
Abstract
The stable isotope technique and the difference method are common approaches for estimating fertiliser N uptake efficiency. Both methods, however, have limitations and their suitability may depend on N management and environmental conditions. A field experiment was conducted on a humus sandy soil in northern Germany to estimate fertiliser N uptake efficiency of silage maize in the year of application (Zea mays L.) by the stable isotope and the difference method as influenced by the type of N fertiliser (mineral vs. cattle slurry), the application mode (separate or combined application), and N rate. Seven N treatments were included (0, 50, 100 and 150 kg mineral N ha−1; 20, 40 m³ cattle slurry ha−1; 50 kg mineral N ha−1 plus 40 m³ slurry ha−1), where either mineral N or slurry N was labelled, and mineral N was split into two dressings. In addition, 4.1 kg ha−1 labelled mineral N was incorporated into otherwise unlabelled treatments (0, 20, 40 m³ ha−1, and 50 kg mineral N ha−1 plus 40 m³ ha−1) to estimate N uptake from the upper soil layer. Uptake of 15N was followed in leaves, stalk, ear, and the whole crop. Fertiliser N uptake efficiency (FNUE15N) of mineral fertiliser N obtained by the isotope technique ranged between 51 and 61%. Recovered fertiliser N was mainly found in the ear, while less labelled N remained in leaves and the stalk. The nitrogen rate tended to increase the amount of recovered N, but the effect was not consistent among plant parts and the whole crop. Plant N uptake from non-fertiliser N was found to increase N input up to 100 kg N ha−1. Nitrogen recoveries of the two mineral N dressings were similar for the different plant parts as well as for the whole crop. Fertiliser N uptake efficiency (FNUEdiff) of mineral N estimated by the difference method resulted in substantially higher values compared to FNUE15N, varying between 56 and 98%. More N was taken up from the upper soil layer with increasing N supply, which is regarded as a major error source of the difference method. Slurry N was taken up less efficient in the year of application than mineral fertiliser N as indicated by recovery rates of 21–22% (FNUE15N) and 39–62% (FNUEdiff), respectively. When mineral N and slurry were applied together, the difference method estimated significantly lower N uptake efficiencies for both mineral and slurry N compared to a single application, while values obtained by the isotope method were not affected.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In northwestern Europe, silage maize is a key component of ruminant diets in intensive dairy farming due to its high yield and metabolic energy content (O’Mara et al. 1998), and its acreage has increased substantially over the last few decades. Although maize has a high N use efficiency (Schmitt and Edwards 1981), field balances still show considerable N surpluses due to excessive input of organic and mineral fertilisers, which are applied alone or in combination (Neeteson 2000; Schröder et al. 2005; Haas et al. 2005). Adjusting N application rates to crop needs can improve N use efficiency and reduce N leaching. To this end, indicators for assessing the N status of crops or soil can provide valuable information (Schröder et al. 2000; Herrmann and Taube 2005; Samborski et al. 2009). However, nitrogen application tailored to crop demand requires accurate knowledge of fertiliser N recovery, which is especially challenging with respect to slurry since short-term and residual N effects have to be considered (Schröder 2005; Schröder et al. 2005).
Two approaches are commonly used to study the fate of N fertilisers in soil and plant, namely the difference method and the 15N isotope technique. The difference method provides an estimate of fertiliser N uptake efficiency (FNUEdiff) by determining the extra amount of mineral N fertiliser or slurry N taken up compared to an unfertilised treatment, while assuming that the N transformation processes are not modified by the added N. The isotopic estimate (FNUE15N) is based on the recovery of labeled N in shoot organs. Both methods are subject to error sources, which are more pronounced on humous soils as (1) fertiliser application can induce priming effects (Bingemann et al. 1953) resulting in an overestimation of fertiliser N uptake efficiency estimated by the difference method, and (2) 15N fertiliser may be involved in mineralisation-immobilisation turnover (MIT) and biological interchange processes, leading to lower N recovery. Consequently, FNUE estimated by the difference method often exceeds that obtained by isotopic methods. Additional variation of published FNUE rates for mineral N and slurry is due to field site history, N management, and cropping system (Xue et al. 2005; Nissen and Wander 2003; Muñoz et al. 2004; Stevens et al. 2005; Cusick et al. 2006). FNUE15N values found for mineral N fertiliser range from 18 to 49% (Timmons and Cruse 1990; Khanif et al. 1984; Zhou et al. 2000; Stevens et al. 2005; Vesterager et al. 2008), while FNUEdiff vary between 22 and 94% (Ma et al. 1999; Schröder 1999; Stevens et al. 2005; Nissen and Wander 2003; Halvorson et al. 2005; Montemurro et al. 2006). Some studies even reported FNUEdiff values up to 145% (Maidl et al. 1999). Similar ranges were detected for slurry N with isotopic techniques between 10 and 50% (Muñoz et al. 2003; Chantigny et al. 2004; Muñoz et al. 2004), and FNUEdiff varying between 5 and 67% (Schröder 1999; Ma et al. 1999; Muñoz et al. 2003; Schröder et al. 2005), but also down to −16% (Maidl et al. 1999).
