
Abstract
Cytokinins are plant hormones that regulate diverse processes during plant growth and development. Targeted manipulation of their metabolism or perception has already shown benefits in agriculture. Recently, we described substances that can affect plants’ endogenous cytokinin status: a cytokinin antagonist, PI-55, and an inhibitor of cytokinin degradation, INCYDE (2-chloro-6-(3-methoxyphenyl)aminopurine). The effects of these substances on seedling growth in the medicinal plant Bulbine natalensis Baker (Asphodelaceae) and the metal tolerant Rumex crispus L. (Polygonaceae) under abiotic stress caused by cadmium (Cd) were tested. Cd is known for its negative effects on plant growth and its toxicity to humans. Treatment with either PI-55 or INCYDE had positive effects on seedling shoot and root growth and the fresh weight of treated seedlings grown in the presence of Cd. Even a single application of either compound at sub-micromolar concentrations was sufficient to reverse the inhibitory effects of Cd. Our results demonstrate that modulating cytokinin status with inhibitors of cytokinin perception and/or degradation may be useful in protecting plants against the adverse effects of high Cd levels.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Heavy metals such as cadmium (Cd) are important abiotic stress factors that limit plant performance and yield. Cd is one of the most toxic non-essential elements, having high mobility and the ability to inhibit the main physiological processes in plants. It is phytotoxic and interacts with photosynthesis, respiration and nitrogen metabolism in plants. This causes poor growth and low biomass due to oxidative stress caused by the generation of free radicals and active oxygen species (Hendry et al. 1992; Sanitá di Toppi and Gabbrielli 1999).
Cytokinins are a class of plant hormones that regulate the cell cycle and diverse developmental and physiological processes, including responses to biotic and abiotic stress. Unlike abscisic acid (ABA), ethylene and jasmonic acid (JA), they are not fundamental plant stress hormones. Nevertheless, they affect stress processes in a number of ways (Ha et al. 2012). It has been suggested that in longer term responses to abiotic stress, hormones such as ABA and cytokinins may regulate the production, metabolism and distribution of metabolites essential for stress survival and recovery (Pospíšilová and Dodd 2005). Cytokinin treatment reduces the negative effects of water deficits on chlorophyll and carotenoid content, which are important protective factors against oxidative stress caused by heavy metal uptake, and on photosynthetic activity (Metwally et al. 1997). Moreover, the transcription of many stress-induced genes is stimulated by cytokinins (Hare and Van Staden 1997). In tobacco and maize, endogenous cytokinin levels fall during water stress, primarily due to the activity of cytokinin oxidase/dehydrogenase (CKX), a key enzyme in cytokinin degradation (Alvarez et al. 2008; Havlová et al. 2008). In maize, CKX is induced by ABA and abiotic stress, so there is a direct connection between cytokinin down-regulation and abiotic stress tolerance. Another example is the expression of SAG12-IPT in cassava leaves, which delays leaf senescence and increases drought resistence (Zhang et al. 2010). Conversely, Arabidopsis lines that have reduced cytokinin sensitivity due to mutations in the cytokinin receptors AHK2, AHK3, or AHK4 are more resistant to drought, osmotic, and cold stress (Jeon et al. 2010; Tran et al. 2007). This suggests that both increases in cytokinin levels and decreases in the intensity of cytokinin signalling could potentially be exploited to increase stress tolerance and promote acclimatization in plants.
We have recently described multiple new synthetic cytokinin derivatives with specific biological effects. Systematic testing led to the identification of two very potent inhibitors of AtCKX2: 2-chloro-6-(3-methoxyphenyl)aminopurine and 2-fluoro-6-(3-methoxyphenyl)aminopurine (Zatloukal et al. 2008). The first of these compounds was named INCYDE (inhibitor of cytokinin degradation). It modulates endogenous cytokinin levels in planta and is a suitable tool for in vivo studies of cytokinin action (Aremu et al. 2012). A second screen of the cytokinin derivative library resulted in the identification of PI-55, the first known molecular antagonist of cytokinin activity at the receptor level. PI-55 accelerated the germination of Arabidopsis seeds and promoted root growth and formation of lateral roots, thus phenocopying the known consequences of a reduced cytokinin status. This clearly demonstrated its ability to inhibit cytokinin perception in planta (Spíchal et al. 2009). Both INCYDE and PI-55 were used in this work to assess the effect of modulating cytokinin levels and responses, respectively, on Cd stress.
