the seedlings were recorded throughout the experiment which was monitored in real time. These manipulations were performed under dim green light (0.03 μmol m−2 s−1). The experiments were repeated at least three times.
Isolation and identification of a possible mediator of gravitropism
Elongating epicotyl regions [95 g fresh weight (FW)] of the 3.5-day-old etiolated Alaska seedlings placed vertically were harvested and immediately frozen in liquid N2, and then stored at −40 °C until extraction. The plant materials were homogenized in a blender and extracted with 1 L of 80% cold aqueous acetone. After filtration, the residues were re-extracted with 100 mL cold acetone. All the extracts were combined and concentrated to dryness at 40 °C in vacuo. The residue (4.72 g) was dissolved in MeOH and applied to a C18 Sep-Pak cartridge column (Waters, USA), and eluted with 0, 10, 20, 30, 50, and 100% MeOH in water (30 mL/step). The 0% MeOH fraction, which contained the peak responsible for the gravitropic stimulation (see Supplementary Fig. 2), was evaporated to dryness at 40 °C in vacuo. The residue (2.78 g) was dissolved in MeOH and applied to a NH2 Sep-Pak cartridge column (Waters, USA), and eluted with 80, 70, 60, 50, and 0% MeOH in water (25 mL/step). The 60% MeOH fraction was evaporated to dryness at 40 °C in vacuo, and then finally purified by a reversed-phase HPLC [column: ODS-120A, 7.8 × 300 mm, Tosoh, Japan; elution: linear gradient from 0 to 100% CH3CN in water for 20 min; flow rate: 2 mL/min, detector: PD-8020 multi-wavelength detector (Tosoh, Japan) absorbance at 195, 205, and 280 nm]. Eluate of the peak (retention time 5.5–7.5 min) corresponding to the quantitative increase in the epicotyls in response to the gravitropic stimulation in Alaska were obtained, and evaporated to dryness at 40 °C in vacuo, which yielded 2 mg of substance.
The 1H-NMR spectra of the compound were measured on a Bruker AVANCE-500 spectrometer (500 MHz) in D2O + AcOD. The 2D NMR 1H–1H correlation spectroscopy, heteronuclear multiple-bond correlation experiment, and NOE correlation spectroscopy analyzes were carried out to identify the compound’s chemical structure. The ESI-MS was recorded on a Waters Synapt G2 mass spectrometer.
Quantification of β-(isoxazolin-5-on-2yl)-alanine
Etiolated Alaska and ageotropum seedlings grown for 3.5 days were divided into two groups: one was incubated vertically as the control, while the other group was subjected to gravitropic stimulation for 60 min. To survey the growth regulator(s) showing quantitative changes in the epicotyls of Alaska seedlings, but not in ageotropum ones, in response to the gravitropic stimulation, 10 mm of the elongating region of the epicotyls ca. 2 mm below the hook was harvested, immediately frozen in liquid N2, and then stored at −40 °C until use.
For the analysis of the endogenous levels of the growth inhibitor, βIA, a 10-mm epicotyl region 2 mm below the hook of the Alaska seedlings positioned vertically and horizontally was harvested; the epicotyl segments were divided into halves (i.e., upper and lower halves, with the longitudinal halves as the control, ca. 20 mg FW) with a razor blade and immediately frozen in liquid N2, and then stored at −40 °C until use.
The frozen samples were homogenized in 10 mL of 80% cold aqueous acetone using a mortar and pestle. The filtrated extracts were concentrated to dryness at 40 °C in vacuo. The samples were dissolved in MeOH and applied to a reversed-phase HPLC, as mentioned previously. To quantify βIA, the area under the peak was measured. This experiment was repeated five times.
Unilateral application of β-(isoxazolin-5-on-2yl)-alanine
Ten 3.5-day-old etiolated Alaska seedlings were treated unilaterally with the growth-regulating substance, βIA, administered as a lanolin paste. The treated seedlings were placed vertically in the dark at 23 °C. The extent of epicotyl bending in the direction of the site of unilateral application at 20, 40, and 60 min after the application was determined. All manipulations were carried out under dim green light as mentioned above.