The objectives of this research were to
-
(1)
investigate if N rate and application mode influence the short-term FNUE calculated by the difference method and the 15N technique,
-
(2)
quantify the effects of N rate and of application mode, i.e. separate versus combined application of mineral fertiliser and cattle slurry, on short-term FNUE by silage maize, and trace the resulting N partitioning in the various plant parts,
-
(3)
study the contribution of the first and second N dressing of a split mineral N application to total N uptake, and
-
(4)
analyse the impact of N fertiliser type and level on maize N uptake from the upper soil layer
Materials and methods
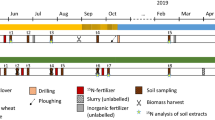
A field experiment was conducted within the framework of the ‘N project Karkendamm’ (Taube and Wachendorf 2000) at the research station Karkendamm (53°55′N, 9°55′E; 14 m a.s.l.) in northern Germany during 3 years (1998–2000). The soil was classified as sandy Aquod, with soil characteristics showing a large spatial variation as indicated by the organic carbon content of the plough layer varying between 4.2 and 7.7% and pH values between 5.2 and 5.7. The climate at the experimental site is moderately maritime, characterised by wet, cool summers and mild winters, with moderate seasonal temperature variation. Long-term mean temperature, annual precipitation and climatic water balance (Haude 1955; Wendling 1995) are 8.4°C, 824 and 312 mm, respectively. Average annual temperatures in the experimental years 1998–2000 were 8.6, 9.6, and 9.3°C, and precipitation amounted to 1,098, 705, and 635 mm. The experimental site had been cropped with forage maize since 1992 and had received 30 to 40 m³ slurry and 50 kg mineral N ha−1 year−1 each year.
The present study was embedded in a larger field trial, which was established as a two-factor split-plot design with four randomised blocks as replicates and a plot size of 255 m². Treatments were three cattle slurry fertilisation rates (0, 20, 40 m³ ha−1), factorially combined with four mineral N rates (0, 50, 100, 150 kg N ha−1 as calcium ammonium nitrate, CAN). The slurry was applied between spring ploughing and sowing and was immediately incorporated into the soil. Mineral N fertilisation was split into two equal applications at the one-leaf and six-leaf stages. All plots received a starter phosphorus fertilisation at sowing, where the fertiliser (superphosphate) was drilled 5 cm below and 5 cm to the side of the seed at a rate of 30 kg P2O5 ha−1. Additionally, 35 kg P2O5 ha−1 were applied at the 1-leaf stage. Potassium chloride was supplemented at sowing in order to adjust the K supply on all plots to the amount of the largest slurry application. An early maize hybrid (Naxos) was sown between late April and early May in rows that were 0.75 m apart, with a final plant density of 10–11 plants m−2. For more details, see Wachendorf et al. (2006a).
We selected seven of the twelve above-mentioned treatments to include them in the underlying study, namely all mineral-only treatments (0, 50, 100, 150 kg N ha−1), two slurry-only treatments (20, 40 m³ ha−1), and a combination of slurry and mineral fertiliser (40 m³ ha−1 plus 50 kg mineral N ha−1). Microplots were installed in each field replicate of the seven treatments, where 15N-labelled mineral fertiliser or 15N-labelled slurry was applied to analyse the N recovery of mineral N (experiment 1) and of slurry (experiment 2), and to study the N uptake from the upper soil layer (experiment 3); see Table 1. Locations of microplots within main plots were moved each year in order to exclude any residual effects resulting from 15N applications in the preceding year(s).
Experiment 1: mineral fertiliser N uptake efficiency
Estimation of the short-term mineral N uptake efficiency was based on the mineral-only treatments (0, 50, 100 and 150 kg N ha−1) and the combined mineral and slurry treatment (50 kg mineral N ha−1 plus 40 m³ ha−1); see Table 1. Rates of mineral fertiliser were given in two equal dressings on two separate microplots of 4.5 m² in size (three maize rows) at the one-leaf (mid-May) and eight-leaf stage (mid-June), where one microplot received labelled 15NH 154 NO3 (1998/1999: 1.43167 at%15N; 2000: 3.84145 at%15N) as a first dressing, and the other microplot as a second dressing. The remaining N amount was applied as unlabelled fertiliser. In the combined mineral and slurry treatment, the mineral part was labelled (15NH 154 NO3), while the slurry (72, 136, and 148 kg N ha−1 in 1998, 1999, and 2000, respectively) was unlabelled.
Experiment 2: slurry N uptake efficiency
Estimation of short-term slurry N uptake efficiency included the slurry-only (0, 40 m³ ha−1) and the combined mineral and slurry treatments (50 kg mineral N ha−1 plus 40 m³ ha−1); see Table 1. Slurry was labelled (0.72776 at%15N) by feeding 15N-enriched maize silage to steers and by collecting their excrements. Between ploughing and sowing, slurry was applied on microplots of 3.5 m² in size (three maize rows) with a watering can at a rate equivalent to 40 m³ ha−1 (1998: 72 kg N ha−1; 1999: 136 kg N ha−1; 2000: 148 kg N ha−1). Slurry was incorporated by hand tools immediately after application to reduce ammonia volatilisation. Mineral fertiliser (CAN) of the combined treatment was unlabelled.
Experiment 3: N uptake from upper soil layer
The experiment included four treatments: control (0 kg N ha−1), 20 m³ slurry (1998: 36 kg N ha−1, 1999: 68 kg N ha−1, 2000: 74 kg N ha−1), 40 m³ slurry (1998: 72 kg N ha−1; 1999: 136 kg N ha−1; 2000: 148 kg N ha−1), and 40 m³ slurry with an additional 50 kg mineral fertiliser N ha−1, where slurry and mineral N were unlabelled. To estimate the N uptake from the upper soil layer, 4.2 m²-sized microplots (three maize rows) were installed in each treatment, where K15NO3 (10.6 at%15 N; 4.1 kg N ha−1) was applied to the upper soil layer. To this end, the labelled N fertiliser was spread with a hand sprayer on 9 m³ before ploughing. Subsequently, additional 4 l of water per m² were applied with a watering can to facilitate infiltration. One to 3 days later, slurry was applied and incorporated, and afterwards maize was sown.
