
Abstract
Adaptation to salinity in plant includes not only their response to stress, but also their ability to recover from it. Seeds of Chinese Iris (Iris lactea var. chinensis) collected randomly from an arid area in inland province of Xinjiang (Xj, E93°30′, N42°48′, 740 m), and a semi-arid coastal area in Beijing Municipality (Bj, N40°34′, E116°10′, 540 m) were studied. The objectives were to determine differences between germplasms in ionic relations or accumulation of proline in the shoots and establish how I. lactea adapts to salt stress and subsequent relief. Hydroponically-grown seedlings of the two germplasms were supplied with nutrient solutions containing 0.1 (control), 140 and 280 mM NaCl for 12 days, and supplied subsequently with a non-saline control solution for 12 days to relieve NaCl stress. Germplasm Xj had larger shoot mass and shoot/root ratio than Bj during stress and recovery periods. Salinity, even subsequent recovery, more adversely affected the water content in Bj than Xj. Under identical provision of treatments, the Xj maintained relatively higher K+/Na+ ratio and selectivity for transportation of K+ over Na+ than Bj. Additionally, we observed a slight proline accumulation in shoots of Bj, but its content was insufficient for lowering the solute potential. Germplasm Xj exhibits better adaptation to salinity and subsequent recovery. This is mainly due to its higher selectivity for transporting K+ over Na+. Proline accumulation did not play a key role the osmotic adjustment of the two I. lactea germplasms studied.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Chinese Iris (Iris lactea var. chinensis) is a monocotyledonous halophyte (Zhao and Li 1999) with attractive blooms of the family Iridaceae, and its shoot has forage value as it is grazed by some herbivores. In China, this plant is widely distributed in the arid areas such as western provinces. Even in the saline soil of western provinces, it is grown normally. Thus, the plant is considered an important salt tolerant pioneer species with great potential for cultivation in areas where salinity and drought are becoming a major problem. However, I. lactea germplasms may exhibit different adaptations to salinity or drought, for example, the germplasm collected from inland China (e.g. western provinces) was often found to be relatively drought-resistant because the environments are far more adverse than conditions in the eastern provinces (Shi et al. 2007). Thus, different ecotypes exhibit variability and detailed information on different adaptations of I. lactea germplasms to salinity are important for successful cultivation. Also, after rains or floods, the adverse effect of salt stress on plants are alleviated or reduced. Therefore, the productivity of the plant may depend not only on its response to stress, but also on its capacity to recover from stress. However, there is limited research information on the response of I. lactea to salt stress and its recovery following stress alleviation. Such information could enhance our understanding of the strategy of the species’ survival in saline environments.
Adaptations to salinity in many salt-resistant plant genotypes rely on controlling inorganic ions uptake and transport for instance, a preferential loading of K+ by the stellar (Gorham et al. 1990) and the removal of Na+ during upward transport in the xylem in wheat and barley (Davenport et al. 2005; Garthwaite et al. 2005; Islam et al. 2007). There was limited transport of Na+ and Cl− across the cortex in a tolerant durum wheat genotype, Line 149, compared with a sensitive durum wheat genotype (Läuchli et al. 2008). Furthermore, these inorganic solute concentrations if maintained during salt stress may affect seedling recovery after salt stress relief. In a study on sorghum, a relatively high recovery was exhibited by a salt tolerant genotype after salt stress relief, probably resulting from its greater ability for re-translocation of salts (De Lacerda et al. 2005). The adaptive feature regulating internal ionic relations is the most important both during and after salt stress.
Organic solutes, in particular proline, accumulated under salt stress, and decreased during subsequent recovery for some plants. Concentrations in the leaves of the salt tolerant sorghum decreased significantly, while those in the salt sensitive genotype remained relatively high (De Lacerda et al. 2005). Proline is commonly considered a compatible osmotic solute for balancing the osmotic potential in the cytoplasm with that in vacuoles when plants are under salt stress. The hypothesis is that high contents of proline in leaves could make a significant contribution to the cytoplasmic solute potential after stress relief. Previous work by our team on I. lactea suggested that the plant accumulates a large amount of proline under salt stress (Bai et al. 2008) but quantification of its contribution to osmotic potential of different germplasms, especially at the stress recovery stage has not been done.
