
Abstract
Farm product diversification, shade provision and low access to fertilizers often result in the purposeful integration of upper canopy trees in cocoa (Theobroma cacao) plantations. Subsequent modification to light and soil conditions presumably affects nutrient availability and cocoa tree nutrition. However, the level of complementarity between species requires investigation to minimize interspecific competition and improve resource availability. We hypothesized beneficial effects of upper canopy trees on cocoa biomass, light regulation, soil fertility and nutrient uptake. We measured cocoa standing biomass and soil nutrient stocks under no shade (monoculture) and under three structurally and functionally distinct shade trees: Albizia zygia (D.C.) Macbr, a nitrogen fixer; Milicia excelsa (Welw.), a native timber species; and Newbouldia laevis (Seem.), a native small stature species. Vector analysis was employed to diagnosis tree nutrition. Cocoa biomass was higher under shade (22.8 for sole cocoa versus 41.1 Mg ha−1 for cocoa under Milicia), and declined along a spatial gradient from the shade tree (P < 0.05). Percent canopy openness differed between the three shade species (P = 0.0136), although light infiltration was within the optimal range for cocoa production under all three species. Soil exchangeable K was increased under Newbouldia, while available P decreased and total N status was unaffected under all shade treatments. Nutrient uptake by cocoa increased under shade (43–80% and 22–45% for N and P, respectively), with K (96–140%) as the most responsive nutrient in these multistrata systems. Addition of low-density shade trees positively affected cocoa biomass close to the shade tree, however proper management of upper stratum trees is required for optimum cocoa productivity and sustainability.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Multistrata planting schemes integrating both upper canopy trees and cocoa (Theobroma cacao L.) are purposefully developed by farmers to diversify farm products, secure land tenure, provide shade for cocoa seedlings and increase biophysical advantages for production. Specifically in regions with low access to inorganic fertilizers, multistrata plantations are established to maintain soil fertility and subsequently increase nutrient availability for cocoa. Presumably, enhanced production in these systems result from improved resource status associated with common agroforestry processes; efficient nutrient cycling, improved soil characteristics, modified light infiltration, enhanced moisture availability and reduced weed competition (Ahenkorah et al. 1987; Beer et al. 1998; Schroth et al. 2001; Hartemink 2005). Often, multiple species in an agroforestry system contribute functions not found in monocultures, such as the partitioning of resources, synchrony of resource use, and the ability of each species to capture and cycle nutrients (Ewel and Bigelow 1996; Schroth et al. 2001). However, facilitative interactions that enhance cocoa productivity may be offset by interspecific competition for particular resources. Presumably, both antagonistic and facilitative interactions may occur between species in a multistrata system. Therefore, knowledge of species complementarity in multistrata systems for maximizing resource availability is important for increasing crop productivity.
To date, several studies have focused on species interactions and subsequent growth in both natural communities and agroecosystems (Alpizar et al. 1986; Fassbender et al. 1991; Ewel and Bigelow 1996; Tilman 1999), where productivity is often found to be dependent on a functional attribute of a species (Silver et al. 1996; Hooper 1998) such as nitrogen fixation, or structural aspects such as canopy architecture and subsequent light availability (Ewel and Bigelow 1996). For cocoa plantation systems, it is hypothesized that once nutrient requirements are met, understory crop production is primarily dependent on the accessibility of solar radiation (Cunningham and Arnold 1962). The effect of upper canopy shade species on light infiltration is presumably dependent on species specific crown structure and subsequent modifications to light availability for the cocoa crop.
Although some work has been conducted in cocoa-shade tree systems (Alpizar et al. 1986; Ahenkorah et al. 1987; Fassbender et al. 1991), little research has examined species-specific modifications to light availability and soil nutrient pools as well as indirect effects on cocoa nutrient uptake. As cocoa nutrition is a critical factor for farm sustainability, we employed vector analysis, a comprehensive diagnostic tool to assess tree nutrient responses (Hasse and Rose 1995; Imo and Timmer 1997), to speculate on long-term maintenance of cocoa nutrition.