Results
Kinetics of negative gravitropic bending in the etiolated Alaska and ageotropum pea epicotyls
The negative gravitropic bending and the growth rates at both the upper and the lower sides in the horizontally placed epicotyls of the etiolated Alaska and ageotropum pea seedlings were measured prior to, and following, the gravitropic stimulation.
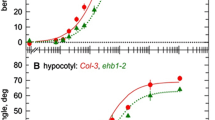
As shown in Fig. 2 and Supplementary Fig. 1, the epicotyls of the etiolated Alaska seedlings showed typical negative gravitropic bending when they were horizontally placed. The negative gravitropic bending in the etiolated Alaska epicotyls steadily increased from 10 min after the onset of gravitropic stimulation (as indicated by the arrow in Fig. 2a). The growth suppression at the upper side in the etiolated Alaska epicotyls began at least 10 min after the onset of gravitropic stimulation (as indicated by arrow a in Fig. 2c) and it continued during the period of gravitropic stimulation. Growth acceleration at the lower side was not observed within 30 min after the onset of gravitropic stimulation (as indicated by arrow b in Fig. 2c). These results indicated that the growth suppression at the upper side is important for generating the negative gravitropic bending in the etiolated Alaska epicotyls.

Time course of the gravitropic curvature (a, c) and the growth rate at the upper and lower sides (b, d) of etiolated epicotyls in the Alaska (a, b) and ageotropum (c, d) pea seedlings prior to, and following, the gravitropic stimulation. Etiolated pea seedlings were grown vertically for 3.5 days and then placed horizontally for gravitropic stimulation at time 0. The dotted line indicates the start time of the gravitropic stimulation
In contrast, the ageotropum epicotyls did not show any negative gravitropic bending, nor did they exhibit changes in growth rate at the upper and the lower sides in the horizontally positioned seedlings (Fig. 2b, d, Supplementary Fig. 1). The growth rate of the ageotropum epicotyls positioned vertically was similar to that of the Alaska ones.
Survey of possible compound(s) involved in the negative gravitropic bending in pea epicotyls
The results shown in Fig. 2 suggest the possible involvement of a plant growth regulator(s) which show quantitative changes in the upper flank of etiolated Alaska epicotyls, but not in the lower one, in response to the gravitropic stimulation. We also expected little quantitative differences in the plant growth regulator(s) between the ageotropum epicotyls positioned vertically and horizontally. To investigate this, we compared the HPLC chromatograms of the extracts from the horizontal- and vertical-positioned pea epicotyls of 3.5-day-old etiolated Alaska and ageotropum seedlings at 60 min after the start of the treatment. The peak A was higher in the horizontal-positioned Alaska epicotyls than in the vertical-positioned ones (Supplementary Fig. 2A, B), whereas the peak went unchanged in the ageotropum epicotyls (Supplementary Fig. 2C, D). This result suggested that the compound corresponding to the peak A may be considered a candidate for regulating the gravitropic response observed in the etiolated pea epicotyls.
Strenuous efforts to find the growth regulator showing a quantitative increase in the epicotyls in response to the gravitropic stimulation were successful. We were able to isolate the peak A compound from the elongating epicotyl region of the 3.5-day-old etiolated Alaska seedlings based on its retention time on HPLC. This compound was identified as βIA (Fig. 1) based on the spectra of the 1H NMR and 2D NMR analyzes, in complete agreement with the data in the reported literature (Lambein et al. 1992).
Inhibitory effect of β-(isoxazolin-5-on-2yl)-alanine on growth in the pea epicotyls
To evaluate the possible role in negative gravitropic bending played by βIA, its biological activity was examined. The unilateral application of βIA caused substantial epicotyl bending toward the direction of the application site in vertically placed etiolated Alaska epicotyls (Table 1), thus indicating that it can function as a growth inhibitor.
Quantitative changes in β-(isoxazolin-5-on-2yl)-alanine in the pea epicotyls in response to gravitropic stimulation
The content of βIA in the elongating region of the etiolated Alaska epicotyls increased almost twice when subjected to gravitropic stimulation for 60 min compared with the vertically placed control (Table 2). On the other hand, the content in etiolated ageotropum epicotyls was not affected by gravitropic stimulation. Since these suggest that βIA is responsible for gravitropic stimulation, detail changes in epicotyls were analyzed.
