
Abstract
Local patterns of seed regeneration and trade that occur outside the formal breeding sector (seed exchange networks) can have a strong influence on the genetic diversity and evolution of traditional crop varieties. Despite this, little is known about the extent to which seed exchange networks influence gene flow and genetic structure in traditional crop varieties. Here we study barley (Hordeum vulgare subsp. vulgare) in rural communes of Northern Morocco in 2008 and 2009. We quantified seed regeneration and exchange by farmers within the seed exchange network using structured interviews. Using SSR markers, we also quantified the neutral genetic diversity and structure of a complex of traditional varieties referred to as Beldi that is managed in this exchange network. The majority of farmers (>88 %) report cultivating Beldi. Most seeds of Beldi (70–90 %) are maintained on-farm, while the remainder of seeds are obtained from local markets within the commune. Beldi has high genetic diversity and there is weak but significant genetic structure between communes (FST = 0.031). From SSR marker data there is evidence of a high level of gene flow between communes not reported in interviews. Seeds purchased in local markets likely represent seeds from a larger geographic region, leading to lower genetic structure among communes than expected based on the reported level of on-farm seed regeneration and local sourcing of seed. We discuss the implications of this seed exchange network for the conservation of traditional barley varieties in the study region.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Gene flow in plant populations occurs via both seed and pollen dispersal. In wild plant populations, geographic isolation frequently acts as a limit to gene flow and leads to the establishment of genetic structure, often in a pattern of isolation by distance (Wright 1943). In the case of cultivated crop plants, however, gene flow by seed dispersal is usually dependent upon human intervention. For many modern crop varieties, seed dispersal occurs primarily through the formal breeding sector, with genetically homogeneous varieties distributed on a continental or intercontinental scale and cultivated in large-scale monocultures. In large areas of the developing world, however, agroecosystems devoted mainly to traditional crop varieties (often referred to as “landraces”) are the principal drivers of agricultural production, representing over 10 million ha worldwide (Altieri 2004). Farmers with small land holdings and/or agriculturally marginal lands are particularly dependent on traditional varieties that are adapted to local conditions (e.g. low rainfall, soil composition, etc.). These traditional varieties can be considered to be “population-varieties” (Bustarret 1944) because they are genetically heterogeneous and continue to be subjected to the evolutionary forces of mutation, recombination, selection (both natural and artificial), and drift under cultivation. They typically have higher genetic diversity than formal breeding sector varieties and represent important reservoirs of potentially adaptive genes for agriculture.
In many developing countries, farmers maintain traditional varieties independently of the formal breeding sector, with seed often obtained from relatives, neighbours or local markets (McGuire 2008). These informal seed supply systems have been referred to using a variety of terms, including “seed exchange networks”, “farmer seed systems”, “traditional seed systems”, and “informal seed systems”. Here, we will use the term “seed exchange networks”, because it most precisely describes the complex movements of seeds among people and does not make assumptions about the social context in which seed movement is occurring or the identity of the participants in the network. Seed exchange networks are the most important factor in the movement and use of crop germplasm for traditional varieties and they therefore have a strong influence on the genetic diversity and structure of cultivated crops (Hodgkin et al. 2007). As a result, the way that seed is handled and distributed in seed exchange networks has the potential to affect the evolution of traditional crop varieties because it influences processes of gene flow, genetic drift, founder effects, local extinctions, and selection (Almekinders et al. 1994; Brush 1995). This is particularly true for self-fertilizing crops because gene flow via pollen dispersal is restricted (Loveless and Hamrick 1984; Brown and Brush 2000; Epperson 2007; Soleri and Cleveland 2007).
A seed exchange network is structured by many cultural, economical, political, social and geographic factors, all of which may limit or enhance germplasm movement among farmers. It is therefore rare for populations of traditional crop varieties to be structured following a simple pattern of isolation by distance (Pusadee et al. 2009). Instead, they are structured according to interactions among many factors, including agro-climatic zone, altitude, biotic stressors, ethnolinguistic regions, access to markets and agricultural extension services, and marriage networks (Teshome et al. 2001; Mercer et al. 2008; Samberg et al. 2010; Delêtre et al. 2011). If we want to implement effective conservation strategies for a crop in a particular region, it is therefore important to understand both the structure of the seed exchange network and the influence it exerts on the genetic structure of a crop.
Traditional crop varieties have previously been characterized using a number of different methods that can be broadly grouped into two categories: (1) descriptions of seed exchange networks derived from interviews with farmers and, (2) measurements of the genotypic composition and structure of a population made using genotypic markers (Soleri and Cleveland 2001; Pautasso et al. 2012). Studies that quantify seed exchange networks with data gathered from farmer interviews have used such information to infer the likely impact that farmer seed management has on genetic diversity and structure (e.g. Alvarez et al. 2005). The flow of seeds between farmers varies with the crop and the agroecosystem, and ranges from as low as 10 % of seeds or less being sourced off-farm via seed exchange networks for maize in Mexico (Badstue et al. 2007), to as high as 76 % of seeds for rice in the Philippines (Carpenter 2005). It is not generally known, however, if these levels of seed exchange translate to genetic differentiation among populations. Conversely, analysis of genotypes at neutral molecular markers such as simple sequence repeats (SSRs) can provide the best indicators of realized gene flow among populations; yet on the other hand provide no direct information about the seed exchange network that is mediating this gene flow (Parzies et al. 2004).
Although these two approaches are frequently used separately, in a recent review of seed exchange networks, Thomas et al. (2011) concluded that an integration of genetic and ethnobotanical analyses are the key to a quantitative interpretation of how farmers’ practices affect genetic diversity. The few existing studies that have concurrently examined both the seed exchange network and the genetic diversity and structure of traditional crop varieties as assessed with molecular marker data confirm this, and provide useful interpretations of the distribution of genetic diversity in the context of a seed exchange network (Perales et al. 2005; Pandey et al. 2011; Bajracharya et al. 2012; Thomas et al. 2012).
In this study, we focus on a seed exchange network for barley (Hordeum vulgare L. subsp. vulgare) in rural communes of Taounate province in Northern Morocco. In Africa, seed exchange networks are often the major sources of seeds for a community (rather than seeds from the formal sector) with as many as 20–30 % of non-commercial seeds sourced off-farm via seed exchange networks (Tripp and Rohrbach 2001). If this level of off-farm seed sourcing holds true for barley in Morocco, it would suggest a relatively high level of gene flow within the seed exchange network. However, despite the likely importance of the seed exchange network in determining gene flow, few published reports are available detailing the patterns of barley seed exchange for farmers in Morocco, and none of these have used molecular marker data (Mahdi 2002; Rhrib et al. 2002). We used interviews with farmers to identify the varieties of barley cultivated in the study region. Results of interviews revealed that a complex of morphologically similar varieties referred to by farmers as Beldi dominates barley cultivation in the region. We then characterized the seed exchange network for Beldi by identifying and quantifying sources of seed. In conjunction, we characterized the genetic diversity, population structure, and gene flow among populations of Beldi in these communes using SSR markers. Using these tools, we were able to compare and contrast expectations for genetic diversity based on farmer interviews with observed patterns of molecular diversity and thereby obtain a more comprehensive description of gene flow via the seed exchange network. Detailed knowledge of the seed system and associated genetic structure and gene flow can provide a more reliable assessment of the current state of genetic resources for this crop in the region studied. Furthermore, it can help inform the design of effective strategies for both in situ and ex situ conservation.