Crop sampling and processing
At silage maturity (5 Oct 1998, 22 Sept 1999, 29 Sept 2000), five labelled plants were cut at the soil surface from the middle row in each microplot and separated into leaves, stalks and ears. Main plots were harvested by a maize chopper. Samples of main plots and microplots were weighed and dried at 58°C to a constant weight. To obtain a more reliable quantification of dry matter yield for the microplots, the relative proportions of leaves, stalks, and ears of the labelled plants harvested in the microplot were multiplied with the dry matter yield (t DM ha−1) recorded in the respective main plot.
Representative subsamples were ground into 1-mm particles for near-infrared reflectance spectroscopy (NIRS) analysis and homogenised with a ball mill (Retsch MM2000, Haan, Germany) for 15N determination. The total N content of the main plots and plant fractions was estimated by NIRS. Ground samples were scanned on a NIRSystems 5000 scanning monochromator (FOSS GmbH, Rellingen, Germany), and software (ISI-version) for data collection and manipulation was supplied by Infrasoft International® (ISI, Port Matilda, PA, USA). Calibrations were developed separately for vegetative and reproductive plant fractions. Calibration and validation subset samples were analysed for N content using a C/N autoanalyser (Vario Max CN, Elementar Analysensysteme, Hanau, Germany). Standard errors of prediction ranged between 0.68 and 1.97 g N kg−1 DM, depending on the plant fraction (Volkers et al. 2003). Due to the experimental design (Table 1), some treatments were included in two (40 m³ ha−1 slurry) or three experiments (40 m³ ha−1 slurry plus 50 kg mineral N ha−1), but with different 15N labelling. In the analysis of the DM and N yield of whole plants and plant parts, averages were calculated for the different labellings. The 15N enrichment of plant parts was determined using a mass spectrometer (ThermoFinnigan Delta C, Bremen, Germany) coupled with an elemental analyser (Carlo Erba 1108, CE Instruments Milano/Italy; Conflo II).
Calculation of N recovery
Using the 15N isotope technique, the amount of N in a given plant part or the whole plant that derived from the labelled mineral fertiliser or slurry (Ndff, kg N ha−1) was estimated by the following equation:
where a, b, and c denote the atom% 15N concentrations in the fertiliser (mineral or slurry), unlabelled control, and labelled plant parts or whole plant, respectively, and p is the total N (kg N ha−1) in a given plant part or the whole plant.
Consequently, the N taken up by a plant part or the whole plant originating from non-fertiliser (non-slurry), Ndfs (kg N ha−1), was obtained by
Fertiliser N uptake efficiency (FNUE15N) was then calculated by
with f, the amount of labelled N fertiliser (kg N ha−1) applied. The difference method estimates the fertiliser N uptake efficiency (FNUEdiff) by determining the extra amount of mineral N fertiliser or slurry N taken up compared to an unfertilised treatment, and by assuming that N transformation processes are not modified by the added N.
where d is the nitrogen yield (kg N ha−1) of the fertilised treatment, e is the nitrogen yield (kg N ha−1) of the unfertilised reference treatment, and g denotes the total amount of N fertiliser applied (kg N ha−1). For mineral- or slurry-only treatments, we used the control (0 kg N ha−1) as a reference (e), while S0M50 was used for treatment S40M50.
Statistical analysis
The field trial was conducted over 3 years, where treatments were allocated to fixed plots. It might be argued that this requires the introduction of year as fixed or random effect in the statistical model. Since, however, year had no strong impact on silage maize yield and N recovery, analysis of variance was conducted without any year effects. The effect of N fertiliser amount and fertiliser type was tested for significance using Proc Mixed of SAS®9.0 (SAS Institute Inc 2001). Statistical analysis was conducted separately for each of the three experiments and each plant fraction. Data presented are LSMeans across years produced in the analysis. Significance was declared at P < 0.05, and multiple comparisons were conducted using the Tukey-test.
Results
Dry matter and nitrogen yield
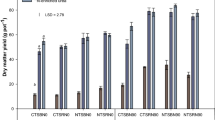
Shoot dry matter yield ranged between 7.1 and 14.3 t DM ha−1, with the lowest significant yield found in the control and highest yield obtained in the mineral-only treatment S0M150 (Table 2). Due to the large contribution of the ear to the shoot yield, an identical ranking was found among treatments for the ear DM yield, with values varying between 3.5 and 8.4 t DM ha−1. Also for the stalk DM yield, the treatments showed a similar order, while for the leaf DM yield, less significances were detected. The treatment ranking for N yield corresponded to the observed DM yield. Consistently, the lowest N yield was recorded for the control, while S0M150 achieved the highest N yield for the shoot as well as all plant parts considered. However, differences in N yield among treatments were more pronounced than differences in DM yield, e.g. stalk N yield of the control was only 32% of the maximum N yield achieved in treatment S0M150, while for the DM yield the control still achieved 66% of the S0M150 yield. Notably, the application of 50 kg mineral N ha−1 in addition to 40 m³ of slurry (1998: 72 kg N ha−1; 1999: 136 kg N ha−1; 2000: 148 kg N ha−1) did not increase the DM yield, but resulted in a significantly higher N yield of the shoot, ear, and leaf.