The content of βIA was increased almost twice in the upper flank, but not in the lower flank of the epicotyls within 30 min after the onset of gravitropic stimulation (Table 3). The endogenous level of the compound in the lower flank was almost equivalent with that in the right and the left haves of epicotyl in etiolated Alaska seedlings positioned vertically. The local gradient of the compound was continued until at least 60 min after the onset of gravitropic stimulation.
Discussion
Gravitropic bending of the epicotyl in pea seedlings
Curvatures due to growth are irreversible, resulting in a tropism. The growth-regulating mechanism underlying the negative gravitropic bending in dicotyledonous shoots has not been settled yet. As described in the “Introduction” section above, if the Cholodny–Went theory relating to auxin redistribution is correct, gravity should induce a negative gravitropic curvature in a horizontal shoot by a deceleration of elongation at the upper side in tandem with an acceleration at the lower side (Firn et al. 2000). The negative gravitropic bending of the epicotyl in the etiolated Alaska seedlings began 10 min after the onset of gravitropic stimulation, which then increased the negative gravitropic curvature (Fig. 2). The growth suppression at the upper side in the etiolated Alaska epicotyls started at least 10 min after the onset of gravitropic stimulation and was maintained for 100 min, whereas the growth acceleration at the lower side only began 30 min after the onset of the gravitropic stimulation (Fig. 2).
Altogether, these results suggest that the growth suppression at the upper side is first induced by gravity, while the growth acceleration at the lower side is the result rather than a cause of the negative gravitropic bending in the gravitropically stimulated etiolated pea epicotyls. It is therefore possible that a redistribution of auxin played a role in the maintenance of gravitropic upward curvature, but not as an inducer of the negative gravitropic response, even if an asymmetric redistribution of auxin occurred in response to the applied gravitropic stimulation.
The pattern of growth in the etiolated Alaska epicotyls was almost similar to that reported for radish hypocotyls during gravitropic stimulation by Tokiwa et al. (2006). The growth pattern in response to the unilateral gravity vector in both etiolated Alaska epicotyls and radish hypocotyls is also similar to that obtained from the phototropic studies of many plant species in which the measure of phototropic bending depended on the magnitude of growth suppression at the illuminated side and an unchanged growth rate at the shaded side (Franssen et al. 1981; Hasegawa et al. 1987, 2004a, b; Hasegawa and Yamada 1992; Yokotani-Tomita et al. 1999). From the abundant physiological and chemical studies of phototropism, the postulated hypothesis is that phototropism is caused by a local gradient of light-induced growth inhibitor(s) at the illuminated side set against a background of an unchanged, even distribution of auxin (so-called Bruinsma–Hasegawa theory, see the Introduction, Bruinsma and Hasegawa 1990; Yamamura and Hasegawa 2001). A lot of papers supporting this mechanistic view have already been published. For example, 8-Epixanthatin (Yokotani-Tomita et al. 1999) and 8-(β-d-glucopyranosyloxy)-3-hydroxy-1,9,14-pentadecatriene-4,6-diyne designated as helian (Hasegawa et al. 2007) from sunflower hypocotyls; benzoxazinoids, such as 2,4-dihydroxy-7-methoxy-1,4-benzoxazin-3-one (DMBOA) and 6-methoxybenzoxazolinone from maize coleoptiles (Jabeen et al. 2007); raphanusanin and its precursor 4-methylthio-3-butenyl isothiocyanate and 4-methylthio-3-butenyl glucosinolate from radish hypocotyls (Hasegawa et al. 2000); indolyleacetnitrile from cabbage and Arabidopsis shoots (Kosemura et al. 1997; Hasegawa et al. 2004a); and uridine from Avena coleoptiles (Tamimi 2004) have all been demonstrated as important in phototropism by functioning as a light-induced growth inhibitor. Therefore, it is not unreasonable to adapt the Bruinsma–Hasegawa theory to gravitropism in the etiolated pea epicotyls of the present study.