In this work, two germplasms of I. lactea were studied for their varied adaptations to salinity, giving an insight into different adaptations to salinity during and after salt stress. The effect of NaCl stress on growth, water status, and absorption of Na+, K+ and Cl− as well as proline content were evaluated. Specifically, our objective was to determine whether differences in the adaptation of the two I. lactea germplasms were related to ionic relations or accumulation of proline in the shoots.
Materials and methods
Seeds of I. lactea were collected randomly from Xinjiang province (Xj, E93°30′, N42°48′, 740 m) or the Beijing Municipality (Bj, N40°34′, E116°10′, 540 m). Characteristics of the seeds are shown in Table 1. The former site is an inland with arid climate, but the latter region has relatively moderate climate with the great majority of its falling in July and August due to the East Asian monsoon, because it is much more coastal (only 149 km far away from Bohai Gulf, East-China Sea). The precipitation and evaporation in Xinjiang province are 34.9 and 2,800 mm in average annually, respectively. But for Beijing, they are 580 and 1,830 mm, respectively. The average annual temperature is 10.0 °C in Xinjiang province (extreme difference in monthly temperature average 37.0 °C) and 8.5 °C in Beijing (extreme difference in monthly temperature average 13.0 °C). The concentrations of water soluble salts in the soils were 2.1 g kg−1 dry soil for Bj and 0.3 g kg−1 dry soil for Xj, respectively.
The experiment was conducted in growth chamber at the College of Resources and Environmental Sciences, China Agricultural University. Day/night temperatures and average relative humidity in the chamber were 24 ± 2/21 ± 1 °C and 60/85 %, respectively. First, the seeds were stored in moistened river sand at 4 °C and soaked using CaSO4 to promote germination. Then, seeds were sown in buckets with acid-rinsed quartz sands under natural light conditions. After emergence, seedlings selected according to uniformity in size and forms were transplanted into holes in a sheet of plastic foam (50 × 30 cm) with three plants per hole. The plastic foam was supported over a large container (50 × 30 × 14 cm) with a nutrient solution containing (mM): K2SO4 0.75, MgSO4 0.65, KCl 0.1, Ca(NO3)2 2.0, KH2PO4 0.25, H3BO30.01, MnSO4 0.001, CuSO4 0.0001, ZnSO4 0.001, (NH4)6Mo2O4 5 × 10−6, Fe-EDTA 0.1. The pH of the nutrient solution was maintained around 6.0 by adding concentrated H2SO4 or KOH. Nutrient solutions were renewed every 2 days.
When the heights of seedlings were about 7.50 cm, a 2 × 2 × 3 factorial experimental design was applied, with the two germplasms, two periods (NaCl stress for 12 days and subsequent recovery for 12 days) and three levels of salinity (0.1 mM NaCl for control, 140 and 280 mM NaCl for two levels of salinity stress). Three replications were used for each treatment and NaCl was applied gradually by adding 70 mM d−1. After attaining the final salinity concentrations, stress treatment was imposed for 12 days followed by another 12 days of recovery from stress (i.e. under control conditions).
All plants were sampled at the end of both stress and recovery periods. First, plants were separated into shoots and roots, and washed with distilled water. After the water on the plant was cleaned with tissue paper, the fresh weight (FW) was immediately recorded. The plant samples were oven dried at 80 °C for 72 h and their dry weights (DW) were measured. Water content (WC) and water deficit (WD) in shoot and root, expressed in milligrams per gram dry weight were calculated from the following equations (Baba and Fujiyama 2003).
where WCCK and WCT represent the water contents (mg g−1) of the control and the stress/recovery treatment, respectively.