Methods
Study location
Although Morocco is not a center of origin for barley (Morrell and Clegg 2007), it is considered a center of diversity (Perrino et al. 1986; Dakir et al. 2002), and 85 % of the cultivated barley area is still composed of traditional varieties (Saidi et al. 2006). This proportion increases to over 95 % in semi-arid and marginal sites (Jarvis et al. 2008). Traditional barley varieties in Morocco are distinct from Middle Eastern varieties (Orabi et al. 2009). Most cereal seed cultivated in Morocco is sourced from seed exchange networks, which include seed maintained on farm and seed obtained through exchanges and local markets (89 % of seed), rather than from certified seed sources (11 % of seed) (Van Mele et al. 2011).
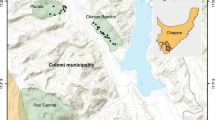
This study was conducted in the Province of Taounate, in the Rif Mountains region of Northern Morocco. This region has a mountainous topography, with severe erosion due to deforestation, overgrazing, and cultivation on steep slopes (Sabir et al. 2002). Precipitation in this region varies between 400 and 600 mm a year. The region is vulnerable to drought, and a severe drought in 2007, the year prior to our study, resulted in a reduced barley harvest with yields decreasing by over 50 % compared to other years (FAOSTAT 2010). In the years of our study (2008 and 2009), the barley harvest was either average (2008), or exceptional (2009) (FAOSTAT 2010). We selected two rural communes for study in 2008: Tissa and Ourtzagh (Fig. 1). For the household surveys in 2009 we added a third commune: Galaz (Fig. 1). Rural communes are administrative divisions that encompass one municipal center and a number of smaller villages. Each commune has a market and an agricultural extension office. Although the communes we studied are situated within the same province, they provide contrasting agro-ecological conditions representative of the region. Tissa commune, in particular, has more farms in more agriculturally productive lowland areas. Ourtzagh and Galaz communes, in contrast, have more farms at higher altitudes, on marginal and sloping terrain. As a result, the study of these contrasting communes should provide a good approximation of diversity patterns and gene flow within the larger region across this agroecological gradient.

The administrative centers of the three rural communes of Taounate Province (Tissa, Ourtzagh and Galaz), where interviews and/or seed sampling were conducted in 2008 and 2009
On-farm interviews and seed collection: 2008
In February of 2008 we met with farmers in the rural communes of Ourtzagh and Tissa to conduct an initial series of interviews and collect samples of the barley varieties cultivated in the region. We visited 10 villages (douars) in Ourtzagh and 9 villages in Tissa (Table 1). These villages were selected to cover a broad area within each commune. Each commune has a weekly local market that is the principal location for seed exchange (Arbaoui 2003). Villages were located at distances ranging between 0.1 and 14.6 km from the market associated with their commune. We interviewed a total of 70 farmers (43 in Ourtzagh, 27 in Tissa). Farmers were randomly selected for interviews that started with an informal discussion, followed by a short, structured, interview (i.e. the interview uses a questionnaire where respondents chose between a selection of answers). Farmers were asked to identify the names of the barley varieties cultivated locally as well as their origin (traditional versus modern). These data were used to determine the overall frequency of use of the different traditional and modern varieties used in the region. During interviews, it became apparent that a traditional variety complex referred to as Beldi dominates barley cultivation in the region. Subsequent questions asked farmers their reasons for preferring to use Beldi compared to other varieties and to identify the sources of the seeds used for planting (e.g. local market, maintained on farm). We also asked farmers to specify the uses made of the barley harvest (e.g. animal fodder, human consumption, etc.).
Following the interview, a small seed sample of approximately 50 g of the variety currently under cultivation was requested from the farmer for further study using SSR markers. Geographic coordinates of each site were recorded using a GPS unit and used to calculate the Euclidean distance between the village and the local market serving the commune.
On-farm interviews: 2009
In March and April of 2009 we conducted more detailed structured individual interviews in the communes of Tissa, Ourtzagh and Galaz to obtain more detailed information regarding barley cultivation and the use and turnover of varieties in the region. A large portion of the questionnaire used in the interviews focused on Beldi, specifically, farmer use and valuation of Beldi, temporal patterns of seed regeneration and sources of gene flow within and among populations Beldi. We revised the questionnaires twice, using test subjects to ensure that the questions were effective for gathering the desired data and to eliminate potential ambiguities. Prior to conducting the interviews, a 5-day training course in participatory rural assessment methods was organised for farmers and community members, with an equal representation of men and women among the course attendees. In addition to providing training, this course fostered the establishment of social relationships and trust among the participants. All the interviewers were selected from the course attendees.
A total of 158 farmers, representing 158 households, were randomly selected for participation in the interviews in the three rural communes: Galaz (N = 63), Ourtzagh (N = 44) and Tissa (N = 51). Interviews were conducted during March and April of 2009 as part of an overall assessment of farmer knowledge and perception of genetic diversity, disease incidence and pest resistance in barley and fava bean crops conducted in collaboration with IAV Hassan II and Bioversity International (Rome, Italy).
Farmers were asked to report the area and number of plots devoted to barley cultivation on their farms. They also reported the different varieties they were cultivating and the area devoted to each variety was then used to calculate Simpson’s diversity index. We assessed change and turnover in barley varieties over time by asking farmers to name the varieties that they currently cultivate and the varieties that they have previously cultivated.
All subsequent interview questions were used for characterising the seed exchange network and, because Beldi dominated the cultivation area, we asked farmers to only report this data for Beldi. Farmers reported how long they had been cultivating Beldi, either as a number of years or as the statement “for a long time”. To determine how often new Beldi seeds are introduced to the Beldi seed stock maintained on-farm, farmers were asked to report the number of years between seed changes (i.e. times when they procure seed from an outside source, such as a market). Some farmers answered, “it depends”, and specified that renewal is dependent on variable external factors (e.g. drought, seed loss during storage), resulting in irregular intervals of seed renewal. Farmers were also asked to report the reason underlying their decision to procure seeds from an outside source. Farmers then reported the volume of seed obtained from each of several potential different sources. This included seed that was maintained on-farm, obtained from neighbours, local markets, and seed companies. The mean percentage of seed obtained from each source was then calculated for each commune.
Genetic diversity analysis
To genotype barley samples, we tested a subset of Beldi seeds collected from farmers during the 2008 interviews. Seeds came from four randomly selected villages in each commune and from two to four farmers within each village. For each sample, three to four seeds were used. This resulted in a total of 84 samples (37 in Tissa, 47 in Ourtzagh) (Table 1). Seeds were germinated and DNA was extracted from a 3 cm segment of the primary leaf using the QIAGEN DNeasy plant mini kit (QIAGEN). DNA samples were genotyped using single sequence repeat (SSR) markers at six loci located on separate chromosomes. The selected loci were: Bmag0013, Bmag0321, Bmag0345, Bmac0316, Bmag0125 and EBmac0541 (Ramsay et al. 2000). Each locus was amplified using sequence-specific primers tagged with an M13 tail and a fluorescently labelled M13 primer (Schuelke 2000). Alleles were resolved on an acrylamide gel using a Li-Cor sequencer (Model 4300) and allele size was quantified using SAGA software (Li-Cor). Detailed methods have previously been described elsewhere (Jensen et al. 2012).
We characterized genetic diversity parameters including allelic richness, and allelic diversity per locus and per site (commune) using the program FSTAT for windows, V 2.9.3.2 (Goudet 2002). Because sample sizes were slightly different between communes, we also performed rarefaction analysis to calculate adjusted allelic richness (standardized to a common sample size) (Kalinowski 2004). For this, we used the program HP-RARE V.1.1 (Kalinowski 2005) and set the number of genes per site to 30, as this was a number slightly lower than that of the site with the lowest number of genes. We also calculated private allelic richness adjusted for population size, as an indicator of the genetic uniqueness of the barley populations in the two communes.