Our approach was to examine the effects of three structurally and functionally distinct upper canopy trees in farmer-designed multistrata agroforestry systems in Ghana. The upper canopy species were: Albizia zygia (D.C.) Macbr, a nitrogen fixer; Milicia excelsa (Welw.), a native timber species; and Newbouldia laevis (Seem.), a native small stature multi-purpose species. The objectives of our study were to 1) compare cocoa tree biomass under no shade and under three different shade species, 2) measure light availability, soil nutrient stocks and cocoa nutrient content under no shade and three shade trees, and 3) employ vector analysis to diagnosis nutrient uptake and relate cocoa biomass and nutrient status with possible causal factors.
Materials and methods
Study site and experimental design
Field work, conducted over one month during the pre-rainy season (2005) with additional data collection during the pre-rainy season in 2006, was in the moist semi-deciduous tropical zone in the Western Region (06°12′ N and 02°29′ W) of Ghana, West Africa. The soils of the study region are dominated by ochrosol-oxisol intergrades (Rhodic Ferralsol) that have developed from granite parent material. These highly leached soils are relatively low in fertility and highly acidic (Soils Survey Division 1969). This region typically has a mean annual precipitation of 1100 mm in bi-modal rainy seasons (April to July and September to November) with an average aboveground temperature of 26.0 °C. The study site, an 8 year-old cocoa (or cacao) plantation (approximately 2 ha in size), was selected due to its history of sole landownership of cocoa production since forest conversion, consisting of bi-monthly weeding without burning and fertilizer use. Stand structure was comprised of two strata: a cocoa stratum at a height of approximately 5 m and a shade stratum as described in Table 1. Although these two strata were found throughout most of the plantation, some areas contained only monoculture cocoa. The three shade species were replanted or naturally regenerated at the time of conversion from secondary forest to cocoa plantation. The plantation had a uniform cocoa planting density of approximately 1100 trees ha−1 interspersed with sparsely distributed upper canopy test species at a density of approximately 30 trees ha−1.
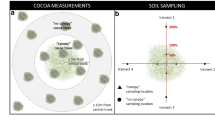
The on-farm experimental design, a randomized complete block design, consisted of three multistrata treatments: cocoa with Albizia zygia (Albizia + cocoa), cocoa with Milicia excelsa (Milicia + cocoa) and cocoa with Newbouldia laevis (Newbouldia + cocoa), and a monoculture control (cocoa). Each multistrata treatment consisted of one shade tree in combination with a cocoa tree stratum. By on-site inspection, 12 random 10 m radius circular plots centered around the upper canopy shade tree were selected with approximately 25 cocoa trees in each plot. Plots were then blocked into three, with each block consisting of the three multistrata treatments and a control as shown in Fig. 1. Shade trees on site were established, either by natural regeneration or planting, during cocoa tree establishment, therefore are the same ages as the cocoa trees. It was confirmed in advance by the landowner that shade tree location, hence treatment application, was on a random basis. Two sampling distances representing a radial spatial gradient were selected 1) within 5 m from the shade tree (<5 m) and 2) 5 to 10 m away from the shade tree (5–10 m). Although the latter is further away from the shade tree, this is not to be confused with sole cocoa (monoculture control), where no shade trees were located. The 5–10 m radial distance is simply a greater distance from the shade tree.

Schematic of on-farm experimental design. Twelve random 10 m radius circular plots centered on the upper canopy shade tree were selected by on-site inspection. Plots were then blocked into three, each block consisting of the three multistrata treatments [Albizia + cocoa (A), Milicia + cocoa (M) and Newbouldia + cocoa (N)] as well as a randomly selected area of monoculture cocoa (C). Biomass and soil sampling was conducted within two radial distances (<5 m and 5–10 m away from shade tree). Cocoa density was relatively uniform (1100 trees ha−1) with approximately 25 trees per 10 m radius plot
Species description
Detailed shade species dimensions and nutrient characteristics are presented in Table 1. Albizia zygia (Fabaceae), a nitrogen fixing species, has a spreading crown architecture and an average height of 11 m at the study site. This genus is recommended as a shade tree for cocoa plantations in Ghana (Anim-Kwapong 2003). Milicia excelsa (Moraceae), a fast growing tall timber species, can reach 50 m in height and has branches only on the upper half of the trunk. It has been identified as a desirable species for cocoa production due to its structural nature (low crown/stem ratio) and ability to provide income as timber (Manu and Tetteh 1987). Newbouldia laevis (Bignoniaceae), a multi-purpose, native tree, is used mainly for shade and live yam stakes as well as for medicinal purposes. It is commonly small in stature with a narrow crown and is easily reproduced from cuttings of lead shoots or lateral roots (Amanor 1994). Minimal research has been conducted on this species.