Oven-dried samples were powder-homogenized, and 40 mg was ashed in a muffle stove. The ash was dissolved with approximately 0.25 ml of concentrated nitric acid and then adjusted to a volume of 50 ml with deionized water. The contents of Na+, K+ and Ca2+ were determined using ICP-AES. The Cl− content was determined by titration with AgNO3. The ion absorption selectivity ratio (ASK, Na) and transportation selectivity ratio (TSK, Na) were calculated based on the following equations (Bai et al. 2008; Kwon et al. 2009):
For determination of proline, 1.0 g fresh shoots from each of the three replications were homogenized in 10 ml of 30 ml l−1 sulfosalicylic acid. The extract was used for the measurement of proline spectrophotometrically. Net proline was calculated as the difference in proline content between that in the control and that in the stress/recovery treatment. Frozen shoot tissues were put into a syringe to thaw. The liquid squeezed from the tissues was analyzed using a freezing point osmometer (Fiske 110; Advanced Instruments Inc., Norwood, MA, USA) to determine the ic value (the value reading from the instrument). The contribution of proline to the osmotic potential was calculated via the following formulae:
where n is the number of solute molecules; R is the universal gas constant; T is ambient temperature in °K and V is the volume in liter.
Cpro is the contribution of proline to the osmotic potential; Ψpro and Ψs are the osmotic potential of proline and all solutes, respectively; Osmotic coefficient of the solutes in tissue water were assumed to equal 1 (Song et al. 2006).
Statistical analysis was performed with the SPSS statistical program (version11.5, SPSS Inc 2003). The data were subject to a two-way analysis of variance (ANOVA) and treatment means were compared using the least significant difference (LSD) at P = 0.05.
Results
Growth status
The biomass accumulation and water deficit were significantly affected by salinity (Table 2). Furthermore, the differences in shoot biomass and water deficit between the two germplasms were significant under NaCl stress and subsequent recovery (Table 2). After the removal of stress, the shoot biomass accumulation and water deficit differed significantly according to germplasm, salinity level and their interaction; the root/shoot ratio was little affected (Table 2).
Both germplasms showed similar dry mass values when grown in control solution (Table 3). Declines in shoot dry mass and shoot/root ratios were observed at 140 and 280 mM NaCl (Table 3), but the two germplasms differed greatly in their percentage reductions both during and after stress. At 140 mM NaCl, for example, the shoot dry mass of Xj was reduced by 5.6 % compared with 11.7 % for Bj. At 280 mM NaCl, the reduction in shoot dry mass for Xj was 16 % compared with 25 % for Bj. These differences were slightly greater at the end of recovery phase. On recovery from 140 and 280 mM NaCl stress, the decline in shoot dry mass relative to controls values were 11.9 and 31.3 % for Xj and 24.8 and 37.7 % for Bj. As a result, the shoot dry mass in both salt-stressed and salt-recovered seedlings of Xj were much higher at either 140 or 280 mM NaCl (Table 3).
The water deficit of shoots under salinity stress for the two germplasms changed similarly with the shoot dry mass. However, the water deficits in shoots of Bj exposed to NaCl were 1.2- to 1.6-fold higher than in shoots of Xj. The greater difference at the end of recovery phase was due to the optimal water status (i.e. the lowest water deficit) of Xj. A similar trend was found for root water deficit at the end of both stress and recovery periods (Table 3).
All the other ionic parameters with exception of AS–K+/Na+ were significantly affected by salinity (Table 4). Difference in K+, Na+ and selectivity ratio of shoot between the two germplasms was significant under NaCl stress (Table 4). The interaction between germplasm and salinity had significant effect on K+, Na+ and K+/Na+. At the end of the recovery period, salinity affected all the ionic parameters significantly and the K+, Na+, Cl−, K+/Na+ and TS +K +/Na differed considerably between germplasms, The content of Na+ was significantly affected by the interaction of germplasm and salinity (Table 4).
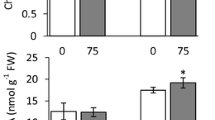
The K+ content decreased with increasing NaCl concentration in shoots of both germplasms (Fig. 1). The lowest K+ content was observed in Bj at 280 mM NaCl level and the K+ content in Xj was higher than in Bj either at 140 or 280 mM NaCl stress (Fig. 1). During the recovery period, the K+ content increased, especially for Xj which increased to 157 % of stress on recovery from 280 mM NaCl, being significantly higher than the 125 % for Bj.