The effects of treatment with PI-55 and INCYDE on germination and seedling growth in two plant species (Bulbine natalensis and Rumex crispus) grown under abiotic stress conditions caused by Cd exposure were investigated. B. natalensis is extensively used in traditional medicine in South Africa; the widespread R. crispus is a metal-tolerant plant with a relatively high biomass that is also used as a medicinal plant (Zhuang et al. 2007).
Materials and methods
Chemicals
Dimethylsulfoxide (DMSO) and mercuric chloride were obtained from Sigma (St., Luis, MO). N 6-benzyladenine was obtained from OlChemIm (Olomouc, Czech Republic). 2-chloro-6-(3-methoxyphenyl)aminopurine (INCYDE) and 6-(2-hydroxy-3-methylbenzylamino)purine (PI-55) were synthesized in by the Laboratory of Growth Regulators, Palacký University & Institute of Experimental Botany AS CR (Olomouc, Czech Republic) as previously described (Zatloukal et al. 2008; Spíchal et al. 2009).
Plant material
Seeds of B. natalensis Baker and R. crispus L. were obtained from the Univerzity of KwaZulu-Natal Botanical Garden, Pietermaritzburg, South Africa. The seeds were stored in brown paper bags at 10 °C in the dark for 6 months before the experiments.
Determination of the optimal PI-55 and INCYDE concentration for germination and seedling establishment
All seeds were surface decontaminated with 0.1 % mercuric chloride for 2 min and rinsed thoroughly with distilled water prior to germination. The first experiment to determine the optimal concentrations of PI-55 and INCYDE for germination and seedling establishment was performed as described by Street et al. (2007). Disposable Petri dishes containing two Whatman No. 1 filter paper discs were used for germination. Three replicates of 15 seeds each were used. Test solutions of either PI-55 or INCYDE in distilled water at concentrations ranging from 0.001–10 μM were added at the start of the experiment. Additional control experiments were performed involving treatment with distilled water, 0.1 % DMSO solution and 10 μM N 6-benzyladenine (BA). The treated Petri dishes were placed in plant growth chambers at 25 ± 0.5 °C under 16:8 h light: dark conditions, with a photosynthetic photon flux density of 80.4 ± 3.5 μmol M−2 s−1. The Petri dishes were checked daily and distilled water was added periodically to maintain moisture levels. Germination (scored as the emergence of a radicle with a length of at least 2 mm) was recorded every day. The experiment was terminated 2 weeks after the first observed germination. The number of lateral roots produced by the Rumex seedlings and adventitious roots produced by the Bulbine seedlings were counted, and the lengths of the roots and shoots and the fresh masses of the seedlings were recorded.
Testing of plant response to PI-55 and INCYDE in the presence of Cd
The second experiment was done in Petri dishes as described above but in the presence of 5 mg L−1 Cd (CdNO3). Three replicates of 25 seeds each were used and samples of the two plant species were treated with 10 nM PI-55, 10 nM INCYDE, and 10 μM BA (B. natalensis); or 10 nM PI-55, 100 nM INCYDE, 10 μM BA (R. crispus). These concentrations were selected based on the results of the initial experiment to determine the optimal concentration of PI-55 and INCYDE for germination and seedling establishment for the two species under investigation. Each treatment was applied in two different ways: (1) compounds diluted in distilled water were added once at the start of the experiment, after which only water or water containing Cd (5 mg L−1) and DMSO (0.1 %) was added every second day to maintain an appropriate moisture level, or (2) the tested compounds were added at the indicated concentrations every second day during watering. Germination (2 mm radicle emergence) was recorded every second day. The experiment was terminated after 21 days when the seedlings were developed, and morphological parameters (root and shoot length, fresh seedling mass and number of lateral or adventitious roots) were measured for each seedling.
Statistical analyses
All arcsine-transformed germination and seedling growth data were analyzed using one-way analysis of variance, and Duncan’s Multiple Range test at the 5 % level of significance was performed to separate the means of the treatments. All analyses were performed using release 12 of the GenStat® statistics package.