To compare the partitioning of genetic variation among villages and between communes we conducted an analysis of molecular variance (AMOVA) using Arlequin 3.5 (Excoffier and Lischer 2010). To detect both fine-scale local structure and slightly broader structure we calculated pairwise FST-values among villages and between communes using the program MSA (Daniel and Christian 2003). We calculated the number of migrants per generation between the two communes from the values of the fixation index (FST) using the equation, FST = 1/(1 + 4Nem), where Nem represents the absolute number of migrant organisms entering the population each generation (Wright 1943). This method provides an estimate for migration but makes assumptions regarding population parameters that may be violated in real populations. Because barley is inbreeding, random mating and Hardy–Weinberg equilibrium within subpopulations are not expected. Moreover, many generations are required to reach genetic equilibrium between drift and migration, and it is unlikely that it has been established in a crop that is subject to changes in population structure due to human-mediated seed movement.
Because of these potential problems in the calculation of migration rates using FST-based measures, we also used two specialized programs, IMa2 (Hey and Nielsen 2007) and MIGRATE (Beerli and Felsenstein 2001; Beerli 2006), that use the coalescent theory and Bayesian methods to calculate long-term migration rates between populations. IMa2 uses a model that separates population isolation from migration (gene flow), and is therefore particularly appropriate to our data. Furthermore, IMa2 does not assume genetic equilibrium and is appropriate for populations that may have recently separated. MIGRATE does not make this distinction, but is still a useful estimator for long-term gene flow. We calculated population migration rates (M = 2Nem) between communes and effective population sizes scaled for a diploid organism (θ = 4Neμ) within each commune using both programs.
In IMa2, we ran a Bayesian inference with uniform priors for migration rate, theta, and splitting time of the population. Prior values were determined empirically based on outputs of initial runs. A stepwise mutation model was applied. We used a burn-in of 1,000, 20 heated chains with a geometric heating scheme, and swap rates between 40 and 80 % between successive chains. A total of 100,000 samples were collected. Chain mixing and convergence were verified by running successive chains from different random starting points and plotting values of estimated parameters over time.
In MIGRATE, we used the Bayesian inference option with FST-values as starting values for M and Theta and wide uniform priors for both M and Theta. We used the Brownian approximation of the ladder model to account for stepwise mutation in microsatellite markers. The range of priors was determined by a number of preliminary runs to test chain convergence and the range of possible solutions. For the final analysis we ran ten replicates of a long chain with a burn-in of 10,000, sampling increments of 20, and a total of 50,000 genealogies sampled. Slice sampling was used to determine new possible states. Four heated chains were run to explore the parameter space more thoroughly.
To determine the number of genotypic clusters present in Beldi in the region and assess if these clusters were associated with specific geographic locations (i.e., communes), we analyzed the data using SRUCTURE V. 2.2 (Pritchard et al. 2000). We set the number of clusters as ranging from K = 1 through K = 10, with five iterations for each value of K, and a burn-in length and run length of 100,000 each. The most probable number of clusters present in the data was estimated from the probability of the data for each value of K and also by using an ad hoc estimation method (Evanno et al. 2005). Outputs of STRUCTURE analysis were plotted using DISTRUCT (Rosenberg 2004).
Results
On-farm interviews, 2008
During informal discussion in 2008, farmers identified the two major varieties cultivated in the region as Beldi and Roumi. Both are six-row types, but Beldi is considered by farmers to be the traditional local variety. In Arabic, Beldi translates as “local” or “of the land”, while Roumi is considered to be an imported variety derived from the formal breeding sector. In contrast to Beldi, the word Roumi translates as “foreign” or “exotic”, possibly deriving from the word “European”, or “Roman”. The simple distinction between the names Beldi and Roumi may, however, hide a more complex reality. Because Beldi means “local”, the term could potentially refer to a number of morphologically distinct varieties, rather than one specific variety, particularly across a broad geographic area. In the case of our study, farmers identified a suite of morphological traits that allow them to distinguish Beldi from other varieties, including 6-row spikes, seed morphology, low spike density, a high proportion of vegetative biomass, and tall stature. There remains the possibility, however, that Beldi also encompasses other morphological types not identified in this study. Furthermore, farmers sometimes conceptualize Beldi as a series of distinct local populations, in which case the name of an individual farmer is used to qualify a particular sample of Beldi. For these reasons, we have referred to Beldi throughout this paper as a “complex of traditional varieties”, rather than as a single variety. All the other varieties identified during interviews, but not studied extensively, are referred to as “named varieties”. In both communes, the majority of farmers (88.9 % in Tissa, 97.7 % in Ourtzagh) cultivate Beldi (Table 2). The remaining barley is almost exclusively Roumi. There were no significant differences between communes for use frequency of Beldi (χ2 = 2.37, df = 1, p = 0.12) or of Roumi (χ2 = 1.04, df = 1, p = 0.31).
The majority of barley seeds of all varieties were maintained on-farm in both communes (Fig. 2). This proportion was lower in Tissa than in Ourtzagh (82 % compared to 93 %), although not significantly so (χ2 = 2.18, df = 1, p = 0.14). The remaining seeds were mostly purchased or traded in markets. The distance from the villages to the nearest market varied from 0.1 to 14.6 km. When considering only seeds of Beldi, the proportion of seeds maintained on-farm was nearly identical (93 % in Tissa and 92 % in Ourtzagh) in both communes.

The sources of the seeds used for planting in 2008 in the rural communes of Ourtzagh and Tissa. Data is presented for all varieties combined (top) and for the Beldi variety complex only (bottom). The majority of seeds were maintained on-farm, but a small proportion of seeds was obtained from outside sources in both communes. The seeds obtained from outside sources in Tissa commune is higher for all varieties combined than for Beldi considered alone
The major reason cited by farmers in both communes for preferring to cultivate Beldi was “high productivity” (67 and 64 % in Tissa and Ourtzagh, respectively) (Table 3). In Ourtzagh, the second most important reason was “resistance to drought” (31 %); this was significantly higher than in Tissa (only 8.5 %) (χ2 = 5.13, df = 1, p = 0.02). Conversely, farmers in Tissa cited “cost effectiveness” (50 %), which was significantly more than farmers citing this reason in Ourtzagh (24 %) (χ2 = 4.66, df = 1, p = 0.03). Similar proportions of farmers in both communes cited local adaptation as a factor in their variety choice (approximately 21 %).
The use made of the barley harvest in both communes was principally as animal fodder (both grain and vegetative biomass) and as seed for planting the following year (Table 4). A high proportion of farmers (58 and 63 % in Ourtzagh and Tissa, respectively) sold a portion of their seed production in local markets. Barley was used for human consumption by 33 and 42 % of farmers in Ourtzagh and Tissa, respectively. Some farmers reported setting aside a portion of the harvest specifically for use as seed in the following year. This practice was significantly more common in Ourtzagh (21 %) than in Tissa (4 %) (χ2 = 4.02, df = 1, p = 0.04).