β-(Isoxazolin-5-on-2yl)-alanine acts as a negative gravitropic-bending regulating substance
As described above, a gravitropic stimulation-induced growth inhibitor is possibly involved in gravitropism. Much effort led to the successful identification of βIA in our study. βIA from pea seedlings has been described as the first member of a unique group of heterocyclic amino acids that are highly labile toward UV light and alkalinity (Lambein et al. 1969, 1976), and it is derived from O-acetyl-l-serine and isoxazolin via cysteine synthase in higher plants (Ikegami et al. 1991). The compound has also been found to be an inhibitory allelopathic substance in the roots and root exudates of pea seedlings while also showing antimycotic activity (Schenk and Werner 1991), and it is the metabolic precursor for the neurotoxin β-N-oxalyl-l-α,β-diaminopropionic acid (ODPA) (Lambein et al. 1990).
Unilateral application of βIA to the etiolated Alaska epicotyls that were positioned vertically caused their epicotyls to bend toward the site of application (Table 1), indicating it acts as an inhibitor. The analysis of the endogenous distribution of the compound between the upper and lower flanks of the epicotyl revealed that its content rapidly increased twice in the upper flank of etiolated Alaska epicotyls than that in the lower one after the onset of gravitropic stimulation, whereas in the etiolated ageotropum epicotyls there was no evident increase of the inhibitor in response to the gravitropic stimulation like for the agravitropic response (Tables 2, 3). This suggests that the gravitropic stimulation suppressed the growth rate at the upper side of pea epicotyls by increasing the βIA content, resulting in the negative gravitropic bending. A role in gravitropism has been demonstrated for jasmonates in rice coleoptiles (Gutjahr et al. 2005) and likewise for 4-methylthio-3-butenyl isothiocyanate and 3,6-disinapoylsucrose in etiolated radish hypocotyls (Tokiwa et al. 2006; Hasegawa et al. 2010). When these are taken together with our present results, an important role of a growth-inhibiting substance(s) for gravitropism in shoots is strongly suggested.
Edelmann (2001) has proposed a model for the principal mechanism regulating gravitropism, in which the gravitropic stimulation temporarily inhibits the infiltration of auxin-induced secreted wall-loosening factors into the outer cell wall of the epidermal cells, resulting in the inhibition of growth at the upper organ flanks in rye coleoptiles. Clark et al. (2000) have reported that close relationships between the gravity vector and the distribution of annexins, a known family of Ca2+-dependent membrane proteins that participate in a wide array of essential cellular processes including secretion, transport, and DNA replication. Hence, it is possible that gravitropic stimulation results in a redistribution of the annexin-mediated secretion or transport, thus modifying the redistribution of growth-regulating substance(s) or factor(s) in gravi-responsive-elongating cells.
In conclusion, our detailed analysis of growth rates at the upper vs. lower sides in the gravitropic-stimulated etiolated epicotyls of Alaska seedlings revealed an important role for growth suppression at the upper side in negative gravitropic bending. Great efforts to find this growth regulator were rewarded by the identification of βIA as a functional growth inhibitor that is synthesized at the gravitropically-stimulated upper flank of the etiolated pea epicotyls. Further analyses of an agravitropic pea mutant, ageotropum, supported the possible involvement of growth-inhibiting substances in gravitropism.