Results
Effect of PI-55 and INCYDE on germination and seedling development
In the initial experiments, plants were treated with PI-55 and INCYDE at concentrations of 1 nM–10 μM to establish the optimal concentrations for germination and seedling development. Neither compound alone had any significant effect on the germination of B. natalensis and R. crispus (Table 1). Characterisation of seedling growth revealed that plant organs responded differently to the PI-55 and INCYDE treatments. In R. crispus, PI-55 had no effect on shoot and root growth at any of the tested concentrations (Fig. 1a–c). However, INCYDE caused dose-dependent inhibition of both root and shoot growth (Fig. 1d, e). As was observed with the cytokinin control (10 μM BA), INCYDE at 10 μM strongly reduced the lengths of the shoots and roots (Fig. 1d, e), and to lesser extent, overall seedling weight (Fig. 1f). In the case of B. natalensis, treatment with PI-55 at concentrations of up to 1 μM did not significantly affect seedling growth. However, growth was reduced by approximately 30 % at the 10 μM concentration (Fig. 2). Treatment with INCYDE had no effect on shoot length (Fig. 2d) but higher concentrations (10 μM) significantly inhibited root growth (Fig. 2e).

The effects of the cytokinin antagonist PI-55 (a-c) and the inhibitor of cytokinin degradation INCYDE (d-f) on Rumex crispus growth. Seeds were germinated in water containing 0.001–10 μM of one of the two compounds, and the effects of the treatment on the seedlings’ shoot length (a, d), root length (b, e), and overall weight (c, f) were estimated. Results for control (Ctr), N6-benzyladenine (BA) and solvent control (DMSO) treatments are also shown. Bars show mean values and SD (n = 45); standard error bars with different letter(s) denote results that differ significantly (p < 0.05)

The effects of the cytokinin antagonist PI-55 (a–c) and the inhibitor of cytokinin degradation INCYDE (d–f) on B. natalensis growth. Seeds were germinated in water containing 0.001–10 μM of one of the two compounds and the effects of the treatment on the seedlings’ shoot length (a, d), root length (b, e), and overall weight (c, f) were estimated. Results for control (Ctr), N6-benzyladenine (BA) and solvent control (DMSO) treatments are also shown. Bars show mean values and SD (n = 45); standard error bars with different letter(s) denote results that differ significantly (p < 0.05). The effect of treatment with 1 μM INCYDE on seedling fresh weight could not be determined due to contamination (indicated by an asterisk in f)
Effect of PI-55 and INCYDE on growth of B. natalensis and R. crispus in the presence of Cd
The effects of PI-55 and INCYDE on growth under Cd stress were investigated using concentrations that had no adverse effects on seedling development in the first set of experiments. For B. natalensis, both compounds were applied at concentrations of 10 nM, while R. crispus was treated with PI-55 at 10 nM and INCYDE at 100 nM. A Cd concentration of 5 mg L−1 was used to induce abiotic stress; this is a typical concentration found in heavily Cd-contaminated soil solutions (Alloway 1990).
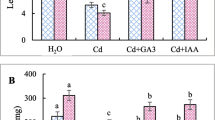
Cd treatment caused significant inhibition of R. crispus growth (Fig. 3), reducing seedling weight and shoot length by 30 % relative to the untreated control (Fig. 3a, d). Cd treatment also inhibited root growth by 40 % (Fig. 3c) and partially reduced the number of lateral roots (Fig. 3b). These negative effects on the root system were more pronounced in the presence of 10 μM BA (Fig. 3b, c). This is consistent with the known function of cytokinins as negative regulators of the root system (Cary et al. 1995; Skoog and Miller 1957). In contrast, BA had positive effects on shoot length and seedling weight in Cd-treated seedlings (Fig. 3a, d).

The effects of treatment with the cytokinin N6-benzyladenine (BA; 10 μM), the cytokinin antagonist PI-55 (10 nM) and the inhibitor of cytokinin degradation INCYDE (100 nM) on R. crispus growth in the presence of 5 mg L−1 Cd. The tested substances were either applied once at the begining of germination or continously at every watering (indicated with an asterisk) and the effects of each treatment on seedling weight (a), the number of lateral roots (b), root length (c), and shoot length (d) were determined. Control (Ctr) seedlings were treated with water containing 0.1 % DMSO. Bars show the mean values obtained in each case and the SD (n ≥ 23); standard error bars with different letter(s) denote results that differ significantly (p < 0.05)
The effect of PI-55 and INCYDE were tested in two ways: the compounds were either applied once at the start of the experiment or continuously throughout the whole experiment. Both compounds had positive effects on R. crispus seedlings grown in the presence of Cd (Fig. 3). PI-55 treatment overcame the negative effects of Cd, increasing the fresh weight of the seedlings, their root and shoot lengths (Fig. 3a, c, d), and the number of lateral roots produced (Fig. 3b). Even with only a single application at the beginning of the experiment, PI-55 protected root development in Cd-treated plants, doubling the number of lateral roots produced relative to the untreated control (Fig. 3b).