On-farm interviews, 2009
Farmers in all three communes cultivated barley in a number of small plots (less than one hectare in size), with total cultivation area and plot size being larger in Tissa than in Galaz or Ourtzagh (Table 5). Farmers cultivated a total of six barley varieties or variety complexes, with Beldi dominating cultivation in all communes (97, 95 and 87 % of cultivation area in Galaz, Ourtzagh and Tissa, respectively) (Table 6). Farmers did not report the use of variety mixtures as a strategy to increase yield or yield stability. The modern variety Roumi was the second most important cultivar, followed by Rouiza (a 2-row variety whose name can be translated as “small grain”), while the remaining three varieties (six-row Rouiza, Touinssi roumi and Stati) were extremely rare, each being reported by only one farmer. The names attributed to these rare varieties provide some indication of their characteristics and/or origin. Six-row Rouiza refers to a small-grained variety with six rows. Touinssi roumi denotes a foreign variety of Tunisian origin and Stati refers to a variety originating from the town of Settat, in the Gharb valley region of Morocco. Although the current varietal richness of six is relatively high, values for Simpson’s diversity index, which represents the probability that two randomly selected individuals will belong to the same variety, were also high, indicating low diversity. This is due to the dominance of Beldi. Tissa had the highest varietal diversity and richness of the three communes. A comparison of varieties currently cultivated and varieties no longer cultivated (Table 7), showed that most farmers in all three communes (over 95 %) currently cultivate Beldi. The use of Roumi is increasing in all three communes, with more farmers currently cultivating Roumi than farmers who report discontinuing use of Roumi. Conversely, use of Rouiza is decreasing in all communes, with much higher percentages of previous use than of current use. There is also evidence of turnover in the varieties available in the region, with six-row Rouiza, Touinssi roumi and Stati representing newly introduced varieties while a free-threshing variety named Chaïr Nabi (i.e. “the Prophets’ barley”) and a variety named Zbiti represent varieties that were previously cultivated in the region but are no longer available.
In the case of Beldi, the majority of farmers in all three communes report that they have been cultivating it “for a long time” (Fig. 3a), often stating that they inherited their Beldi seeds from their parents. Many farmers in each commune (between 25 and 60 %) report that they never renew their seed stock of Beldi from outside sources (Fig. 3b). This was the most frequent response in both Ourtzagh and Tissa communes, although in Galaz commune the majority of farmers report renewing their seed every 2 years. The remaining farmers reported periodically using outside seeds to renew or supplement their seed supply. These responses were mostly distributed between seed renewals every year to every 5 years, indicating relatively frequent influxes of new seed. Some farmers (2–12 %) also indicated that frequency of seed renewal depends on outside factors and, as such, is variable.

The patterns of use of Beldi over time represented as the number of years that farmers report cultivating Beldi (a) and the frequency with which farmers renew their supply of Beldi seeds from outside sources (b). (Data from the 2009 interviews)
Farmers report a number of reasons for renewing their seeds of Beldi from outside sources (Table 8). The most frequent reasons cited included that the farmers’ seeds were mixed with seeds of other, undesirable, species (i.e. weeds), the seed supply was insufficient, or the seeds had poor germination. The number of farmers citing insufficient seed supply was significantly higher in Galaz than in Ourtzagh or Tissa (χ2 = 7.104, df = 2, p = 0.03). On average, for each farmer, 70–90 % of the Beldi seeds planted in 2009 came from stocks that had been maintained on-farm (“self”) (Fig. 4). The remaining seeds were sourced from local markets, neighbours, seed companies, or non-local markets. There were no significant differences among the three communes in the percentage of seeds obtained from these different sources.

The sources of Beldi seeds used for planting in 2009 in the rural communes of Galaz, Ourtzagh and Tissa. The percentage of seeds obtained from each source is expressed as a percentage (+SE). There are no significant differences among communes
The seed exchange network in 2008 and 2009
Overall, the seed exchange network is similar in each of the three communes studied and is also similar in both years studied. The Beldi variety complex has been cultivated for a long time in the region and is maintained almost exclusively through an informal seed exchange network, with very few seeds purchased from seed companies. Farmers in all communes produce most of the seeds used for planting on-farm (over 90 % in 2008 and over 75 % in 2009). Most of the remainder of seed is purchased in local markets. A relatively small proportion of farmers reserve a specific portion of the harvest for use as seed in the following year, but most do not reserve seed specifically for this purpose and simply plant the new crop using the seed remaining from the previous year.
The reasons cited by most farmers for obtaining seed from outside sources reflect problems with their own seed source, such as having insufficient seeds, a seed stock that is contaminated with too many weed seeds, or poor germination. There does not appear to be a conscious choice to renew their seed from outside sources in the hopes of obtaining new adaptive traits or to increase the diversity of their own seed stock. Although many farmers report never needing to use seed from outside sources, many others report using outside seeds, mostly at intervals varying from every year to every 5 years.
Genetic diversity, gene flow and genetic structure in Beldi
In samples of Beldi, between 6 and 8 alleles were identified for each SSR locus (Table 9). Values of gene diversity were high, and similar in both communes with most values being above 0.6. Similarly, allelic richness was nearly identical between Tissa and Ourtzagh, especially when rarefaction analysis was used to compensate for differences in sample size between communes (approximately 4.8 alleles per locus are expected in a sample size of N = 30 for each commune). Private alleles were present and there was similar private allelic richness in each commune. Overall, private allelic richness was relatively low, with approximately one private allele per locus expected in samples of the same size from each commune. Observed heterozygosity was extremely low, as is expected for a crop with an inbreeding mating system (results not shown). FST-values between villages were not significant (results not shown). In contrast, the FST-value measuring genetic differentiation between communes was low, but significant (FST = 0.031, p = 0.02). The number of migrants per generation between the two communes as calculated from the FST value was very high (Nem = 7.81). Similarly, results of AMOVA found a small (1.9 %), but significant proportion of variation partitioned between communes, whereas the variation partitioned among villages within a commune (12 %) was not significant (Table 10). The majority of variation (86 %) was present within villages within communes, indicating high diversity maintained in each village. Bayesian analysis of population migration rates between communes using IMa2 identified similar and high levels of gene flow (with 95 % confidence intervals) occurring in both directions between the communes [MT→O = 2.12 (0.28, 9.14) and MO→T = 2.82 (0.80, 8.60)]. Analysis with MIGRATE gave values for population migration rates similar to those obtained using IMa2 [MT→O = 1.53 (0.05, 8.77) and MO→T = 2.13 (0, 9.29)]. The rates of mutation-scaled migration (mT→O = 1.08) and mO→T = 1.72) in the MIGRATE analysis were similar to the mutation scaled population-sizes (θTissa = 1.78 and θOurtzagh = 2.54) indicating extremely high rates of historical gene flow between the two communes.
The most probable number of clusters identified using STRUCTURE was K = 4. A relatively high proportion of genotypes from Tissa (frequency = 0.421) were assigned to cluster 3, while genotypes from Ourtzagh were more likely to be assigned to cluster 1 (frequency = 0.374) or cluster 4 (frequency = 0.337) (Fig. 5). Fewer genotypes overall were assigned to cluster 2, but it was much more frequent in Tissa (frequency = 0.151) than in Ourtzagh (frequency = 0.052). These clusters indicate the presence of some geographic structure between communes, but there is also considerable overlap between communes, with some individuals in each commune assigned to each of the four clusters.

Population structure of Beldi seeds from Tissa and Taounate communes estimated using genotypic clustering in STRUCTURE for K = 4 clusters (red cluster 1, yellow cluster 2, blue cluster 3, green cluster 4). (Color figure online)
Combined results of interview and genotype data
The combination of interview data and genotypic data results in a more comprehensive depiction of seed exchange networks in Ourtzagh and Tissa (Fig. 6). Because the IMa2 model conforms more closely to our data than the MIGRATE model, we have elected to report the IMa2 values in the figure representing gene flow in the seed exchange network (Fig. 6). Although interview data identified the likely presence of gene flow among farms within a commune, genotypic data identified, in addition, a high level of gene flow between communes that was not reflected in the interviews data.