Recovery of mineral fertiliser N
Recovery of labelled N in the aboveground plant parts revealed that the main N sink at silage maturity was the ear. Each of the two mineral N dressings contributed about 7.0 to 30.3 kg N ha−1 to the overall N uptake of the ear (Fig. 1a). Stalks and leaves were minor N sinks, with 15N recovery from mineral fertiliser varying between 1.7 and 6.6 kg N ha−1 and 3.3 and 10.7 kg N ha−1 in each dressing, respectively (Fig. 1b, c).

Effect of mineral N application rate on the 15N recovery (kg N ha−1) of a ears, b stalks, c leaves, and d whole plant in the first and second fertiliser dressing (Ndff), and N amount derived from non-fertiliser N (Ndfs). Values represent means of 3 years and 4 replicates
For leaves, 15N recovery from the first dressing in mineral fertiliser-only treatments was lowest in S0M50 (3.3 kg N ha−1), but similar in S0M100 and S0M150, with 8.0 and 10.7 kg N ha−1. N recovery from the second dressing ranged between 3.8 and 9.6 kg N ha−1. For a given N rate, fertiliser N uptake derived from the first and second dressings did not differ significantly. In stalks, N recovered from the first dressing (1.7–6.5 kg N ha−1) revealed a significant increase with N fertilisation rate (Fig. 1b). N uptake from the second dressing ranged between 2.1 and 6.6 kg N ha−1 and was highest in S0M150, but similar in all other mineral fertiliser treatments. As with the leaves, the amounts of N recovered from the first and second dressings were similar within a given treatment. Divergent effects were observed for the ear (Fig. 1a). Differences in 15N recovery between both dressings were detected only for the S0M100 treatment, where the ear contained more fertiliser N from the first than the second dressing. An increasing N rate resulted in an increased N recovery of the first N dressing for the mineral-only treatments. Nitrogen originating from the second dressing was highest in S0M150, but similar in the other treatments, amounting to 9.3, 9.5 and 11.1 kg N ha−1 for S0M50, S40M50 and S0M100, respectively. On a whole plant basis, N recovered from the first dressing amounted to 12.0, 13.7, 29.6 and 43.4 kg N ha−1 for S0M50, S40M50, S0M100 and S0M150, respectively (Figs. 1d, 2), and differed between mineral-only treatments, while additional slurry application (S0M50 vs. S40M50) had no impact. Recoveries of mineral fertiliser N in the second dressing were 15.2, 16.3, 21.3 and 46.5 kg N ha−1 for S0M50, S40M50, S0M100 and S0M150, respectively, with S0M150 having a significantly higher 15N recovery than the other treatments. For a given fertiliser treatment, N recovery tended towards higher values in the second compared to the first dressing (except for S0M100), but no significant differences became evident. Whole plant N uptake originating from non-fertiliser N (Ndfs) increased with N rate up to treatment S0M100, while further N input showed no further increase or even caused a slight decrease. Consistently, a similar trend was observed for all plant parts.

Whole plant N recovery (kg N ha−1) from first and second dressing of mineral N, slurry, the upper soil layer, and non-fertiliser N as influenced by fertiliser treatment, with ctrl: control, S20M0: slurry only (20 m³ ha−1), S40M0: slurry only (40 m³ ha−1), S40M50: combined slurry (40 m³ ha−1) and mineral N (50 kg N ha−1). In treatment S20M0 only N applied to upper soil layer was labelled. Values represent means of 3 years and 4 replicates
Fertiliser N uptake efficiency (FNUE15N) estimated by the isotope technique for the whole plant were 54, 51, 60 and 61% for treatments S0M50, S0M100, S0M150 and S40M50, respectively, and did not differ significantly (Fig. 3a). Corresponding values estimated by the difference method (FNUEdiff) were considerably higher, except for treatment S40M50, amounting to 98, 86, 81, and 54%. The FNUEdiff of mineral-only treatments did not differ substantially, and additional slurry application (S0M50 vs. S40M0) caused a significant decrease of FNUEdiff.

Impact of N fertiliser treatment on whole plant fertiliser N uptake efficiency estimated by the isotopic technique (FNUE15N, %; black bars) and by the difference method (FNUEdiff, %; grey bars) for (A) mineral fertiliser N and (B) slurry N. Values represent means of 3 years and 4 replicates. Error bars indicate standard errors of the mean values
Recovery of slurry N and of mineral N from the upper soil layer
On the whole plant basis, nitrogen originating from slurry application was similar in the S40M0 and S40M50 treatments, totalling to 21.8 and 22.2 kg N ha−1 (Fig. 2). While 61% of the slurry N was found in ears, leaves and stalk contributed 27 and 13%, respectively (data not presented), indicating that the sink-source relationship for slurry N followed the same pattern as for mineral fertiliser N. Additional slurry application (S40M0 vs. S40M50) caused significantly higher N recovery in stalks, but not in leaves and ears (data not shown).
Uptake of labelled N from the upper soil layer (0–10 cm) was significantly higher in the S40M0 (9.3 kg N ha−1) and S40M50 (10.1 kg N ha−1) treatments than in the control (3.0 kg N ha−1) and the S20M0 treatment (6.4 kg N ha−1); see Fig. 2. In accordance with slurry N recovery, less labelled soil N was found in leaves and stalks than ears (data not presented). Uptake of non-fertiliser N from layers below 10 cm was considerably lower in the control (43.9 kg N ha−1) compared to the S40M0 (80.8 kg N ha−1) and S40M50 (77.3 kg N ha−1) treatments.
Slurry N uptake efficiency (FNUE15N) in the whole plant obtained by the isotope technique revealed a similar level for treatments S40M0 (22%) and S40M50 (21%); see Fig. 3b. In contrast, the difference method (FNUEdiff) resulted in a significantly higher slurry N uptake efficiency for S40M0 (62%) than for S40M50 (39%).