Treatment with INCYDE had positive effects on shoot length and seedling fresh weight similar to those observed with BA (Fig. 3a, d). However, the typical cytokinin-like negative effect on the root system was not observed even under long-term INCYDE treatment (Fig. 3b, c): the roots of seedlings treated with both INCYDE and Cd were almost as long as those of untreated control seedlings (Fig. 3c). Seedlings treated with INCYDE also produced around 30 % more lateral roots than controls treated with Cd alone (Fig. 3b).
In the monocotyledonous B. natalensis, the negative effect of Cd on plant growth was not as pronounced as in R. crispus (Fig. 4). Cd exposure reduced the seedlings’ fresh weight by more than 30 % compared to the control (Fig. 4a) but did not affect the number of roots produced at the investigated concentration. However, the root length was reduced by about 20 % compared to the control (Fig. 4b, c). Cd exposure also reduced the length of the shoots by around 25 % (Fig. 4d). Cytokinin (BA) treatment intensified the negative effects of Cd on plant performance, primarily by further reducing the length of the seedlings’ roots (Fig. 4c). Although treatment with PI-55 alone increased seedling fresh weight by about 15 %, it failed to overcome the negative effects of Cd (Fig. 4a). However, PI-55 treatment did provide significant protection against the retardation of root and shoot growth regardless of whether it was applied once or repeatedly (Fig. 4c, d). The effects of INCYDE were very similar to those of PI-55 (Fig. 4): although it did not overcome the effect of Cd on seedling fresh weight, it did protect the plants against Cd-induced retardation of shoot and root growth (Fig. 4c, d).

The effects of treatment with the cytokinin N6-benzyladenine (BA; 10 μM), the cytokinin antagonist PI-55 (10 nM), and the inhibitor of cytokinin degradation INCYDE (10 nM) on B. natalensis growth in the presence of 5 mg L−1 Cd. The tested substances were either applied once at the begining of germination or continously at every watering (indicated with an asterisk) and the effects of each treatment on seedling weight (a), number of roots (b), root length (c), and shoot length (d) were determined. Control (Ctr) seedlings were treated with water containing 0.1 % DMSO. Bars show the mean values obtained in each case and the SD (n ≥ 23); standard error bars with different letter(s) denote results that differ significantly (p < 0.05)
Discussion
Environmental pollution of soils and water by heavy metals has become a serious problem in recent times due to human activities. Low concentrations of heavy metals such as Fe, Cu, Zn or Ni are essential for some physiological processes in plants, but other metals such as Cd, Pb, Hg and Cr are phytotoxic (Clemens 2001). Heavily polluted areas may contain up to 100 μg g−1 Cd, giving soil concentrations of up to 32 μM (Alloway 1990). We performed experiments to determine whether modulating plants’ cytokinin status using a cytokinin antagonist (PI-55) and/or an inhibitor of cytokinin degradation (INCYDE) could promote germination and seedling establishment in the presence of Cd at concentrations that are common in polluted environments.
The rationale behind the use of the cytokinin antagonist to increase plant resistance against heavy metal stress was that decreased cytokinin perception has been shown to help plants in coping with osmotic, drought and salt stress (Tran et al. 2007). We have previously shown that the cytokinin antagonist PI-55 blocks the binding of natural cytokinins to their receptors, and that plants treated with this compound phenocopied cytokinin receptor mutants (Spíchal et al. 2009). As expected, PI-55 treatment stimulated root development in R. crispus and B. natalensis seedlings and remained effective in the presence of Cd (Figs. 3, 4). It is possible that PI-55 affected cytokinin perception in the treated seedlings and thereby conferred enhanced stress tolerance similar to that observed in plants with mutated cytokinin receptors. PI-55 also positively affected the growth of the root system and thus certainly improved nutrient uptake and utilization, which is otherwise limited in plants growing under Cd stress due to the retardation of root system growth. Even a single application of PI-55 at the start of germination was sufficient to protect root growth in R. crispus against the detrimental effects of Cd exposure.
