
Abstract
Spelt wheat is a neglected crop that could be used in the quality breeding of modern common wheat. One important aspect of this quality is the starch composition which is related to the waxy proteins. A collection of 420 accessions of Spanish spelt wheat was analysed for waxy protein composition by SDS–PAGE. Polymorphism was found in the three waxy proteins, detecting differences both in size and in activity, and a new waxy allele (Wx-D1g) was identified. Seed amylose content was also determined and significant differences were detected among the different allelic combinations. In general, the accessions carrying one or two waxy null alleles showed less amylose content. The variation found could be used to enlarge the genetic pool of common wheat, or to develop lines of spelt with different levels of amylose content.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Starch is the main component of the wheat grain. Starch granules consist of two types of glucose polymers, the essentially linear amylose and the highly branched amylopectine, in a ratio in the range of 22–35%/68–75%. In addition, there are lipids and proteins linked to the starch granules although they are present in small quantities (Schofield and Greenwell 1987). The proteins associated with the wheat starch granules have been widely studied and these days their function is well known (Morell et al. 2001). The most important of these proteins, or at least the most studied, is the granule-bound starch synthase I (GBSSI) or waxy protein. This protein is the enzyme responsible for amylose synthesis in the seed. Since some starch properties such as gelatinization, pasting and gelation depend on the amylose/amylopectine ratio (Zeng et al. 1997), the waxy protein is crucial for flour quality.
Three waxy proteins are present in hexaploid wheats: Wx-A1 (located on chromosome 7AS), Wx-B1 (chromosome 4AL) and Wx-D1 (chromosome 7DS) (Yamamori et al. 1994). Several studies have been carried out looking for polymorphism in wheat waxy proteins. The diversity found is low, even more when compared with other wheat seed proteins such as the storage proteins. Nonetheless, various forms of the waxy proteins have been found including null alleles, which are of particular interest. Nakamura et al. (1993) crossed two wheat cultivars lacking one and two different waxy proteins to get waxy wheat with amylose-free starch. This kind of starch has been reported to be better for the noodles quality (Oda et al. 1980) and more efficient than standard wheat if the seed is used as substrate for biofuel production (Wu et al. 2006). Furthermore, resistant starch (high amylose content) is required by the food industry to create healthier foods, because resistant starch is less well digested in the small intestine than standard one (Topping and Clifton 2001). Therefore, the search for different forms of the waxy proteins is interesting in order to provide new resources for breeding programmes focused on starch properties.
One useful gene pool for wheat breeding is the hulled wheats (Padulosi et al. 1996). Our team has carried out several studies directed toward the evaluation of genetic resources of these underutilized and neglected wheats, mainly focussed on materials of Spanish origin. In the case of one of these, spelt (Triticum spelta (T. aestivum L. em. MacKey ssp. spelta (L.) Thell.), we have studied a large Spanish collection for the variation of morphological traits and seed storage proteins in hulled wheats (Caballero et al. 2001, 2004, 2008a, b; Alvarez et al. 2007). This species is still grown in Asturias (North of Spain), where, recently, a collecting mission was carried out to evaluate the existing genetic variability of this crop (Caballero et al. 2007).
The objective of the current study was to evaluate the polymorphism of the waxy proteins, as well as its possible effects on the amylose content, in the same Spanish spelt wheat collection previously evaluated for morphological and seed storage proteins content by our team.
Materials and methods
Plant materials
Four hundred and twenty accessions of Spanish spelt wheat obtained from the National Small Grain Collection (Aberdeen, USA), Centre for Genetic Resources (Wageningen, Netherlands) and the Plant Genetic Resource Center-INIA (Centro de Recursos Fitogenéticos-INIA, Alcalá de Henares, Spain), were analysed in this study. Most of these accessions are landraces and were collected in Asturias (province in the north of Spain). These plants were grown during 2006–2007 and 2007–2008 in the IFAPA station (Cordoba, Spain) according to standard agricultural practice in the region. Several spikes per plant were protected to avoid random crosses. Chinese Spring and Mexicali cultivars were used as standards.