Discussion
Mineral N fertiliser use efficiency
Fertiliser N uptake efficiency of mineral fertiliser varied between 51 and 61% at the whole crop level, which is considerably higher than reported previously for sandy soils (Khanif et al. 1984). In our study, however, the N rate was split to better match the N demand of the crop (López-Bellido et al. 2006). It may therefore be assumed that less fertiliser N was subject to N losses caused by nitrate leaching and nitrous oxide emission during the vegetation period (Torbert et al. 1993; Maidl 1990). The split N application tended to result in higher N recoveries for the second dressing, except for the S0M100 treatment. This finding deviates from several other works, which documented a clear effect of fertiliser N timing on 15N recovery, with substantially higher N use efficiency for N applied sidedressed than at planting (Bigeriego et al. 1979; Russelle et al. 1981; Jokela and Randall 1997; Seo et al. 2006). A high proportion of the nitrogen taken up from the first dressing was found in the ear while recovery in leaves and stalks was low, for instance 60, 25, and 14%, respectively, in the S0M150 treatment. Nitrogen applied in early growth stages and taken up by vegetative plant parts is remobilised during reproductive growth and translocated to the grains. The remobilisation pattern, however, varies among plant parts, with mobilisation of N in stalks starting earlier than in leaves (Weiland and Ta 1992). The later N is taken up during vegetative growth, the more likely it is mobilised to the grains, while N absorbed during grain filling is mainly allocated directly to the grains (Subedi and Ma 2005). Therefore, a higher N recovery of the second dressing compared to the first would have been expected for the ear. In our study, however, differences in N recovery between the first and second dressing were not pronounced, and treatment S0M100 even showed a reverse effect. This may partly be due to the relatively short time interval of 3–4 weeks between the two N dressings.
An increasing N rate neither caused a consistent trend in FNUE15N for single plant parts nor at the whole plant level, which is contrary to previous studies (Bigeriego et al. 1979; Reddy and Reddy 1993; Jokela and Randall 1997; Nissen and Wander 2003). Moreover, we did not find any effect of N rate on 15N distribution among plant parts, while Subedi and Ma (2005) reported the proportion of 15N reaching the grain to be inversely related to N supply. One possible explanation for the deviating results in our study may be that mineral N was not provided in excess, as indicated by the linear increase of total N yields in the mineral-only treatments (Table 2). Support for our findings comes from a meta-analysis of N fate in grain cropping systems conducted by Gardner and Drinkwater (2009). The authors concluded that a reduced N rate had no impact on 15N recovery by the crop, but a positive effect on total 15N recovery of plant and soil, and that the corresponding N rate was an important predictor of unrecovered 15N.
FNUEdiff of mineral fertiliser obtained in the present study (81–98%) was in the range reported by Varvel and Peterson (1990), but higher compared to observations by Schröder (1999). This is probably mainly attributed to differences in the yielding performance of the control treatment caused by the manuring history of the site (Harmsen 2003a, b), but FNUEdiff can also be affected by environmental conditions and the maize genotype under evaluation (Coque et al. 2006). Currently available results on the effect of mineral N fertilisation on FNUEdiff are ambiguous (Van Dijk and Brouwer 1998; Ma et al. 1999; Nissen and Wander 2003; Halvorson et al. 2005; Stevens et al. 2005; Montemurro et al. 2006). Our own findings strongly support decreasing FNUEdiff with increasing N supply. In low N treatments, 15N data revealed a considerable additional non-fertiliser N uptake, which is ascribed to mineral fertiliser when calculating N recovery by FNUEdiff. In highly fertilised plots, however, the additional N uptake of plants mainly derived from fertiliser N (Fig. 2), leading to a minor discrepancy between both approaches. This may also explain why additional slurry application (S0M50 vs. S40M50) caused a substantial decline of FNUEdiff while FNUE15N was nearly constant. Contrary to our hypothesis, FNUE15N data reveal that slurry application obviously did not exert any direct effect on mineral N recovery, which may be due to a sizeable interval between slurry application and mineral N dressings.
The phenomenon of increased soil N uptake in fertilised plots is mainly restricted to N-deficient sites with high soil organic matter content (Harmsen 2003a). In our study, an apparent added nitrogen interaction (ANI) (Jenkinson et al. 1985) may have occurred. Considering the different causes of an apparent ANI (Kuzyakov et al. 2000; Blagodatskaya and Kuzyakov 2008), displacement effects and MIT (mineralisation-immobilisation and turnover) processes seem likely. Furthermore, a stronger and deeper root system in the fertilised treatments may have resulted in additional N uptake.
These effects only apply to N recovery estimation by FNUE15N, and FNUEdiff thus seems to be a more appropriate tool. When evaluating both approaches, one should, however, keep in mind that different N pools are considered. While FNUE15N detects recently applied labelled N, the difference method includes recently applied N as well as N given in the past, which entered the soil N pool and has been released in exchange with recently applied fertiliser N. The amount of N released from the soil N pool can be considerable, as indicated by significant N leaching losses despite negative field N balances (Wachendorf et al. 2006a, b). The fact that N released from the soil N pool is included in FNUEdiff estimation limits its suitability for long-term experiments, where control plots show severe N deficiency and may cause an overestimation of N recovery by FNUEdiff (Schindler and Knighton 1999; Stevens et al. 2005) as in, for instance, long-term monoculture cultivation.