The mechanism of PI-55 action is complicated by the fact that PI-55 is only a partial receptor antagonist. Although it blocks AHK4, it weakly activates AHK3 (Spíchal et al. 2009). Weak cytokinin activity may also have important protective effects. Exogenous cytokinin treatment reportedly has positive effects on the growth of Cd-stressed plants (Wozny et al. 1995; Lukatkin et al. 2003, 2007; Ul’yanenko et al. 2004). This is partially consistent with the results obtained in this work, which demonstrated that BA has protective effects on seedling shoot length and overall biomass formation in R. crispus (Fig. 3a, d) and to a lesser extent in B. natalensis (Fig. 4a, d). On the other hand, cytokinins negatively regulate root growth (Werner et al. 2001); in keeping with this function, we observed that Cd exposure in conjunction with BA treatment had an additive adverse effect on root system development. Cd is generally toxic to plants because it induces oxidative stress (Briat and Lebrun 1999), but treatment with exogenous cytokinin-derived compounds can protect against the oxidative damage caused by exposure to transition metals such as Cd (Lukatkin et al. 2007). The apparent difference in the effect of BA on B. natalensis and R. crispus may be due to the higher resistance of B. natalensis to Cd. Whereas only a single application of Cd inhibited root growth of R. crispus (Fig. 3c), continuous Cd application was needed to inhibit root growth in B. natalensis (Fig. 4c). In addition, B. natelensis forms adventitious roots that appear later during root system development and thus this morphological trait may not have been affected by Cd applied under the experimental conditions. Previous works have shown that treatment with the synthetic cytokinin kinetin or the CKX inhibitor thidiazuron (Chatfield and Armstrong 1986) can mitigate the negative effects of high concentrations of heavy metals such as Zn and Ni by increasing the activity of ascorbate peroxidase, one of the key antioxidant defence enzymes (Lukatkin et al. 2007). This finding explains the protective effect of INCYDE, an inhibitor of CKX that blocks cytokinin degradation in vivo (Gemrotová et al. 2009). Moreover, we recently showed that INCYDE treatment can significantly increase antioxidant capacity in plants (Gruz and Spíchal 2011). Overall, our data indicate that both approaches to modulating cytokinin status (i.e. decreasing it by blocking cytokinin signalling, or causing finely tuned increases in endogenous cytokinin levels by inhibiting their degradation) can protect against the adverse effects of Cd exposure. It was clearly shown that these protective effects are cytokinin-dependent. However, they are also likely to be influenced by a complex mixture of primary and secondary factors.
Rumex crispus is a plant that is known to tolerate high levels of heavy metals (Zhuang et al. 2007). Together with hyperaccumulators such as Viola baoshanensis, Thlaspi caerulescence and Sedum alfredii, it is suitable for phytoextraction (Zhuang et al. 2005). While its tolerance is lower than that of other hyperaccumulators, it accumulates high levels of Cd (Zhuang et al. 2007), has a relatively high biomass and can be grown worldwide. Recent studies performed under field conditions indicated that due to its high biomass, R. crispus can extract 26.8 kg ha−1 Zn and 0.16 kg ha−1 Cd from metal-contaminated soil with high efficiency (Zhuang et al. 2007). The phytoextraction potential of R. crispus is thus high and could be further improved by treatment with PI-55 or INCYDE at sub-micromolar concentrations (10–100 nM) to protect the roots against Cd stress. This may even increase the number of roots produced relative to untreated plants (Fig. 3). Moreover, even a single application of PI-55 or INCYDE at an early stage of development was sufficient to induce strong positive effects, further enhancing the practicality of these compounds. Overall, the results suggest that phytoextraction facilitated by these compounds could be useful in plant-based environmental remediation technologies for removing Cd from polluted areas or in biofortification processes.
Many medicinal plants are exposed to heavy metals in their natural habitats due to mining activities and the disposal of processed and unprocessed wastes (Street et al. 2008). Cd is one of the major sources of contamination and is hazardous to plants, affecting their growth and development (Dong et al. 2006; Patra et al. 2004). This work examined the responses of the monocotyledonous plant B. natalensis, which is harvested from the wild for use in traditional medicine, to Cd exposure. The number of roots and the lengths of the roots and shoots produced by Cd-exposed B. natalensis were significantly improved in plants treated with PI-55. This clearly shows the potential utility of PI-55 in alleviating the detrimental effects of Cd exposure. As such, it may be useful for protecting medicinal plants against the effects of heavy metal contamination.