Starch extraction and electrophoretic analysis
Twenty milligrams of flour were mixed with 1 ml of distilled water and incubated at 4°C for 24 h. The homogenate was filtered through Miracloth and centrifuged at 14,000 g for 1.5 min. The pellet was washed with 1 ml of buffer A [55 mM Tris–HCl pH 6.8, 2.3% (w/v) sodium dodecyl sulphate, 2% (w/v) dithiotreitol, 10% (v/v) glycerol], according to Echt and Schwartz (1981). Then, 1 ml of buffer A was added to the pellet and was left for 30 min at room temperature. The pellet was washed three times with distilled water, once with acetone and then air-dried. The residue was mixed with 80 μl of buffer A, heated in a boiling bath for 2 min, cooled in ice and centrifuged.
An aliquot of supernatant (20 μl) was loaded in vertical SDS–PAGE slabs in a discontinuous Tris–HCl-SDS buffer system (pH: 6.8/8.8) at a polyacrylamide concentration of 12% (w/v, C = 0.44%). The Tris–HCl/glycine buffer system of Laemmli (1970) was used. Electrophoresis was performed at a constant current of 30 mA/gel at 18°C for 4 h after the tracking dye migrated off the gel. Protein bands were visualised by silver staining.
Amylose analysis
All samples for each waxy combination found were evaluated for apparent amylose content (Table 2). This content was determined in triplicate as described by Watanabe et al. (1998).
Statistical analyses
Allelic frequency and the effective numbers of alleles per locus (Ne) at the Wx-A1, Wx-B1 and Wx-D1 loci were calculated. Genetic diversities was estimated using Nei’s diversity index -He- (Nei 1972, 1973).
Data on amylose content were analysed by ANOVA, and means were compared using Duncan’s multiple range test.
Results
Polymorphism at GBSSI or waxy proteins
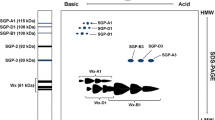
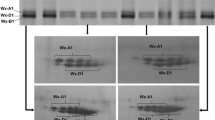
The electrophoretic analysis of the waxy proteins revealed polymorphism for the three waxy loci (Fig. 1; Table 1). For the Wx-A1 locus, two alleles were found: the Wx-A1a, similar to the allele present in cv. Chinese Spring, and the Wx-A1b (null type), which lacks the protein (lanes 1, 2 and 5). The Wx-A1b allele was present in 36 accessions (8.57%).

SDS–PAGE gel electrophoresis patterns of waxy proteins. Lanes are as follow: 1, PI-348544 (Wx-A1b, Wx-B1b, Wx-D1a,); 2, BGE-012911 (Wx-A1b, Wx-B1a, Wx-D1a); 3, BGE-012913 (Wx-A1a, Wx-B1b, Wx-D1a); 4, PI-348693 (Wx-A1a, Wx-B1a, Wx-D1b); 5, PI-348701 (Wx-A1b, Wx-B1a, Wx-D1g); 6, cv. Chinese Spring (Wx-A1a, Wx-B1a, Wx-D1a); 7, PI-348495 (Wx-A1a, Wx-B1c′, Wx-D1a,); and 8, cv. Chinese Spring (Wx-A1a, Wx-B1a, Wx-D1a)
For the Wx-B1 locus, three different alleles were found: the Wx-B1a as Chinese Spring (lane 6); the null protein Wx-B1b (lane 3); and the Wx-B1c′ allele, with less mobility than Wx-B1a (lane 7). Both Wx-B1b and Wx-B1c′ alleles were less common (11.67 and 12.86%).
The lines evaluated presented a large homogeneity for the Wx-D1 locus, where 99.52% of the accessions showed the Wx-D1a allele. Two additional alleles were detected: the null allele (Wx-D1b, lane 4) that was only present in one accession; and a novel waxy allele (lane 5), not previously described, with a slightly lower electrophoretic mobility than the Wx-D1a allele. This new allele was provisionally named Wx-D1g.
The effective number of alleles (Ne) was especially low for the Wx-D1 locus, where one of the detected alleles (Wx-D1a) was clearly dominant over the others. This indicates that the other two of the alleles detected are in clear danger of erosion. In addition, the He value for this locus was very low (Table 1), representing approximately 10% of the genetic diversity that this locus would have if the allelic variants were distributed randomly.