Contents of K+, Na+ and Cl− (mmol g−1) in shoots of two I. lactea germplasms. Error bars represent standard deviation of three replications
The shoot Na+ content increased with increasing external Na+ (Fig. 1) and differences in content between the two germplasms also increased with increasing NaCl. At the 140 mM NaCl level, Na+ content in Bj was 16 % higher than in Xj, while at the 280 mM NaCl, it was 38 % higher. The Na+ content in recovered-seedlings was lower than in stressed seedlings (Fig. 1), but the germplasms, particularly Bj, were not able to decrease their Na+ content to control levels (Fig. 1).
The Cl− content also increased with increasing NaCl supply (Fig. 1), and the maximum Cl− content in shoots was observed in the 280 mM NaCl treatment; the Cl− content in Bj was 5 % higher than that in Xj at the 140 mM NaCl. At the 280 mM NaCl, it was 6 % higher. At the end of the recovery period, the Cl− content differred little between the two germplasms.
The K+/Na+ and Ca2+/Na+ ratios significantly decreased under NaCl stress (Fig. 2a, c). Selectivity ratio (K over Na) of plants increased with increasing NaCl concentration to 140 mM (Fig. 2a, c), but from this external Na+ level to the final level of 280 mM, a further increase occurred only in transportation selectivity ratio (K over Na) (Fig. 2a). The K+/Na+ ratio and its transportation selectivity ratio were higher in Xj than in Bj, especially under stress condition (Fig. 2). Furthermore, the values of transportation selectivity ratio were slightly lower on recovery from NaCl concentration of 280 mM than of 140 mM.

Ionic relations in shoots of two I. lactea germplasms. Error bars represent standard deviation of three replications
Proline content
The content of proline increased with increasing NaCl concentration in shoots of both germplasms. From the Na+ level of 140 mM to the final level of 280 mM, a further increase in net proline accumulation occurred especially for Bj (Fig. 3). Proline concentration in shoots of Bj was 2 % higher than that in Xj at 140 mM NaCl level and 12 % higher at 280 mM NaCl level. At the end of recovery period, these values were 28 and 25 %, respectively.

Net proline accumulation in shoots of two I. lactea germplasms. Error bars represent the standard deviation
There was a negative correlation between shoot proline concentration and shoot dry weight, with this effect particularly marked during stress stage (Fig. 4).

Relationship between the proline content and shoot dry weight of two I. lactea germplasms under salt stress
The contribution of proline to Ψs increased with increasing NaCl concentration for both germplasms (Table 5); however, it was almost below 1 % across treatments for both germplasms despite statistically higher values obtained for Bj.
Discussion
Shoot and root growth
Many dicotyledonous halophytes require quite high concentration of NaCl (Munns and Tester 2008) for optimum growth. This is not the case for the plants used in this study. The optimal growth of I. lactea was achieved at NaCl concentrations of 0.1 mM (Table 2). If its growth is stimulated by salinity, then this is a low concentration of NaCl, which is commonly observed among monocotyledonous halophytes.
Table 2 show the effect of NaCl stress on the dry matter accumulation. Increase in NaCl concentrations in the growth medium led to reductions in shoot biomass of I. lactea. However, responses of the two germplasms to salt stress and subsequent relief were different; differences may reflect their respective salt adaptation strategies. Lutts et al. (1996) found a positive relationship between shoot biomass and salt resistance in rice. A large shoot/root dry matter ratio and its minimal reduction after saline treatment are considered important tolerance indicators of salt stress in sorghum (De Lacerda et al. 2001, 2005). In addition, shoot biomass seem to be related to salt stress acclimation, which fit with the general finding that shoot growth is more sensitive than root growth (Munns and Tester 2008). Undoubtedly, in this study, it was found that the germplasm Xj accumulated significantly higher dry matter than Bj (Table 2), which allow us to characterize Xj as the germplasm more adaptable to salinity.
The water deficit increased with increasing NaCl concentration in both germplasms (Table 2), and the inhibitory effect of osmotic stress was greater at 280 mM NaCl. Interestingly, Xj maintained a favorable water status. Plants under salinity suffer both osmotic stress and specific ion stress. Inhibitory effects of salt on plant growth are at least partially due to the osmotic effect of NaCl (Zhao et al. 2010) and maintenance of an adequate water status may contribute to an improvement in plant growth (Bai et al. 2008). At the 12-day relief stage, the growth recovery of Xj was higher and more rapid. Liu and Staden (2001) suggested that NaCl have a continuing adverse effect on growth even after relief. From our data, we suggest that adequate water uptake is critical for adaptation of I. lactea to saline environments.