The Beldi seed exchange network and gene flow within and between the rural communes of Tissa and Ourtzagh. The sources of seed (indicated as percentages of total seed used in a year) are derived from interview data. The seeds sourced within the village include seeds renewed on-farm and seeds obtained from a neighbour in the same village. Population migration rates (the rate at which genes from one commune are supplanted by genes from the other commune) are calculated from SSR data using IMa2 and are reported as M values (M = 2Nem) with 95 % confidence intervals. Farmers in both rural communes report using over 80 % of their own seed and buying the remainder from local markets within the commune
Discussion
Diversity of barley in the study region
In all three communes, the diversity of the named barley varieties identified in interviews is relatively low because a complex of morphologically similar traditional varieties identified by the name Beldi occupies most of the cultivation area. For this reason, we have focused primarily on Beldi in our analyses and discussion. A variety named Beldi in this region generally represents a traditional variety that has been cultivated by most farmers for so long that they are unable to report the specific time when they started cultivating it. Most farmers report obtaining their original stock of Beldi seeds from their parents, further confirming that varieties identified by this name have been present in the region for a long time. Farmers value Beldi because it has high productivity in the region and is resistant to drought. As a result, Beldi is considered reliable, cost effective and of good market value. Furthermore, the Beldi plant has a higher proportion of vegetative biomass than Roumi and therefore provides good quality animal fodder, which is a major use of the barley crop in the region, even more important than use as human food.
Beldi populations on individual farms have high genetic diversity and low structure, with each village containing a large proportion of the diversity of the population as a whole. This is similar to results for barley in other agro-ecosystems. In central Morocco, an evaluation of traditional varieties using storage proteins (hordeins) as markers also identified high diversity (Dakir et al. 2002). In Eritrea, a study using SSR markers also found high diversity and low structure, with the average individual field retaining 97 % of the diversity of the population (Backes et al. 2009). By contrast, in Tunisia, barley varieties also have high diversity, but show some structuring according to geographic origin (Raoudha et al. 2007).
The large percentage of farmers cultivating Beldi indicates that farmers are maintaining a high use and valuation of Beldi, despite the availability of other named varieties, including varieties from the formal breeding sector. However, the slight increase in the use of Roumi in all communes indicates that Roumi has replaced Beldi in a small portion of the barley cultivation area. Further studies of this region in future years will be necessary to determine if use of Roumi will continue to increase, and potentially displace Beldi. We believe it is likely, however, that Beldi will remain important in this region. The continued use of traditional varieties despite the availability of other varieties has been observed in other major crops, including maize in Mexico (Perales et al. 2003) and wheat in Turkey (Brush and Meng 1998). In these cases, genetic erosion of traditional varieties is inhibited because the local varieties are uniquely adapted to both agro-ecological conditions and cultural needs (Bellon 1996). As a result, their role in the local agro-ecosystem is not easily replaced and they provide better food security than most varieties from the formal breeding sector. This is particularly the case for barley in Morocco, where the different parts of the plant fill a large number of niches in traditional Moroccan farming systems. The grain provides both fodder for livestock (80 % of production) and food for humans (20 % of production) (Ceccarelli et al. 2001). In addition, the quality of barley straw is important because it provides livestock feed during the dry season (Rhrib and Amri 2005) and can represent a significant proportion of the crop value (Annicchiarico and Pecetti 2003). Beldi typically has better quality straw and much more straw biomass. Farmers in this region also practice “green stage grazing”, allowing livestock to graze the barley plants in the early stages of growth and then allowing the plants to recover to produce tillers and grains (Belaid and Morris 1990; Johnson 1996). This is an advantageous practice that can increase the overall biological yield of the crop by 1,000–2,000 kg annually without reducing grain yield (Yau 2003). Very few modern cultivars can perform as well as traditional cultivars under this practice (Anderson 1985; Yau and Mekni 1987). These multiple uses are reflected in farmer selection criteria, which differ from formal breeding sector criteria that prioritize semi-dwarf, high grain-yielding varieties (Ceccarelli et al. 2001; Yau 2003).
The seed exchange network for Beldi
By combining interview data with genotypic data, we have obtained a more comprehensive characterization of genetic diversity and gene flow within the seed exchange network than with either method used alone. Because of this, we have focused our discussion of the seed exchange network for Beldi in Tissa and Ourtzagh communes, where both types of data are available. In each commune, in a given year, approximately 10–20 % of the seed supply is purchased in local markets, while the remainder of the seeds are from the farmers’ own seed supplies. These influxes of seed should lead to a moderate level of gene flow within the commune.
There are some differences in the valuation and conservation of seed for Beldi between Tissa and Ourtzagh, that may be related to the differences in agroecological conditions between the two communes. In particular, the marginal conditions in Ourtzagh may increase vulnerability to drought, which may explain the much higher number of farmers in Ourtzagh who cite drought resistance as a reason for cultivating Beldi. Farmers in Ourzagh were also much more likely to reserve a portion of their seeds for planting the following year. This could be because they are more likely to experience low yields (due to smaller cultivated areas, poor soil, and higher vulnerability to drought), and have therefore adopted this practice to help mitigate against years with a poor harvest.
The use of outside seeds does not appear to be higher in 2008 (the year following a poor harvest after a drought year in 2007) than in 2009 (the year following a good harvest). This is likely because some seeds from the 2006 harvest remained from the harvest in the year prior to the drought and these were used to plant the 2008 crop. In Ourtzagh, this may be partly due to the farmer practice of reserving a portion of the harvest specifically for planting. This conservation of seeds for planting was also undoubtedly helped by aid provided to farmers by the Moroccan government after the drought. Free sacks of barley were distributed to farmers through the local agricultural extension offices. Farmers used these seeds to feed their livestock, allowing them to retain their own seed for planting. If, however, the government does not provide assistance after a drought, or there are poor harvests for two or more subsequent years, seed reserves may be exhausted, leading to higher influxes of outside seed.
Based solely on the results of interview data, gene flow appears to be restricted to the scale of the commune because it occurs mainly between the local market and farms within the commune. There is also a small influx of seed from the formal breeding sector (seed companies) in the Tissa commune. However, the very small volume of seed from the formal sector is indicative of the overall dominance of seed exchange networks as the main source of seed.
Data from molecular markers indicates that gene flow is also occurring on a larger scale than the commune, with a high number of migrants between Beldi populations in the two communes, as indicated both by classical population genetic tests and estimates of gene flow using Bayesian methods. High gene flow between communes is also corroborated by the low private allelic richness in each commune. This gene flow between communes is high enough that genetic structure between communes, although present, is weak.
Because the interview data do not directly indicate the cause of gene flow between communes, we must consider a number of different explanations. Gene flow between communes could occur through three potential routes: (1) vendors who sell seed in both of the local markets, (2) farmers from one commune travelling to the market in the other commune to purchase seed, or (3) farmers from different communes purchasing or trading seeds amongst themselves. It is also possible that the gene flow detected represents the signature of common ancestry of the Beldi populations in both communes, rather than ongoing gene flow, though this seems a less likely explanation because our model distinguishes population isolation from gene flow.
We believe that route (1), vendors selling seed in more than one local market, is most likely to account for gene flow. This type of gene flow would be facilitated by the structure of traditional rural markets in Morocco. These markets are periodic, i.e. they convene once a week in each market place, and the day of the week for the markets in a region are staggered, allowing one vendor or one farmer to attend more than one market. Historically, the markets have attracted vendors and buyers from a small radius of approximately 20–30 km due to poor road conditions (Mikesell 1958). However, with improvements in roads and transportation (Buerli et al. 2008), this radius may have expanded, resulting in increased long-distance gene flow, although recent data is not available. This is likely the mechanism for high gene flow between communes, and suggests that genetic structure in the region may have decreased in recent years, compared to times when travel was more difficult and vendors would have been limited to fewer markets. The low genetic structure of Beldi among communes is therefore consistent with the levels of seed exchange and gene flow reported by farmers, given that market seed is likely to represent seed from a broader region than the commune. Similarly, a study of traditional maize varieties in Mexico (an outcrossing species), found that a reported 5 % influx of seed per year from outside the community was sufficient to obscure any evidence of neutral genetic structuring between communities (Perales et al. 2005).