Conclusion
Heavy metal pollution is a serious environmental problem in many countries. New substances such as PI-55 and INCYDE could potentially be used to protect crop plants against heavy metal accumulation or to increase the effectiveness of desirable processes such as biofortification and phytoremediation. In order to use these processes to their full potential, it will be necessary to understand the molecular mechanisms of metal efflux and uptake in plants, and so these should be investigated further. While the exact mechanism by which modulation of cytokinin status protects plants against Cd stress still needs to be resolved, the results presented herein demonstrate that treatment with substances that modulate plants’ cytokinin status can significantly improve seedling establishment under high-Cd conditions.
Abbreviations
- ABA:
-
Abscisic acid
- AHK:
-
Arabidopsis thaliana histidine kinase
- BA:
-
N 6-benzyladenine
- CKX:
-
Cytokinin oxidase/dehydrogenase
- DMSO:
-
Dimethylsulfoxide
- INCYDE:
-
Inhibitor of cytokinin degradation
- JA:
-
Jasmonic acid
References
Alloway BJ (1990) Heavy metals in soils. Wiley, New York ISBN: 0470215984
Alvarez S, Marsh EL, Schroeder SG, Schachtman DP (2008) Metabolomic and proteomic changes in the xylem sap of maize under drought. Plant Cell Environ 31:325–340
Aremu AO, Bairu MW, Novák O, Plačková L, Zatloukal M, Doležal K, Finnie JF, Strnad M, Van Staden J (2012) Physiological responses and endogenous cytokinin profiles of tissue-cultured ‘Williams’ bananas in relation to roscovitine and an inhibitor of cytokinin oxidase/dehydrogenase (INCYDE) treatments. Planta doi: 10.1007/s00425-012-1721-z
Briat JF, Lebrun M (1999) Plant responses to metal toxicity. CR Acad Sci Ser 3 322:43–54
Cary AJ, Liu W, Howell SH (1995) Cytokinin action is coupled to ethylene in its effects on the inhibition of root and hypocotyl elongation in Arabidopsis thaliana seedlings. Plant Physiol 107:1075–1082
Chatfield JM, Armstrong DJ (1986) Regulation of cytokinin oxidase activity in callus tissues of Phaseolus vulgaris L. cv Great Northern. Plant Physiol 80:493–499
Clemens S (2001) Molecular mechanism of plant tolerance and homeostasis. Planta 212:475–486
Dong J, Wu F, Zhang G (2006) Influence of cadmium on antioxidant capacity and four microelement concentrations in tomato seedlings (Lycopersicon esculentum). Chemosphere 64:1659–1666
Gemrotová M, Zatloukal M, Strnad M; Spíchal L (2009) Novel inhibitors of cytokinin oxidase/dehydrogenase and their potential use for in vivo studies. Young Scientist Forum. FEBS Journal doi:10.1111/j.1742-4658.2009.07050.x
Gruz J, Spíchal L (2011) Application of purine derivative LGR-1814 improves functional properties of field-grown lettuce (Lactuca sativa), International PSE Symposium on Phytochemicals in Nutrition and Health, Giovinazzo, Bari, Italy, pp 30
Ha S, Vankova R, Yamaguchi-Shinozaki K, Shinozaki K, Tran LP (2012) Cytokinins: metabolism and function in plant adaptation to environmental stresses. Trends Plant Sci 17:172–179
Hare PD, Van Staden J (1997) The molecular basic of cytokinin action. Plant Growth Regul 23:41–78
Havlová M, Dobrev PI, Motyka V, Štochová H, Libus J, Dobrá J, Malbeck J, Gaudinová A, Vaňková R (2008) The role of cytokinins in responses to water deficit in tobacco plants over-expressing trans-zeatin O-glucosyltransferase gene under 35S or SAG12 promoters. Plant, Cell Environ 31:341–353
Hendry GAF, Baker AJM, Ewart CF (1992) Cadmium tolerance and toxicity, oxygen radical processes and molecular damage in cadmium-tolerant and cadmium-sensitive clones of Holcus lanatus. Acta Bot Neerlandica 41:271–281
Jeon J, Kim NY, Kim S, Kang NY, Novák O, Ku SJ, Cho Ch, Lee DJ, Lee EJ, Strnad M, Kim J (2010) A subset of cytokinin two-component signaling system plays a role in cold temperature stress response in Arabidopsis. J Biol Chem 285:23371–23386