Ionic relations
A transfer of plants to a saline culture solution would entail a reduction in K+ content with an increase in external Na+/K+ ratio. However, the responses of the two I. lactea germplasms to different NaCl levels varied. There was significantly higher K+ in Xj than Bj only at 140 mM NaCl during stress and at both 140 and 280 mM NaCl during the relief stage (Fig. 1), suggesting that both salinity and germplasm significantly affected the K+ content. On the other hand, it appeared that the content of K+ in the shoots of I. lactea relative to that of Na+ may be a more important contributing factor to salinity tolerance (Fig. 2a), being consistent with observations for other species, e.g. strawberry (Keutgen and Pawelzik 2009). For Halosarcia pergranulata subsp. Pergranulata and Tecticornia pergranulata, no reduction in tissue K+ was found as NaCl in the medium was raised from 100 to 800 mM (Short and Colmer 1999; Colmer et al. 2009).
Estimates of the selectivity of the absorption or transportation for K+ over Na+ (net ASK:Na or TSK:Na) are available for both halophytes and glycophytes. However, it does not appear that the germplasm Xj exhibited higher ASK, Na than Bj (Fig. 2c, d), suggesting that the salinity tolerance of germplasm Xj was affected by selectivity of the overall transportation for K+ over Na+ rather than absorption of K+ over Na+. De Lacerda et al. (2001) showed that the tolerant genotype of sorghum under salt stress transferred higher amounts of K+ to the shoot and was thus able to maintain an adequate K+ concentration in leaves. The germplasm Xj maintained a higher level of K+-Na+ transportation selectivity than Bj. Although the cause is still unclear, one physiological mechanism, exclusion of Na+, often associated with K+/Na+ has been proposed, as previously observed in Bai et al. (2008) who suggested that the control of Na+ accumulation in shoots of I. lactea could be an important index for identifying salt tolerance. It is generally held that plants maintain a high concentration of K+ and a low concentration of Na+ by regulating the expression of activity of Na+ and K+ transporters as well as the H+ pumps that generate the driving force for transport (Zhu 2003). For example, a stress-inducible membrane protein (AcPMP3) has been proposed to act as a regulator of Na+ and K+ accumulation (Inada et al. 2005) in the monocotyledonous halophyte, Aneurolepidium chinense. Our data suggested that I. lactea transported a limited K+ to shoots, but maintained low Na+ concentrations (Fig. 1); these traits may help the plant adapt to different saline environments.
There was a significantly lower Cl− in Xj than Bj at 140 and 280 mM NaCl levels during the relief stage (Fig. 1). The Na+/Cl− in the shoots, which was low in the control treatment, increased to 4.3 and 4.0 times in Xj and Bj with the application of 280 mM NaCl stress, suggesting that Xj could better manage the Cl− transport under NaCl stress. Also, there were higher accumulate concentrations of Cl− and not Na+ both in Xj and Bj shoots under 140 mM NaCl stress (Fig. 1). Thus, the lower amounts of Cl− transported to shoots play an important role in the mechanism of salt tolerance as observed previously for citrus (Sykes 2011).
Proline content
Proline is commonly considered a compatible osmotic solute for balancing the osmotic potential in the cytoplasm with that in vacuoles when plants are under salt stress. In Triglochin maritima, proline may represent 10–20 % of shoot dry weight (Stewart and Lee 1974). Flowers et al. (1977) suggested that proline occur at sufficiently high concentrations in leaves to contribute to the osmotic pressure in the cell as a whole. Proline could make a significant contribution to the cytoplasmic solute potential if a cytoplasm is confined to equivalent of 10 % of the water content. However, in this study, we did not observe that proline content equaled the osmolarity increase in the vacuole of salt-stressed shoot (Table 5). Moreover, the highest proline content, expressed on a water content basis, was insufficient for lowering the solute potential, even if we assumed that the cytoplasm occupies 10 % of total cellular volume. A similar result was reported for rice (Oryza sativa L.) when plants salinized with Yoshida solution containing NaCl (Lutts et al. 1996). Greenway and Munns (1980) suggested that an adaptive role of proline is related to the survival rather than to the growth of a plant. Song et al. (2006) also suggested that proline may not play an osmotic role in the euhalophyte, Suaeda physophora and the xerophyte, Haloxylon persicum because its contribution to osmotic potential was less than 0.2 %. Similar suggestions were made by Naidoo (2001). Thus, we consider that proline is not involved in osmotic adjustment of salt stressed I. lactea.