Soil conditions
Soil particle size distribution (sand = 68.0 ± 3.53; silt = 14.0 ± 1.45; clay = 18.0 ± 1.64) was determined by the hydrometer method, soil pH (5.9 ± 0.07) by a 1:1 paste of water:soil, percent organic matter (3.4 ± 0.09) by oxidation and cation exchange capacity (17.5 cmol kg−1 ± 0.71) by leaching and subsequent atomic absorption spectrophotometry (Allen 1974). Bulk density (0.9 g cm−3 ± 0.02) was determined for each treatment by collecting a known volume of soil with a metal core placed into the top 20 cm of soil, drying (105 °C, 48 h) and weighing the soil. All tests were conducted in triplicate.
Soil sampling
Soil fertility was assessed by collecting three composite soil samples (approximately 100 g per sample), to a depth of 0–20 cm [the active lateral root zone of cocoa (Kummerow et al. 1982)] in each plot at the two locations (<5 m and 5–10 m) from the shade tree. At the sole cocoa treatment, six random composite soil samples were collected. Soil samples were air-dried and sieved to pass 2 mm. Samples were wet-digested and analyzed for total N (Kjeldahl method), available P by Bray’s method and measured colorimetrically using molybdate method, and exchangeable K by leaching air-dried soil samples with ammonium acetate and measured quantitatively by atomic absorption spectrophotometry (Allen 1974). Estimates of soil nutrient stocks (kg ha−1) were based on measured bulk density and a 20 cm sampling depth.
Plant sampling
Aboveground biomass for the upper shade stratum and the cocoa stratum were estimated from diameter at breast height (DBH) measures (n = 15 per distance). Since no species-specific biomass equation was available in the literature, we used a generalized tree biomass equation developed for this specific precipitation zone (Brown 1997):
where y = dry aboveground biomass (kg) and DBH = diameter at breast height (cm). This equation is considered suitable for estimating total aboveground tree biomass of individuals with <150 cm DBH in relatively dense stands (Brown 1989, 1997), and is recommended for aboveground biomass estimation where destructive sampling can not be conducted (Anderson and Ingram 1993), as in our case due to farmer set restrictions. Although cocoa pod yield is the focal issue in farm production, our dependent variable was cocoa biomass that functions as an indicator of long-term farm sustainability. There is usually a strong correlation between cocoa tree components, biomass and pod yield under low shade intensity (Zuidema et al. 2005). Height (m), DBH (cm) and crown radius (m) of the three upper canopy trees were measured and listed in Table 1. Cocoa and shade tree densities were calculated from the number of trees per 20-m2 subplot extrapolated to a per hectare basis.
Samples of foliage, branch (approximately 100-g wet weight for each component type) and stem (approximately 40-g wet weight) tissue were collected from three randomly selected cocoa trees for all treatments and both locations (<5 m and 5–10 m). Foliage sampling was consistent for all individuals as recent and fully mature leaves from current season’s growth around the basal perimeter of the crown (shaded leaves). Newer branches were sampled from the lower perimeter of the crown, and stem tissue was collected with a sampling corer at DBH. Samples were oven-dried (70 °C) for 72 h and ground in a Wiley Mill (n = 9 per distance). Foliage and bulked woody tissue (composite branch and stem) were wet digested separately with hydrogen peroxide and sulfuric acid and analyzed for total N by auto-analysis, P by molybdate method and K by atomic absorption spectrophotometry (Allen 1974). Subsequently, woody tissue and foliage concentrations were averaged for each individual and multiplied by total aboveground dry weight to derive plant nutrient content. Similarly, foliage material (approximately 100-g wet weight for each species) was randomly collected from the three upper canopy species and analyzed by similar methods.
Light transmission measurements
Estimates for light transmission both above and below the cocoa strata were based on image analysis. A hemispherical lens (180-degree equidistant fisheye lens) was attached to a digital camera (Nikon Cool Pix 950), mounted onto a tripod, oriented north and leveled for each photo. Light transmission through the shade tree was measured on four transects at a 1 m radial distance from the shade tree above the cocoa stratum for each shade tree (approximately at a 5 m height) (n = 12). Images were examined by gap light analysis (Gap Light Analyzer, Version 2 1999), providing estimates of transmission indices, specifically percent canopy openness (% open sky beneath the canopy) for each shade tree species.