Considering the variation for the Wx-A1, Wx-B1 and Wx-D1 loci, eight combinations were detected. Their frequencies are shown in Table 2, where the clear dominance of the combination Wx-A1a, Wx-B1a, Wx-D1a can be observed; it appears in 69.52% of the samples. For the rest, three combinations appeared with a relatively high frequency, the combination Wx-A1a, Wx-B1c′, Wx-D1a appears in 47 accessions, Wx-A1a, Wx-B1b, Wx-D1a in 44 and Wx-A1b, Wx-B1a, Wx-D1a in 23. The other four combinations were rare with frequencies of under 5% (Table 2).
Apparent amylose content
Apparent amylose content was measured in all accessions evaluated, which were grouped according to waxy allelic combination (Table 2). An analysis of variance was carried out using the allelic combination as variation source. The differences between accessions per each combination were not significant, while the differences between combinations were significant. The results from comparison of means by the Duncan’s multiple range test is shown in Table 2. In general, higher values were found in the accessions with the three active waxy alleles, while the accessions carrying one or two null waxy alleles had lower values.
Discussion
In this study, a wide collection of Spanish spelt wheat was analysed to study polymorphism in the waxy proteins. This collection represents a substantial proportion of the Spanish spelt wheat accessions conserved in Germplasm Banks.
Although the polymorphism found was low, different forms for each waxy protein (Wx-A1, Wx-B1 and Wx-D1) were detected. Null forms of each locus were detected. These are very important for breeding programmes focused on the development of waxy wheats (amylose-free starch). Several accessions contained the Wx-A1b (8.75%) or Wx-B1b (11.63%) proteins, these results being similar to those found by Rodriguez-Quijano et al. (1998) in a similar study focused on T. aestivum and T. spelta Spanish wheats (Wx-A1b, 7.63%; Wx-B1b, 12.50%). The detection of the null allele for the Wx-D1 locus is particularly important. This allele is extremely rare and has not been described previously in spelt wheat. As far as we know this null allele has been detected previously only in five accessions of common wheat (Yamamori et al. 1994; Urbano et al. 2002). As cv. Bai Huo (Wx-D1b) is repeatedly used in breeding programmes, the discovery of new materials carrying the Wx-D1 null allele is important in order to enlarge the range of possible parentals used in waxy wheat production.
In the cases of the Wx-B1 and Wx-D1 proteins differences in size were also detected between the different accessions. One of the alleles found (Wx-D1g) has not been described previously and was only present in one line. Another well-known allele, Wx-B1c′, was found in several accessions, with a similar frequency (12.82%) to that described by Yamamori et al. (1994) in their study with T. aestivum accessions from all over the world (15.70%) and higher than that described by Rodriguez-Quijano et al. (1998) in a study on Spanish spelt (3.47%).
Apparent amylose content was determined in samples of each allelic combination of waxy loci detected in order to determine whether the differences found affect the amylose content. Although the method used overestimates the amylose values it seems very useful to compare the different genotypes. Significant differences were found between some allelic combinations; in particular it seems evident that samples carrying one or two null waxy alleles have less amylose than those carrying the three waxy proteins, which is in agreement with the results of Yamamori and Quynh (2000). These authors also established that the lack of Wx-A1 protein had less effect than the lack of Wx-B1 or Wx-D1 proteins. Although the differences between accessions Wx-A1 null and Wx-D1 null or Wx-B1 null were not significant, the accessions lacking Wx-D1 protein or Wx-B1 protein showed lower mean values, supporting the data from Yamamori and Quynh (2000), who found that Wx-B1 protein is the protein which generates most amylose and Wx-D1 the second one.
In conclusion, the current study has demonstrated that the safeguard of these spelt accessions, together with others stored in Germplasm Banks, is fundamental for the maintenance of the genetic diversity in this crop. This diversity could be of special relevance for plant breeding, mainly for starch quality. In addition, spelt is an interesting genetic resource for the search of useful genes for plant breeding of the modern wheats, and could enlarge the gene pool of waxy proteins in common wheat.