Slurry N use efficiency
As expected, for both investigated approaches (FNUE15N, FNUEdiff) fertiliser N uptake efficiency of slurry was lower compared to mineral fertiliser, mainly due to the organically bound N fraction, which makes up about 45% of total nitrogen and is little available in the year of application. Taking the proportion of organically bound N into consideration, the difference in FNUE between slurry and mineral fertiliser will diminish, e.g. to 38–40% FNUE15N for slurry versus 51–61% FNUE15N for mineral N. A further cause might be due to gaseous N losses after slurry spreading. In our study, however, slurry was immediately incorporated after application, so that we may assume ammonia losses to be negligible.
FNUE15N was not affected by additional mineral fertiliser application and varied from 21 to 22% (Fig. 2). Recovery rates reported by Muñoz et al. (2003) for dairy manure in the year of application were somewhat lower (17%), possibly because dairy excreta were mixed with straw, leading to an increased C/N ratio. As is well-known, the C/N ratio is an important determinant of short-term plant availability (Sørensen et al. 2003; Chantigny et al. 2004; Powell et al. 2005). Non-fertiliser N uptake increased by more than 40 kg N ha−1 in slurry treatments compared to the control (Fig. 2), yet the uptake of non-labelled N was always lower than unaccounted slurry N. As for mineral N-only treatments, a likely explanation for that finding is an apparent added nitrogen interaction (ANI). Support comes from previous studies (Sørensen and Jensen 1995; Chadwick et al. 2000) documenting that slurry N underlies immobilisation processes and is released in the long term. Our soil labelling data, however, indicate that a real positive ANI might have occurred. With increasing slurry application (control, S20M0, S40M0), the N uptake from the upper soil layer increased linearly by a factor of three (r² = 0.99). A real ANI would not exclude an apparent ANI since both processes can take place simultaneously.
Slurry FNUEdiff of the present study was in the range found in previous studies under similar soil and climatic conditions (Schröder et al. 1993; Schröder et al. 2005), but exceeded the FNUEdiff values reported by Schröder (1999). The low FNUEdiff in the latter study is somewhat surprising since the field trials were also conducted on sandy soil and since the slurry was banded or incorporated into the soil. Yet, soil N status was higher compared to our study due to regular, intensive slurry application, leading to high-yielding control plots, reflecting the importance of manuring history to FNUEdiff estimation (Mallory and Griffin 2007).
In accordance with mineral N recovery estimation, slurry FNUEdiff resulted in substantially higher recovery rates than slurry FNUE15N, which has also been reported elsewhere (Muñoz et al. 2003). The difference between FNUEdiff and FNUE15N was not constant among slurry treatments (Fig. 3b), since different reference treatments were used for estimating FNUEdiff, i.e. control for S40M0 and S0M50 for S40M50. According to the 15N data, the additional uptake of unlabelled N in S40M0 compared to the reference (control) was higher (43.1 kg N ha−1) than for S40M50 (18.7 kg N ha−1) when compared to its respective reference (S0M50). This led to an underestimation of slurry N recovery for treatment S40M50 and corroborates the assumption that constant plant N uptake in reference and fertilised treatments is one of the major error sources in FNUEdiff estimation (Rao et al. 1992). Likewise, the recovered mineral N for treatment S40M50 (27.8 kg N ha−1) that was obtained by the difference method is biased since S40M0 served as a reference. When assuming a constant soil N uptake in all treatments, the sum of soil N and recovered fertiliser N in S40M50 (43.8 kg slurry N, 27.8 kg mineral fertiliser N) was 21.1 kg N ha−1 lower than the actual measured N yield (Table 2). In conclusion, the present study gives strong evidence that the difference method does not allow an accurate estimation of N recovery when different N sources are applied together.
Conclusions
The present study substantiated the fact that the approaches used for estimating fertiliser N uptake efficiency, i.e. stable isotopic technique vs. difference method, consider different N pools, differ in magnitude, and have inherent estimation errors. The recovery rates obtained by FNUE15N, for instance, exceeded FNUEdiff values by a factor of 1.45 in the case of mineral N and by 2.42 in the case of slurry N. It could clearly be demonstrated that the approaches under evaluation come to different conclusions when applying mineral and organic N fertilisers separately compared to a combined application, and correspondingly their suitability depends on the question of interest. While N recovery by FNUE15N was not influenced by additional N sources, be it mineral or organic, FNUEdiff resulted in substantially lower values for the combined application. Similarly, an increasing N rate did not influence FNUE15N, but led to a considerable decline in FNUEdiff. We therefore regard FNUEdiff as an inappropriate tool for estimating N recovery of a single N source when applied in combination with other N fertiliser types. Recovery rates obtained by FNUEdiff should be treated with caution. Future challenges will include the simultaneous labelling of soil N and fertiliser N fractions to facilitate evaluation of their contribution to yield formation and N transformation processes in the soil.