Lukatkin AS, Bashmakov DI, Kipaikina NV (2003) Protective role of thidiazuron treatment on cucumber seedlings exposed to heavy metals and chilling. Russ J Plant Physiol 50:305–307
Lukatkin AS, Gracheva NV, Grishenkova NN, Dukhovskis PV, Brazaitite AA (2007) Cytokinin-like growth regulators mitigate toxic action of zinc and nickel ions on maize seedlings. Russ J Plant Physiol 54:381–387
Metwally A, Tsonev T, Zeinalov Y (1997) Effect of cytokinins on the photosynthetic apparatus in water stressed and rehydrated bean plants. Photosynthetica 34:563–567
Patra M, Bhowmik N, Bandopadhyay B, Sharma A (2004) Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ Exp Bot 52:199–223
Pospíšilová J, Dodd IC (2005) Role of plant growth regulators in stomatal limitation to photosynthesis during water stress, 2nd edition. Revised and Expanded. In: Pessarakli M (ed) Handbook of Photosynthesis. Marcel Dekker, New York, pp 811–825
Sanitá di Toppi L, Gabbrielli R (1999) Response to cadmium in higher plants. Environ Exp Bot 41:105–130
Skoog F, Miller CO (1957) Chemical regulation of growth and organ formation in plant tissue cultured in vitro. Symp Soc Exp Biol 11:118–131
Spíchal L, Werner T, Popa I, Riefler M, Schmülling T, Strnad M (2009) PI-55 is a purine-derived cytokinin antagonist that blocks cytokinin action via receptor inhibition. FEBS J 276:244–253
Street RA, Kulkarni MG, Stirk WA, Southway C, Van Staden J (2007) Toxicity of metal elements on germination and seedling growth of widely used medicinal plants belonging to Hyacinthaceae. Bull Environ Contam Toxicol 79:371–376
Street RA, Kulkarni MG, Stirk WA, Southway C, Van Staden J (2008) Variation in heavy metals and microelements in South African medicinal plants obtained from street markets. Food Addit Contam 25:953–960
Tran LS, Urao T, Qin F, Maruyama K, Kakimoto T, Shinozaki K, Yamaguchi-Shinozaki K (2007) Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis. Proc Natl Acad Sci USA 51:20623–20628
Ul’yanenko LN, Kruglov SV, Filipas AS, Arysheva SO (2004) Effects of growth regulators on barley plant development and accumulation of heavy metals and Caesium-137. Agrokhimiya 12:5–22
Werner T, Motyka V, Strnad M, Schmülling T (2001) Regulation of plant growth by cytokinins. Proc Natl Acad Sci USA 98:10487–10492
Wozny A, Schneider J, Gwozdz EA (1995) The effects of lead and kinetin on greening barley leaves. Biol Plant 37:541–552
Zatloukal M, Gemrotová M, Doležal K, Havlíček L, Spíchal L, Strnad M (2008) Novel potent inhibitors of Arabidopsis thaliana cytokinin oxidase/dehydrogenase. Bioorg Med Chem 16:9268–9275
Zhang P, Wang WQ, Zhang GL, Kamínek M, Dobrev P, Xu J, Gruissem W (2010) Senescence-inducible expression of isopentenyl transferase extends leaf life, increases drought stress resistance and alters cytokinin metabolism in cassava. J Integr Plant Biol 52:653–669
Zhuang P, Ye ZH, Lan CY, Xie ZW, Shu WS (2005) Chemically assisted phytoextraction of heavy metal contaminated soils using three plant species. Plant Soil 276:153–162
Zhuang P, Yang QW, Wang HB, Shu WS (2007) Phytoextraction of heavy metals by eight plant species in the field. Water Air Soil Pollut 184:235–242
Acknowledgments
This work was supported by the University of KwaZulu-Natal, the Academy of Science of the Czech Republic, the Czech Ministry of Education, Youth, and Sports (MSM 6198959216 and LC06034); Grant No. ED0007/01/01 Centre of Region Haná for Biotechnological and Agricultural Research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gemrotová, M., Kulkarni, M.G., Stirk, W.A. et al. Seedlings of medicinal plants treated with either a cytokinin antagonist (PI-55) or an inhibitor of cytokinin degradation (INCYDE) are protected against the negative effects of cadmium. Plant Growth Regul 71, 137–145 (2013). https://doi.org/10.1007/s10725-013-9813-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-013-9813-8