The other two explanations for gene flow among communes (farmers travelling to markets in other communes and farmers obtaining seed from farmers in other communes) are less likely to be significant because they would have been identified in the farmer interviews. On the questionnaire, farmers had the option to identify either “local markets” or “non-local markets” as the source of their seed, so farmers purchasing from non-local markets as a source of gene flow would have been detected during the interviews. Similarly, on the questionnaire, farmers had the option to identify “friend or relative from another village” as a source of seed and this was not reported, making it unlikely that significant levels of gene flow occurred via direct trade between farmers in different communes. Gene flow between communes occurring due to vendors selling in both local markets, however, was not accounted for in the survey questions.
The seed exchange network for barley appears to be similar to that for fava bean (Vicia faba L.), a legume crop that is cultivated in rotation with barley in the region. In the case of V. faba, most seed exchanges happen locally, between neighbours or relatives in the same area. However, some seed exchanges happen on a larger scale, when farmers purchase from vendors representing a larger geographic area (Arbaoui 2003). It is therefore likely that these two interconnected crops share similar patterns of genetic diversity and genetic structure.
A recent study of traditional barley varieties in high-hill agro-ecosystems in Nepal identified levels of seed exchange for two different barley varieties similar to those reported in this study, with 20–25 % of barley seeds obtained through exchanges with other farmers in a given year (Bajracharya et al. 2012). This, coupled with frequent renewal of seed stocks following crop failure led to a similar lack of genetic differentiation, particularly in marginal and poor environments. The parallels between our findings and those for barley in Nepal likely reflect the similar environments in which traditional barley varieties are cultivated, i.e. agriculturally marginal environments that are unsuitable for higher value crops such as wheat or rice.
Conservation implications
Close to 100 % of barley seed stocks in the region are managed via the seed exchange network with very few inputs from the formal seed sector. The seed exchange network is therefore critical to barley supply and diversity in the region, even more so than for other cereals in Morocco (mainly wheat) where it accounts for 89 % of seed (Van Mele et al. 2011). As a result, any vulnerability in the seed exchange network will likely have an impact on the diversity and evolution of Beldi.
This study highlights some vulnerable points in the seed exchange network that should be considered in the context of crop yield stability and food security in the region, as well as for in situ conservation of barley diversity in this region. In particular, the small proportion of farmers who set aside seed specifically for use in planting the following year is a cause for concern because in the case of a year (or a series of years) with a poor harvest (e.g. due to drought, fungal disease, or insect pests), farmers may use all their seeds prior to planting time, and may then be forced to purchase seeds from outside sources. These seeds may not be of high quality or may come from a different region and be poorly adapted to local conditions. This effect is currently mitigated by government aid programs that provide additional seeds to farmers for feeding their livestock after a poor harvest, allowing farmers to retain some of their own seeds for planting in the following year.
Even in a year with a good harvest, however, the remaining seeds from the previous harvest that are used for planting the new crop may not be the highest quality seeds, resulting in poor germination, or poor yield. This vulnerability in the seed system is already apparent in the number of farmers who periodically renew their seed supply from outside sources because their own seed is insufficient or of poor quality rather than because of a deliberate choice to acquire new seeds. By comparison, in other seed exchange networks, there are frequent reports of farmers deliberately choosing to renew their seeds in search of new traits (e.g. culinary properties, early maturity), to increase quality and yield, or as a component of a social ritual (e.g. a gift or exchange at a wedding) (e.g., Asfaw et al. 2007; Delaunay et al. 2008).
Additional factors that cause decreased yields and poor quality harvest, such as climate change, are likely to compound this vulnerability in the seed system and may have an influence on the continued evolution of Beldi in the region (Pautasso et al. 2012). Morocco has been subject to drier conditions and prolonged periods of drought since the 1980s (Esper et al. 2007). In this context, it is especially important for farmers to reserve a portion of their stored seed for planting the crop in the following year. Additional interventions that can help mitigate loss of seed stores due to adverse environmental conditions include community seed banks, diversity-kits, diversity fairs, and micro-credit schemes (Jarvis et al. 2011). Any strategies that involve the development of new community seed storage facilities will, however, require good design and monitoring to avoid seed loss to pests (e.g. insects and rodents) and/or poor storage conditions (e.g. excess heat and humidity).
Future research directions
This study points to a number of questions that merit further study. The combined use of interview data and molecular markers has identified local markets as a potential source of cryptic long-distance gene flow. In future studies, assessing the diversity and sources of seeds available in local markets would provide valuable information regarding sources of gene flow. Also, although we only detected weak genetic structure on the basis of neutral molecular markers, it is possible that markers for adaptive traits or loci would show some structuring across an agro-climatic gradient. Further research should therefore investigate differences in adaptive traits, in addition to neutral markers. It is also possible that we did not detect strong structure because the spatial scale investigated was too small and future studies might detect stronger barriers to gene flow if samples are collected from a much larger geographical area.
Here, we have identified Beldi as a traditional variety complex that has been cultivated in the region for a long time. A suite of common morphological characteristics is consistently associated with the varieties identified as Beldi in the study region. However, because the variety name simply means “local”, it is possible that the variety considered to be local has changed over time, but the name has remained the same. Work on traditional crop varieties in Morocco has demonstrated that, with increasing geographic distance, varieties with the same names tend to differ morphologically (Sadiki et al. 2007). It is likewise possible that with increasing time, varieties with the same name will also differ morphologically. Further work using samples collected at previous times and stored in seed banks will be required to ensure that the same morphological variety is being referred to and that Beldi is, in fact, representative of varieties that have historically been cultivated in the region.
Conclusions
Future studies of barley in this region should focus more extensively on the complex of varieties identified as Beldi, given the high importance and valuation of Beldi, rather than on the other named varieties identified in this study. This will allow better understanding of the nature of this variety complex, in particular the extent of morphological divergence among different populations of Beldi. The use of a combination of interview and molecular data in the study of traditional crop varieties allows a more comprehensive assessment of gene flow via the seed exchange network than either method used alone. In this case, combining both methods allowed us to identify moderate to high levels of gene flow both within and between communes. It is advisable to combine both types of data, particularly in systems that are not yet well characterized. Insights obtained using this approach may then be used to refine future studies and conservation strategies. Finally, the seed exchange network in this region is likely to be vulnerable to years with poor harvests, which may be compounded by the effects of climate change and periodic drought. Efforts at in situ conservation of barley in this region should aim to increase the proportion of farmers and/or communities maintaining a reserve supply of seeds.