Although proline content in the shoots increased with increasing NaCl concentration (Fig. 3), The contribution of proline to Ψs in both germplasms was low across treatments (Table 5), suggesting that proline accumulation would be a reaction to salt stress damage, but not a plant response associated with osmotic adjustment. Furthermore, proline concentration had a negative correlation with shoot biomass expressed as dry weight during stress and relief stages (Fig. 4). This suggests that proline could be used as an indicator of salt stress in I. lactea rather than salt tolerance. Nevertheless, the results in this study do not indicate that proline is merely a symptom of injury. Additional possible regulations should be taken in consideration. During salinity-induced redox stress, proline could also be synthesized from glutamate and ornithine by ∆1-pyrroline-5-carboxylate synthetase (P5CS) and ∆1-pyrroline-5-carboxylate reductase (P5CR). Redox stress will cause an inhibition of photosynthesis. O2 is used as electron acceptor because of the limited NADP+. Proline synthesize is an important defense mechanism for plants to consume excess reducing power from NADH/NADPH in photosynthetic shoots (Hare et al. 1998). Thus, regulation of redox potential could be linked to synthesis and degradation of proline. It may play a role in buffering cellular redox potential (Hare et al. 1998). Proline synthesize maintain NADP+/NADPH ratios at values compatible with metabolism and also play a critical role in antioxidation (Hare and Cress 1997; Hare et al. 1999). Upon relief from stress, proline could support mitochondrial oxidative phosphorylation (i.e. generate reducing equivalents), and probably provide carton, nitrogen and energy (Hare and Cress 1997).
To conclude, compared with Bj, the germplasm Xj exhibits better adaptation and recovery from salinity stress. This is mainly due to its higher selectivity for transportation of K+ over Na+. Proline accumulation did not play a key role in the osmotic adjustment.
References
Baba T, Fujiyama H (2003) Short-term response of rice and tomato to NaCl stress in relation to ion transport. Soil Sci Plant Nutr 49:513–519
Bai WB, Li PF, Li BG, Fujiyama H, Fan FC (2008) Some physiological responses of Chinese Iris to salt stress. Pedosphere 18:454–463
Colmer TD, Vos H, Pedersen O (2009) Tolerance of combined submergence and salinity in the halophytic stem-succulent Tecticornia pergranulata. Ann Bot 103:303–312
Davenport R, James RA, Zakrisson-Plogander A, Tester M, Munns R (2005) Control of sodium transport in durum wheat. Plant Physiol 137:807–818
De Lacerda CF, Cambraia J, Oliva MA, Ruiz HA (2001) Plant growth and solute accumulation and distribution in two sorghum genotypes under NaCl stress. Revista Brasileira de Fisiologia Vegetal 13:270–284
De Lacerda CF, Cambraia J, Oliva MA, Ruiz HA (2005) Changes in growth and in solute concentrations in sorghum leaves and roots during salt stress recovery. Environ Exp Bot 54:69–76
Flowers TJ, Troke PF, Yeo AR (1977) The mechanism of salt tolerance in halophytes. Annu Rev Plant Physiol 28:89–121
Garthwaite AJ, Von Bothmer R, Colmer TD (2005) Salt tolerance in wild Hordeum species is associated with restricted entry of Na+ and Cl− into the shoots. J Exp Bot 56:2365–2378
Gorham J, Jones RGW, Bristol A (1990) Partial characterization of the trait for enhanced K+–Na+ discrimination in the D genome of wheat. Planta 180:590–597
Greenway H, Munns R (1980) Mechanisms of salt tolerance in nonhalophytes. Annu Rev Plant Physiol 31:149–190
Hare PD, Cress WA (1997) Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul 21:79–102