References
Blixt S, Ehrenberg L, Gelin O (1958) Quantitative studies of induced mutations in peas. I. Methodological investigation. Agric Hortic Genet 16:238–250
Bruinsma J, Hasegawa K (1990) A new theory of phototropism—its regulation by a light-induced gradient of auxin-inhibiting substances. Physiol Plant 79:700–704
Clark GB, Rafati DS, Bolton RJ, Dauwalder M, Roux SJ (2000) Redistribution of annexin in gravistimulated pea plumules. Plant Physiol Biochem 38:937–947
Cosgrove DJ (1990) Rapid, bilateral changes in growth rate and curvature during gravitropism of cucumber hypocotyls: implications for mechanism of growth control. Plant Cell Environ 13:227–234
Digby J, Firn RD (1979) An analysis of the changes in growth rate occurring during the initial stages of geocurvature in shoots. Plant Cell Environ 2:145–148
Edelmann HG (2001) Lateral redistribution of auxin is not the means for gravitropic differential growth of coleoptiles: a new model. Physiol Plant 112:119–126
Firn RD, Digby J, Riley H (1978) Shoot geotropic curvature—the location, magnitude and kinetics of the gravity-induced differential growth in horizontal sunflower hypocotyls. Ann Bot 42:465–468
Firn RD, Wagstaff C, Digby J (2000) The use of mutants to probe models of gravitropism. J Exp Bot 51:1323–1340
Franssen JM, Coole SA, Digby J, Firn RD (1981) Measurements of differential growth causing phototropic curvature of coleoptiles and hypocotyls. Z Pflanzenphysiol 103:207–216
Gutjahr C, Riemann M, Müller A, Düchting P, Weiler EW, Nick P (2005) Cholodny-Went theory revisited: a role for jasmonat in gravitropism of rice coleoptiles. Planta 222:575–585
Hasegawa K, Yamada K (1992) Even distribution of endogenous indole-3-acetic acid in phototropism of pea epicotyls. J Plant Physiol 139:455–459
Hasegawa K, Noguchi H, Tanoue C, Sando S, Takada M, Sakoda M (1987) Phototropism in hypocotyls of radish IV. Frank growth and lateral distribution of cis- and trans-raphanusanins in the first positive phototropic curvature. Plant Physiol 85:379–382
Hasegawa T, Yamada K, Kosemura S, Yamamura S, Hasegawa K (2000) Phototropic stimulation induces the conversion of glucosinolate to phototropism-relating substances of radish hypocotyls. Phytochemistry 51:275–279
Hasegawa T, Yamada K, Shigemori H, Goto N, Miyamoto K, Ueda J, Hasegawa K (2004a) Isolation and identification of blue light-induced growth inhibitor from light-grown Arabidopsis shoots. Plant Growth Regul 44:81–86
Hasegawa T, Yamada K, Shigemori H, Goto N, Miyamoto K, Ueda J et al (2004b) Isolation and identification of phototropism-relating substances benzoxazinoids from maize coleoptiles. Heterocycles 63:2707–2712
Hasegawa T, Togo S, Hisamatsu Y, Yamada K, Suenaga K, Sekiguchi M et al (2007) Isolation and structure elucidation of a potent growth inhibitor, helian, from blue light-illuminated sunflower (Helianthus annuus) hypocotyls. Heterocycles 71:609–617
Hasegawa T, Wai Wai Thet Tin, Shigemori H, Otomatsu T, Hirose K, Miyamoto K, Ueda J, Hasegawa K (2010) Isolation and identification of a gravity-induced growth inhibitor in etiolated rasish hypocotyls. Heterocycles 81:2763–2770
Ikegami F, Itagaki S, Ishikawa T, Ongena G, Kuo YH, Lambin F, Murakoshi I (1991) Biosynthesis of beta-(isozazolin-5-on-2-yl)alanine, the precursor of the neurotoxic amino acid beta-N-oxalyl-l-alpha, beta-diaminopropionic acid. Chem Pharm Bull 39:3376–3377
Jabeen R, Yamada K, Hasegawa T, Minami E, Shigemori H, Hasegawa K (2007) Direct involvement of benzoxazinoids in the growth suppression induced by phototropic stimulation in maize coleoptiles. Heterocycles 71:523–529
Kosemura S, Niwa H, Emori H, Yokotani-Tomita K, Hasegawa K, Yamamura S (1997) Light-induced auxin-inhibiting substances from cabbage (Brassica oleacea L.) shoots. Tetrahedron Lett 38:8327–8330