References
Bigeriego M, Hauck RD, Olson RA (1979) Uptake, translocation and utilization of 15N-depleted fertilizer in irrigated corn. Soil Sci Soc Am J 43:528–533
Bingemann CW, Varner JE, Martin WP (1953) The effect of the addition of organic materials on the decomposition of an organic soil. Soil Sci Soc Amer Proc 17:34–38
Blagodatskaya EV, Kuzyakov Y (2008) Mechanisms of real and apparent priming effects and their interdependence on soil microbial biomass and community structure: critical review. Biol Fertil Soils 45:115–131
Chadwick D, John F, Pain BF, Chambers BJ, Williams J (2000) Plant uptake of nitrogen from the organic nitrogen fraction of animal manures: a laboratory experiment. J Agric Sci 134:159–168
Chantigny MH, Angers DA, Morvan T, Pomar C (2004) Dynamics of pig slurry nitrogen in soil and plant as determined with 15N. Soil Sci Soc Am J 68:637–643
Coque M, Bertin P, Hirel B, Gallai A (2006) Genetic variation and QTLs for 15 N natural abundance in a set of maize recombinant inbred lines. Field Crops Res 97:310–321
Cusick PR, Kelling KA, Powell JM, Muñoz GR (2006) Estimates of residual dairy manure nitrogen availability using various techniques. J Environ Qual 35:2170–2177
Gardner JB, Drinkwater LE (2009) The fate of nitrogen in grain cropping systems: a meta-analysis of 15N field experiments. Ecol Appl 19:2167–2184
Haas G, Bach M, Zerger C (2005) Landwirtschaftsbürtige Stickstoff- und Phosphor-Bilanzsalden. LÖBF-Mitteilungen, February
Halvorson AD, Schweissing FC, Bartolo ME, Reule CA (2005) Corn response to nitrogen fertilization in a soil with high residual nitrogen. Agron J 97:1222–1229
Harmsen K (2003a) A comparison of the isotope-diution and the difference method for estimating fertilizer nitrogen recovery fractions in crops. I. Plant uptake and loss of nitrogen. Neth J Agr Sci 50:321–347
Harmsen K (2003b) A comparison of the isotope-diution and the difference method for estimating fertilizer nitrogen recovery fractions in crops. II. Mineralization and immobilization of nitrogen. Neth J Agr Sci 50:349–381
Haude W (1955) A basic approach to assess the transpiration of plants (in German). Mitt Dt Wetterdienst 11:1–24
Herrmann A, Taube F (2005) Nitrogen concentration at maturity—an indicator of nitrogen status in forage maize. Agron J 97:201–210
Jenkinson DS, Fox RH, Rayer JH (1985) Interactions between fertilizer nitrogen and soil nitrogen—the so-called ‘priming’ effect. J Soil Sci 36:425–444
Jokela WE, Randall GW (1997) Fate of fertilizer nitrogen as affected by time and rate of application on corn. Soil Sci Soc Am J 61:1695–1703
Khanif YM, van Cleemput O, Baert L (1984) Field study of the fate of labelled fertilizer nitrate applied to barley and maize in sandy soils. Fert Res 5:289–294
Kuzyakov Y, Frieder JK, Stahr K (2000) Review of mechanisms and quantification of priming effects. Soil Biol Biochem 32:1485–1498
López-Bellido L, López-Bellido RJ, López-Bellido FJ (2006) Fertilizer nitrogen efficiency in durum wheat under rainfed mediterranean conditions: effect of split application. Agron J 98:55–62
Ma BL, Dwyer LM, Gregorich EG (1999) Soil nitrogen amendment effects on nitrogen uptake and grain yield of maize. Agron J 91:650–656
Maidl F-X (1990) Pflanzenbauliche Aspekte einer gezielten N-Versorgung und verbesserten N-Ausnutzung. Landwirtschaftl Jahrbuch 67. Sonderheft 2:71–87
Maidl F-X, Sticksel E, Valta R (1999) Untersuchungen zur verbesserten Gülleverwertung im Maisanbau. 1. Report: utilization of nitrogen in slurry by maize (silage and grain) using different application techniques. Pflanzenbauwiss 3:9–16
Mallory EB, Griffin TS (2007) Impacts of soil amendment history on nitrogen availability from manure and fertilizer. Soil Sci Soc Am J 71:964–973
Montemurro F, Maiorana M, Ferri D, Convertini G (2006) Nitrogen indicators, uptake and utilization efficiency in a maize and barley rotation cropped at different levels and sources of N fertilization. Field Crops Res 99:114–124
Muñoz GR, Powell JM, Kelling KA (2003) Nitrogen budget and soil N dynamics after multiple applications of unlabeled or 15N-enriched dairy manure. Soil Sci Soc Am J 67:817–825
Muñoz GR, Kelling KA, Powell JM, Speth PE (2004) Comparison of estimates of first-year dairy manure nitrogen availability or recovery using nitrogen-15 and other techniques. J Environ Qual 33:719–727
Neeteson JJ (2000) Nitrogen and phosphorus management on Dutch dairy farms: legislation and strategies employed to meet the regulations. Biol Fertil Soils 30:566–572
Nissen TM, Wander MM (2003) Management and soil-quality effects on fertilizer-use efficiency and leaching. Soil Sci Soc Am J 67:1524–1532
O’Mara FP, Fitzgerald JJ, Murphy JJ, Rath M (1998) The effect on milk production of replacing grass silage with maize silage in the diet of dairy cows. Livestock Prod Sci 55:79–87
Powell JM, Kelling KA, Muñoz GR, Cusick PR (2005) Evaluation of dairy manure nitrogen-15 enrichment methods on short-term crop and soil nitrogen budgets. Agron J 97:333–337
Rao ACS, Smith JL, Parr JF, Papendick RI (1992) Considerations in estimating nitrogen recovery efficiency by the difference and isotopic dilution methods. Fert Res 33:209–217
Reddy GB, Reddy KR (1993) Fate of nitrogen-15 enriched ammonium nitrate applied to corn. Soil Sci Soc Am J 57:111–115