Hare PD, Cress WA, Van Staden J (1998) Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ 21:535–553
Hare PD, Cress WA, Van Staden J (1999) Proline synthesis and degradation: a model system for elucidating stress-related signal transduction. J Exp Bot 50:413–434
Inada M, Ueda A, Shi W, Takabe T (2005) A stress-inducible plasma membrane protein 3 (AcPMP3) in a monocotyledonous halophyte, Aneurolepidium chinense, regulates cellular Na+ and K+ accumulation under salt stress. Planta 220:395–402
Islam S, Malik AI, Islam AKMR, Colmer TD (2007) Salt tolerance in a Hordeum marinum-Triticum aestivum amphiploid, and its parents. J Exp Bot 58:1219–1229
Keutgen AJ, Pawelzik E (2009) Impacts of NaCl stress on plant growth and mineral nutrient assimilation in two cultivars of strawberry. Environ Exp Bot 65:170–176
Kwon TR, Shaheed Siddiqui Z, Harris PJC (2009) Effects of supplemental calcium on ion accumulation, transport and plant growth of salt sensitive Brassica rapa landrace. J Plant Nutr 32:644–667
Läuchli A, James RA, Huang CX, McCully M, Munns R (2008) Cell-specific localization of Na+ in roots of durum wheat and possible control points for salt exclusion. Plant Cell Environ 31:1565–1574
Liu T, Van Staden J (2001) Growth rate, water relations and ion accumulation of soybean callus lines differing in salinity tolerance under salinity stress and its subsequent relief. Plant Growth Regul 34:277–285
Lutts S, Kinet JM, Bouharmont J (1996) Effects of salt stress on growth, mineral nutrition and proline accumulation in relation to osmotic adjustment in rice (Oryza sativa L.) cultivars differing in salinity resistance. Plant Growth Regul 19:207–218
Munns R, Tester M (2008) Mechanisms of salinity tolerance. Ann Rev Plant Biol 59:651–681
Naidoo G, Naidoo Y (2001) Effects of salinity and nitrogen on growth, ion relations and proline accumulation in Triglochin bulbosa. Wetlands Ecol Manage 9:491–497
Shi XX, Mao PC, Zhang GF, Meng L (2007) The comparison of drought resistance on 15 Chinese Iris germplasm at seedling stage. Acta Agrestia Sinica 15:352–358 (in Chinese)
Short DC, Colmer TD (1999) Salt tolerance in the halophyte Halosarcia pergranulata subsp. pergranulata. Ann Bot 83:207–213
Song J, Ding XD, Feng G, Zhang FS (2006) Nutritional and osmotic roles of nitrate in a euhalophyte and axerophyte in saline conditions. New Phytol 171:357–366
Stewart GR, Lee JA (1974) The role of proline accumulation in halophytes. Planta 120:279–289
Sykes SR (2011) Chloride and sodium excluding capacities of citrus rootstock germplasm introduced to Australia from the People’s Republic of China. Sci Hortic 128:443–449
Zhao KF, Li FZ (1999) Halophyte in China. Science Press, Beijing, pp 358–360 (in Chinese)
Zhao KF, Song J, Fan H, Zhou S, Zhao M (2010) Growth response to ionic and osmotic stress of NaCl in salt-tolerant and salt-sensitive maize. J Integr Plant Biol 52:468–475
Zhu JK (2003) Regulation of ion homeostasis under salt stress. Curr Opin Plant Biol 6:441–445
Acknowledgments
This work was supported financially by the National Key Basic Research Program of China (2009CB825101) and the National Natural Science Foundation of China (30870237). We are thankful to Professor Anthony Egrinya Eneji (University of Calabar, Nigeria) and Hai-Min Wang (College of Humanities and Development, China Agricultural University, China) for their critical reading and revision of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zhang, B., Li, PF. & Fan, FC. Ionic relations and proline accumulation in shoots of two Chinese Iris germplasms during NaCl stress and subsequent relief. Plant Growth Regul 68, 49–56 (2012). https://doi.org/10.1007/s10725-012-9692-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10725-012-9692-4