Lambein F, Schamp N, Vandendriessche L, Van Parijs R (1969) A new UV-sensitive heterocyclic amino acid from pea seedlings: 2-alanyl-3-isoxazolin-5-ones. Biochem Biophys Res Commun 37:375–382
Lambein F, Kuo YH, Van Parijs R (1976) Isaoxazolin-5-ones, chemistry and biology of a new class of plant products. Heterocycles 4:567–593
Lambein F, Ongena G, KuoY-H (1990) β-Isoxazoline-alanine is involved in the biosynthesis of the neurotoxin β-N-oxalyl-L-α,β-diaminopropionic acid. Phytochemistry 29:3793–3796
Lambein F, Khan JK, Becua C, Bruyn AD (1992) Characterization of γ-glutamyl-β-(isoxazolin-5-on-2yl)-alanine from Lathyrus sativus and its decarboxylation product from Lathyrus odoratus. Phytochemistry 31:887–892
Miyamoto K, Uheda E, Oka M, Ueda J (2011) Auxin polar transport and automorphosis in plants. Biol Sci Space 25:57–68
Miyamoto K, Yamasaki T, Uheda E, Ueda J (2014) Analysis of apical hook formation in Alaska pea with a 3-D clinostat and gravitropic mutant ageotropum. Front Plant Sci 5:137. doi:10.3389/fpls.2014.00137
Olsen GM, Iversen T-H (1980a) Growth and curvature in seedlings of Pisum sativum and an ageotropic mutant. Physiol Plant 50:269–274
Olsen GM, Iversen TH (1980b) Ultrastructure and movements of cell structures in normal and an ageotropic mutant. Physiol Plant 50:275–284
Schenk SU, Werner D (1991) β-(Isoxazolin-5-on-2-yl)-alanine from Pisum: allelopathic properties and antimycotic bioassay. Phytochemistry 30:467–470
Schöldéen C, Burström H (1960) Physiological studies of an ageotropic pea mutant. Physiol Plant 13:831–838
Takahashi H, Suge H (1991) Root hydrotropism of an agravitropic pea mutant, ageotropum. Physiol Plant 82:24–31
Takahashi H, Suge H, Jaffe MJ (1991) Agravitropic growth and its relation to the formation of the plumular hook in etiolated shoots of the pea mutant, ageotropum. J Plant Physiol 138:216–222
Tamimi SM (2004) Uridine and the control of phototropism in oat (Avena sativa L.) coleoptiles. Plant Growth Regul 43:173–177
Tokiwa H, Hasegawa T, Yamada K, Shigemori H, Hasegawa K (2006) A major factor in gravitropism in radish hypocotyls is the suppression of growth on the upper side of hypocotyls. J Plant Physiol 163:1267–1272
Ueda J, Miyamoto K, Yuda T, Hoshino T, Fujii S, Mukai C et al (1999) Growth and development, and auxin polar transport in higher plants under microgravity conditions in space: BRIC-AUX on STS-95 space experiment. J Plant Res 112:487–492
Ueda J, Miyamoto K, Uheda E, Oka M, Higashibata A, Ishioka N (2014) Close relationships between polar auxin transport and gravitresponse in plants. Plant Biol 16:43–49
Went FW, Thimann KV (1937) Phytohormones. The MacMillan Company, New York
Yamamura S, Hasegawa K (2001) Chemistry and biology of phototropism-regulating substances in higher plants. Chem Rec 1:362–372
Yokotani-Tomita K, Kato J, Yamada K, Kosemura S, Yamamura S, Bruinsma J et al (1999) 8-Epixanthatin, a light-induced growth inhibitor, mediates the phototropic curvature in sunflower (Helianthus annus L.) hypocotyls. Physiol Plant 106:326–330
Acknowledgements
This study was partially supported by a Grant-in-Aid for Scientific Research (C) (Grant No. 24620008 to KM, and Grant No. 26506018 to JU) from the Japanese Society for the Promotion of Science. The authors would like to thank Enago (http://www.enago.jp) for the English language review.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Hasegawa, T., Omiya, Y., Koide, M. et al. A gravitropic stimulation-induced growth inhibitor, β-(isoxazolin-5-on-2yl)-alanine, is a possible mediator of negative gravitropic bending of epicotyls in etiolated Pisum sativum seedlings. Plant Growth Regul 82, 431–438 (2017). https://doi.org/10.1007/s10725-017-0269-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-017-0269-0