References
Almekinders CJM, Louwaars NP, de Bruijn GH (1994) Local seed systems and their importance for an improved seed supply in developing countries. Euphytica 78:207–216
Altieri MA (2004) Linking ecologists and traditional farmers in the search for sustainable agriculture. Front Ecol Environ 2:35–42
Alvarez N, Garine E, Khasah C, Dounias E, Hossaert-Mckey M, McKey D (2005) Farmers’ practices, metapopulation dynamics, and conservation of agricultural biodiversity on-farm: a case study of sorghum among the Duupa in Sub-Sahelian Cameroon. Biol Conserv 121:533–543
Anderson WK (1985) Production of green feed and grain from grazed barley in Northern Syria. Field Crops Res 10:57–75
Annicchiarico P, Pecetti L (2003) Developing a tall durum wheat plant type for semi-arid, Mediterranean cereal-live stock farming systems. Field Crops Res 80:157–164
Arbaoui M (2003) Analyse des facteurs évolutifs de la diversité génétique de la fève (Vicia faba L.) in situ et leurs impacts sur son maintien et sa gestion à la ferme: contribution au développement des bases scientifiques pour la conservation in situ de la fève au Maroc. Dissertation, Institut Agronomique et Vétérinaire Hassan II, Département d’Agronomie et d’Amélioration des Plantes
Asfaw A, Almekinders C, Getahun D, Fistum A (2007) Bean seed flow and exchange networks in Southern Ethiopia. Seed Info 33:14–17
Backes G, Orabi J, Wolday A, Yahyaoui A, Jahoor A (2009) High genetic diversity revealed in barley (Hordeum vulgare) collected from small-scale farmer’s fields in Eritrea. Genet Resour Crop Evol 56:85–97
Badstue LB, Bellon MR, Berthaud J, Ramirez A, Flores D, Juarez X (2007) The dynamics of farmers’ maize seed supply practices in the Central Valleys of Oaxaca, Mexico. World Dev 35:1579–1593
Bajracharya J, Brown AHD, Joshi BK, Panday D, Baniya BK, Sthapit BR, Jarvis DI (2012) Traditional seed management and genetic diversity in barley varieties in high-hill agro-ecosystems of Nepal. Genet Resour Crop Evol 59:389–398
Beerli P (2006) Comparison of Bayesian and maximum-likelihood inference of population genetic parameters. Bioinformatics 22:341–345
Beerli P, Felsenstein J (2001) Maximum likelihood estimation of a migration matrix and effective population sizes in n subpopulations by using a coalescent approach. Proc Nat Acad Sci 98:4563–4568
Belaid A, Morris ML (1990) Wheat and barley production in rainfed marginal environments of West Asia and North Africa: problems and prospects. CIMMYT Economics working paper 91, 76 pp
Bellon MR (1996) The dynamics of crop infraspecific diversity: a conceptual framework at the farmer level. Econ Bot 50:26–39
Brown AHD, Brush SB (2000) The genetic structure of crop landraces and the challenge to conserve them in situ on farms. In: Brush SB (ed) Genes in the field. Lewis Publishers, IDRC and IPGRI, Ottawa, pp 19–48
Brush SB (1995) In situ conservation of landraces in centers of crop diversity. Crop Sci 35:346–354
Brush SB, Meng E (1998) Farmers’ valuation and conservation of crop genetic resources. Genet Resour Crop Evol 45:139–150
Buerli M, Aw-Hassan A, Rachidi YL (2008) The importance of institutions in mountainous regions for accessing markets: an example from the Moroccan High Atlas. Mount Res Dev 28:233–239
Bustarret J (1944) Variétés et variations. Ann Agron 14:336–362
Carpenter D (2005) The in situ conservation of rice plant genetic diversity: a case study from a Philippine Barangay. Agric Hum Values 22:421–434
Ceccarelli S, Grando S, Bailey E, Amri A, El-Felah M, Nassif F, Rezgui S, Yahyaoui A (2001) Farmer participation in barley breeding in Syria, Morocco and Tunisia. Euphytica 122:521–536
Dakir EH, Ruiz ML, Garcia P, De La Vega MP (2002) Genetic variability evaluation in a Moroccan collection of barley, Hordeum vulgare L., by means of storage proteins and RAPDs. Genet Resour Crop Evol 49:619–631
Daniel D, Christian S (2003) Microsatellite analyser (MSA): a platform independent analysis tool for large microsatellite data sets. Mol Ecol Notes 3:167–169
Delaunay S, Tescar RP, Oualbego A, vom Brocke K, Lançon J, Haut-Pont L (2008) Growing cotton does not disrupt traditional sorghum seed exchange. Cah Agric 17:189–194
Delêtre M, McKey DB, Hodkinson TR (2011) Marriage exchanges, seed exchanges, and the dynamics of manioc diversity. Proc Nat Acad Sci 108:18249–18254
Epperson BK (2007) Plant dispersal, neighbourhood size and isolation by distance. Mol Ecol 16:3854–3865
Esper J, Frank D, Büntgen U, Verstege A, Luterbacher J, Xoplaki E (2007) Long-term drought severity variations in Morocco. Geophys Res Lett 34:L17702
Evanno G, Regnaut S, Goudet J (2005) Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol 14:2611–2620
Excoffier L, Lischer HEL (2010) Arlequin suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour 10:564–567
FAOSTAT (2010) Food and Agriculture Organization of the United Nations Website: http://faostat.fao.org
Goudet J (2002) FSTAT: a program to estimate and test gene diversities and fixation indices. v. 2.9.3.2. (Online) Available at http://www.unil.ch/izea/softwares/fstat.html
Hey J, Nielsen R (2007) Integration within the Felsenstein equation for improved Markov chain Monte Carlo methods in population genetics. Proc Nat Acad Sci 104:2785–2790
Hodgkin T, Rana R, Tuxill J, Balma D, Subedi A, Mar I, Karamura D, Valdivia R, Collado L et al. (2007) Seed systems and crop genetic diversity in agroecosystems. In: Jarvis DI, Padoch C, Cooper HD (eds) Managing biodiversity in agricultural ecosystems. Columbia University Press, New York, pp 77–116
Jarvis DI, Brown AHD, Cuong PH, Collado-Panduro L, Latournerie-Moreno L, Gyawali S, Tanto T, Sawadogo M, Mar I et al. (2008) A global perspective of the richness and evenness of traditional crop-variety diversity maintained by farming communities. Proc Nat Acad Sci 105:5326–5331
Jarvis DI, Hodgkin T, Sthapit BR, Fadda C, Lopez-Noriega I (2011) An heuristic framework for identifying multiple ways of supporting the conservation and use of traditional crop varieties within the agricultural production system. Crit Rev Plant Sci 30:125–176
Jensen HR, Dreiseitl A, Sadiki M, Schoen DJ (2012) The Red Queen and the seed bank: pathogen resistance of ex situ and in situ conserved barley. Evol Appl 5:353–367
Johnson DL (1996) Development trends and environmental deterioration in the agropastoral systems of the central Middle Atlas, Morocco. In: Swearingen WD, Bencherifa A (eds) The North African environment at risk. Westview Press, Inc., Boulder, pp 35–54
Kalinowski ST (2004) Counting alleles with rarefaction: private alleles and hierarchical sampling designs. Conserv Genet 5:539–543
Kalinowski ST (2005) Hp-rare 1.0: a computer program for performing rarefaction on measures of allelic richness. Mol Ecol Notes 5:187–189
Loveless MD, Hamrick JL (1984) Ecological determinants of genetic structure in plant populations. Annu Rev Ecol Syst 15:65–95
Mahdi M (2002) Effets des flux et de la circulation des semences et des produits sur la conservation des variétés locales au Maroc. In: Birouk A, Sadiki M, Nassif F, Saidi S, Mellas H, Bammoun A, Jarvis DI (eds) La conservation in situ de la biodiversité agricole: un défi our une agriculture durable. IPGRI, Rome, pp 176–194
McGuire SJ (2008) Securing access to seed: social relations and sorghum seed exchange in eastern Ethiopia. Hum Ecol 36:217–229