Russelle MP, Deibert EJ, Hauck RD, Stevanovic M, Olson RA (1981) Effects of water and nitrogen management on yield and 15N-depleted fertilizer use efficiency of irrigated corn. Soil Sci Soc Am J 45:553–558
Samborski SM, Tremblay N, Fallon E (2009) Strategies to make use of plant sensors-based diagnostic information for nitrogen recommendations. Agron J 101:800–816
SAS Institute Inc (2001) SAS Software Release 8.2. SAS Institute Inc, Cary
Schindler FV, Knighton RE (1999) Fate of fertilizer nitrogen applied to corn as estimated by the isotopic and difference methods. Soil Sci Soc Am J 63:1734–1740
Schmitt MR, Edwards GE (1981) Photosynthetic capacity and nitrogen use efficiency of maize, wheat, and rice: a comparison between C3 and C4 photosynthesis. J Exp Bot 32:459–466
Schröder JJ (1999) Effect of split applications of cattle slurry and mineral fertilizer-N on the yield of silage maize in a slurry-based cropping system. Nutr Cycl Agroecosyst 53:209–218
Schröder JJ (2005) Revisiting the agronomic benefits of manure: a correct assessment and exploitation of its fertilizer value spares the environment. Bioresour Technol 96:253–261
Schröder JJ, ten Holte L, van Keulen H, Steenvoorden JHAM (1993) Effects of nitrification inhibitors and time and rate of slurry and fertilizer N application on silage maize yield and losses to the environment. Fert Res 34:267–277
Schröder JJ, Neeteson JJ, Oenema O, Struik PC (2000) Does the crop or the soil indicate how to save nitrogen in maize production? Field Crops Res 66:151–164
Schröder JJ, Jansen AG, Hilhorst GJ (2005) Long-term nitrogen supply from cattle slurry. Soil Use Manag 21:196–204
Seo J-H, Meisinger JJ, Lee H-J (2006) Recovery of nitrogen-15-labeled hairy vetch and fertilizer applied to corn. Agron J 98:245–254
Sørensen P, Jensen ES (1995) Mineralization-immobilization and plant uptake of nitrogen as influenced by the spatial distribution of cattle slurry in soils of different texture. Plant Soil 173:283–291
Sørensen P, Weisbjerg MR, Lund P (2003) Dietary effects on the composition and plant utilization of nitrogen in dairy cattle manure. J Agric Sci 141:79–91
Stevens WB, Hoeft RG, Mulvaney RL (2005) Fate of nitrogen-15 in a long-term nitrogen rate study: II. Nitrogen uptake efficiency. Agron J 97:1046–1053
Subedi KD, Ma BL (2005) Effects of N-deficiency and timing of N supply on the recovery and distribution of labeled 15N in contrasting maize hybrids. Plant Soil 273:189–202
Taube F, Wachendorf M (2000) The Karkendamm project: a system approach to optimize nitrogen use efficiency on the dairy farm. Grassland Sci Eur 5:449–451
Timmons DR, Cruse RM (1990) Effect of fertilization method and tillage on nitrogen-15 recovery by corn. Agron J 82:777–784
Torbert HA, Hoeft RG, Vanden Heuvel RM, Mulvaney RL, Hollinger SE (1993) Short term excess water impact on corn yield and nitrogen recovery. J Prod Agric 6:337–344
Van Dijk W, Brouwer G (1998) Nitrogen recovery and dry matter production of silage maize (Zea mays L.) as affected by surface band application of mineral nitrogen fertilizer. Neth J Agric Sci 46:139–155
Varvel GE, Peterson TA (1990) Nitrogen fertilizer recovery by corn in monoculture and rotation systems. Agron J 82:935–938
Vesterager JM, Nielsen NE, Hogh-Jensen H (2008) Effects of cropping history and phosphorus source on yield and nitrogen fixation in sole and intercropped cowpea-maize systems. Nutr Cycl Agroecosyst 80:61–73
Volkers KC, Wachendorf M, Loges R, Jovanovic NJ, Taube F (2003) Prediction of the quality of forage maize by near-infrared reflectance spectroscopy. Anim Feed Sci Technol 109:183–194
Wachendorf M, Büchter M, Volkers K, Bobe J, Rave G, Loges R, Taube F (2006a) Performance and environmental effects of forage production on sandy soils. V. Impact of grass understorey, slurry application and mineral N fertilizer on nitrate leaching under maize for silage. Grass Forage Sci 61:243–252
Wachendorf M, Volkers KC, Loges R, Rave G, Taube F (2006b) Performance and environmental effects of forage production on sandy soils. IV. Impact of slurry application, minerall fertilizer and grass understorey on yield and nitrogen surplus of maize for silage. Grass Forage Sci 61:232–242
Weiland RT, Ta CT (1992) Allocation and retranslocation of 15N by maize (Zea mays L.) hybrids under field conditions of low and high N fertility. Aust J Plant Physiol 19:77–88
Wendling U (1995) Estimation of the transpiration by grass with the FAO model by Penman-Monteith (in German). Wasserwirtschaft 85:602–604
Xue JM, Sands R, Clinton PW, Payn TW, Skinner MF (2005) Priming effect of biuret addition on native soil N mineralisation under laboratory conditions. Soil Biol Biochem 37:1959–1961
Zhou X, Madramootoo CA, MacKenzie AF, Kaluli JW, Smith DL (2000) Corn yield and fertilizer N recovery in watertable-controlled corn-ryegrass systems. Eur J Agron 12:83–92
Acknowledgments
We are indebted to N. Jovanovic and K. Volkers for data sampling and gratefully acknowledge the excellent technical assistance of B. Biegler. We also thank G. Rave for support in the data analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nannen, D.U., Herrmann, A., Loges, R. et al. Recovery of mineral fertiliser N and slurry N in continuous silage maize using the 15N and difference methods. Nutr Cycl Agroecosyst 89, 269–280 (2011). https://doi.org/10.1007/s10705-010-9392-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-010-9392-2