References
Alvarez JB, Caballero L, Martín LM (2007) Variability for morphological traits and high-molecular-weight glutenin subunits in Spanish spelt lines. Plant Genet Resour Charact Util 5:128–130
Caballero L, Martín LM, Alvarez JB (2001) Allelic variation of the HMW glutenin subunits in Spanish accesions of spelt wheat (Triticum aestivum ssp. spelta L. em. Thell.). Theor Appl Genet 103:124–128
Caballero L, Martín LM, Alvarez JB (2004) Variation and genetic diversity for gliadins in Spanish spelt wheats accesions. Genet Resour Crop Evol 51:679–686
Caballero L, Martín LM, Alvarez JB (2007) Agrobiodiversity of hulled wheats in Asturias (North of Spain). Genet Resour Crop Evol 54:267–277
Caballero L, Martín LM, Alvarez JB (2008a) Genetic diversity in Spanish populations of Triticum spelta L. (escanda): example of an endangered genetic resource. Genet Resour Crop Evol 55:675–682
Caballero L, Bancel E, Debiton C, Branlard G (2008b) Granule-bound starch synthase (GBSS) diversity of ancient wheat and related species. Plant Breed 127:548–553
Echt CS, Schwartz D (1981) Evidence for the inclusion of controlling elements within structural gene at the waxy locus in maize. Genetics 99:275–284
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680–685
Morell MK, Rahman S, Regina A, Appels R, Li Z (2001) Wheat starch biosynthesis. Euphytica 119:55–58
Nakamura T, Yamamori M, Hirano H, Hidaka S (1993) Production of waxy (amylose-free) wheats. Mol Gen Genet 248:253–259
Nei M (1972) Genetic distances between different populations. Am Nat 106:283–292
Nei M (1973) Analysis of gene diversity in subdivided populations. Proc Natl Acad Sci USA 70:3321–3323
Oda M, Yasuda Y, Okazaki S, Yamauchi Y, Yokoyama Y (1980) A method of flour quality assessment for Japanese noodles. Cereal Chem 54:253–254
Padulosi S, Hammer K, Heller J (1996) Hulled wheats. International Plant Genetic Resources Institute, Rome
Rodriguez-Quijano M, Nieto-Taladriz MT, Carrillo JM (1998) Polymorphism of waxy proteins in Iberian hexaploid wheats. Plant Breed 117:341–344
Schofield JD, Greenwell P (1987) Wheat starch granule proteins and their technological significance. In: Morton ID (ed) Cereals in a European context. Ellis Horwood, Chichester, pp 407–420
Topping DL, Clifton PM (2001) Short chain fatty acids and human colonic function-roles of resistant starch and non-starch polysaccharides. Physiol Rev 81:1031–1064
Urbano M, Margiotta B, Colaprico G, Lafiandra D (2002) Waxy proteins in diploid, tetraploid and hexaploid wheats. Plant Breed 121:465–469
Watanabe N, Noda Y, Goto N, Miura H (1998) Variation for apparent amylose content of endosperm starch in Triticum durum and Aegilops squarrosa. Euphytica 101:283–286
Wu X, Zhao R, Wang D, Bean SR, Seib PA, Tuinstra MR, Campbell M, O`Brien A (2006) Effects of amylase, corn protein and corn fiber contents on production of ethanol from starch-rich media. Cereal Chem 83:569–575
Yamamori M, Quynh NT (2000) Differential effects of Wx-A1, -B1 and -D1 protein deficiencies on apparent amylose content and starch pasting properties in common wheat. Theor Appl Genet 100:32–38
Yamamori M, Nakamura T, Endo R, Nagamine T (1994) Waxy protein deficiency and chromosomal location of coding genes in common wheat. Theor Appl Genet 89:179–184
Zeng M, Morris CF, Batey II, Wrigley CW (1997) Sources of variation for starch gelatinization, pasting, and gelation properties in wheat. Cereal Chem 74:63–71
Acknowledgments
This research was supported by grant AGL2007-65685-C02-02 from the Spanish Ministry of Science and Innovation and the European Regional Development Fund (FEDER) from the European Union. The first author thanks to the Spanish Ministry of Education and Science (FPU programme) for a predoctoral fellowship and Josefina Sillero for field help.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Guzmán, C., Caballero, L., Moral, A. et al. Genetic variation for waxy proteins and amylose content in Spanish spelt wheat (Triticum spelta L.). Genet Resour Crop Evol 57, 721–725 (2010). https://doi.org/10.1007/s10722-009-9507-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-009-9507-2