Mercer K, Martinez-Vasquez A, Perales HR (2008) Asymmetrical local adaptation of maize landraces along an altitudinal gradient. Evol Appl 1(3):489–500
Mikesell MW (1958) The role of tribal markets in Morocco: examples from the “Northern Zone”. Geogr Rev 48:494–511
Morrell PL, Clegg MT (2007) Genetic evidence for a second domestication of barley (Hordeum vulgare) East of the Fertile Crescent. Proc Nat Acad Sci 104:3289–3294
Orabi J, Jahoor A, Backes G (2009) Genetic diversity and population structure of wild and cultivated barley from West Asia and North Africa. Plant Breed 128:606–614
Pandey A, Bisht IS, Bhat KV, Mehta PS (2011) Role of informal seed system in promoting landrace diversity and their on-farm conservation: a case study of rice in Indian Himalayas. Genet Resour Crop Evol 58:1213–1224
Parzies HK, Spoor W, Ennos RA (2004) Inferring seed exchange between farmers from population genetic structure of barley landrace Arabi Aswad from Northern Syria. Genet Resour Crop Evol 51:471–478
Pautasso M, Aistara G, Barnaud A, Caillon S, Clouvel P, Coomes O, Delêtre M, Demeulenaere E, De Santis P et al. (2012) Seed exchange networks for agrobiodiversity conservation. A review. Agron Sustain Dev. doi:10.1007/s13593-012-0089-6
Perales HR, Brush SB, Qualset CO (2003) Landraces of maize in Central Mexico: an altitudinal transect. Econ Bot 57:7–20
Perales HR, Benz BF, Brush SB (2005) Maize diversity and ethnolinguistic diversity in Chiapas, Mexico. Proc Nat Acad Sci 102:949–954
Perrino P, Polignano GB, Sui-Kwong J, Khouya-Ali M (1986) Collecting germplasm in Southern Morocco. Plant Genet Resour Newsl 65:26–28
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics 155:945–959
Pusadee T, Jamjod S, Chiang YC, Rerkasem B, Schaal BA (2009) Genetic structure and isolation by distance in a landrace of Thai rice. Proc Nat Acad Sci 106:13880–13885
Ramsay L, Macaulay M, degli Ivanissevich S, MacLean K, Cardle L, Fuller J, Edwards KJ, Tuvesson S, Morgante M et al. (2000) A simple sequence repeat-based linkage map of barley. Genetics 156:1997–2005
Raoudha A, Hatem CM, M’Barek BN, Chaabane R, Leîla B-K, Jeanette BH (2007) Simple sequence repeat markers and physiological characterization of some local Tunisian barley accessions. J Cell Mol Biol 6:19–29
Rhrib K, Amri A (2005) Contribution to the elaboration of in situ conservation strategies of Moroccan barley populations. In: Jordan A, Amri A, Damania A, Shehadeh A, Noaman M, Jana S (eds) Promoting community-driven conservation and sustainable use of dryland agrobiodiversity, ICARDA, Aleppo, Syria, p 80
Rhrib K, Amri A, Sadiki M (2002) Caractérisation agro morphologique des populations locales d’orge des sites Tanant et Taounate. In: Birouk A, Sadiki M, Nassif F, Saidi S, Mellas H, Bammoun A, Jarvis DI (eds) La conservation in situ de la biodiversité agricole: un défi our une agriculture durable. IPGRI, Rome, Italy, pp 286–294
Rosenberg NA (2004) Distruct: a program for the graphical display of population structure. Mol Ecol Notes 4:137–138
Sabir M, Maddi M, Naouri A, Barthes B, Roose E (2002) Runoff and erosion risks indicators on the main soils of the Mediterranean Mountains of Occidental Rif Area (Morocco). In: Conference proceedings: 12th International Soil Conservation Organization (ISCO) conference, Beijing, China, pp 370–375
Sadiki M, Jarvis DI, Rijal D, Bajracharya J, Hue NN, Camacho-Villa TC, Burgos-May LA, Sawadogo M, Balma D et al. (2007) Variety names: an entry point to crop genetic diversity and distribution in agroecosystems? In: Jarvis DI, Padoch C, Cooper HD (eds) Managing biodiversity in agricultural ecosystems. Columbia University Press, New York, pp 34–76
Saidi S, Amri A, Jarvis D, Sbeih D, Saad A (2006) On-farm conservation of barley landraces in Morocco. In: Conference proceedings: African Regional workshop on sustainable use (focus on agriculture), Nairobi, Kenya, ICARDA, Aleppo, Syria
Samberg LH, Shennan C, Zavaleta ES (2010) Human and environmental factors affect patterns of crop diversity in an Ethiopian Highland agroecosystem. Prof Geogr 62:395–408
Schuelke M (2000) An economic method for the fluorescent labeling of PCR fragments. Nat Biotechnol 18:233–234
Soleri D, Cleveland DA (2001) Farmers’ genetic perceptions regarding their crop populations: an example with maize in the Central Valleys of Oaxaca, Mexico. Econ Bot 55:106–128
Soleri D, Cleveland DA (2007) Isolation by distance under diverse systems of mating. J Cell Mol Biol 31:39–59
Teshome A, Brown ADH, Hodgkin T (2001) Diversity in landraces of cereal and legume crops. Plant Breed Rev 21:221–261
Thomas M, Dawson JC, Goldringer I, Bonneuil C (2011) Seed exchanges, a key to analyze crop diversity dynamics in farmer-led on-farm conservation. Genet Resour Crop Evol 58:321–338
Thomas M, Demeulenaere E, Dawson JC, Khan AR, Galic N, Jouanne-Pin S, Remoue C, Bonneuil C, Goldringer I (2012) On-farm dynamic management of genetic diversity: the impact of seed diffusions and seed saving practices on a population-variety of bread wheat. Evol Appl. doi:10.1111/j.1752-4571.2012.00257.x
Tripp R, Rohrbach D (2001) Policies for African seed enterprise development. Food Policy 26:147–161
Van Mele P, Bently JW, Guéi RG (2011) African seed enterprises: sowing the seeds of food security. CABI, Oxfordshire
Wright S (1943) Isolation by distance. Genetics 28:114–139
Yau S-K (2003) Yields of early planted barley after clipping or grazing in a semiarid area. Agron J 95:821–827
Yau SK, Mekni MS (1987) Breeding dual-purpose barley. Field Crops Res 15:267–276
Acknowledgments
Funding for this work was provided by the Natural Sciences and Engineering Research Council of Canada in the form of a Canada Graduate Scholarship (to H.R.J.) and in the form of a Discovery Grant (to D.J.S.), by the International Development Research Center in the form of a Doctoral Research Award (to H.R.J.), by the Canadian Foundation for Innovation in the form of the Leaders Opportunity Fund (to D.J.S), by Bioversity International, and by IAV Hassan II. Inputs from Moroccan researchers and site level teams and farmers were funded through grants provided by the United Nations Environmental Programme/Global Environmental Facility (UNEP/GEF), the United Nations Food and Agriculture Organization (FAO), and the Swiss Agency for Development and Cooperation (SDC). Field collection of barley seeds in Morocco was completed with the help of Maria El Ouatel and Mohammed Abou El Ouafa. Training sessions and farmer interviews were conducted with Maria El Ouatel, Ghita Chleyah, Hafida Hamida, Manana Saïd, Salsabil Taoufiqi, Mariam El Bedraoui, Naoual Touati, Siham Atfaoui, Fatima Boujema and Bouchra Sadiqi. We also thank the local site teams who participated in this research and the farmers who provided the information and seed materials for this study. Characterization of SSR markers was completed with the help of Nella Mauro. Helpful advice was provided by Abderrahman Khila and Adam Herman. Two anonymous reviewers provided helpful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Jensen, H.R., Belqadi, L., De Santis, P. et al. A case study of seed exchange networks and gene flow for barley (Hordeum vulgare subsp. vulgare) in Morocco. Genet Resour Crop Evol 60, 1119–1138 (2013). https://doi.org/10.1007/s10722-012-9909-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-012-9909-4