Statistical analysis
Data were analyzed as one-way analysis of variance (ANOVA) using the Proc GLM in SAS version 8.0 (SAS Institute Inc. Cary, NC, USA). Significant ANOVA’s were subsequently assessed by a Tukey’s test (HSD) and a Type I error rate was set at 0.05 for all statistical tests. Nutrient concentration and nutrient content were compared to monoculture cocoa using the Dunnett test. T-tests were applied to assess parameter differences close (<5 m) and far (5–10 m) from shade trees. Independence, randomness of residuals and a mean error equal to zero were confirmed with a test of residuals for biomass, biomass nutrient concentration and soil nutrient data. Normality of residuals was tested using the Shaprio–Wilk test.
Vector analysis
Dry mass and nutrient status of the plants was examined by vector analysis to asses nutritional response associated with the three agroforestry treatments (Haase and Rose 1995; Imo and Timmer 1997). The vector diagrams (Fig. 2) reflect the function that nutrient content (or amount) in a plant (bottom horizontal axis) is the product of its nutrient concentration (vertical axis) multiplied by its biomass (top horizontal axis). Changes in these parameters relate to two key processes driving plant growth: nutrient uptake and dry matter production that characterize differing treatment responses. Responses are expressed relative to the control (that is normalized to 100) to facilitate comparisons between various treatments and nutrients. Individual responses are depicted by vectors (arrows) that may differ in length and direction. Vector length represents response magnitude, and vector direction identifies specific nutritional responses. Thus a treatment induced increase in both nutrient uptake and dry mass with decreased nutrient concentration (Shift A) would reflect a dilution of this nutrient due to accelerated growth. A similar response without change in concentration (Shift B) signifies sufficiency of this nutrient since concentration was stable and kept up with increased growth and nutrient uptake. Increased biomass, nutrient concentration, and nutrient content (Shift C) would reflect an enrichment response to a deficiency because both growth and nutrient uptake improved. A similar response without biomass change (Shift D) would suggest luxury consumption because nutrient uptake was enhanced without growth increase. Other vector shifts depicting toxicity (Shift E, increased concentration with reduced growth and nutrient uptake) and antagonistic (Shift F, declines in the three parameters) responses were not evident in our study, but are described in more detail by Haase and Rose (1995) and Imo and Timmer (1997). The cocoa monoculture treatment was designated as the control, or reference(R), in our analysis.

Interpretation of directional changes in relative dry mass and nutrient status of plants depicted in a vector diagram. The reference condition (R) is normalized to 100. Diagnosis (A to F) is based on shifts (increase [+], decrease [−] or no change [0]) of individual nutrients characterized in response curves relating plant growth, nutrient concentration, and nutrient content to treatment (modified from Imo and Timmer 1997)
Results and discussion
Cocoa biomass
Biomass of cocoa trees was significantly higher for trees grown under shade than with no shade (P = 0.004; Table 2). Aboveground biomass was almost doubled under Milicia and Albizia canopies as compared to sole crop culture (41.0, 39.6 vs 22.8 Mg ha−1 for Milicia + cocoa, Albizia + cocoa and sole cocoa, respectively). Cocoa trees closer to the shade tree (<5 m) were significantly larger than those further away (5–10 m). Only the Milicia + cocoa treatment exhibited more cocoa biomass (21.97 kg per tree) at a greater distance as compared to monoculture cocoa (Table 2). Biomass estimates were within the range of other reported measures (Fassbender et al. 1991) and specifically for this region (Isaac et al. 2005). Our results support eariler studies that indicate cocoa biomass benefits from shade (Alpizar et al. 1986; Fassbender et al. 1991; Beer et al. 1998; Isaac et al. 2005).
Light availability
Percent open canopy between the cocoa and the shade strata was significantly different (P = 0.0136; Table 1). Canopy openness was similar for Albizia and Milicia (69.6 and 68.1%, respectively), but higher for Newbouldia (80.1%). However, light infiltration under all three species was within the optimal range for cocoa growth and pod yield, where growth is generally maximized at low shade levels as photosynthetic rates of cocoa decrease at high light intensities (Galyuon et al. 1996; Zuidema et al. 2005). Thus here, higher cocoa biomass was found under low levels of shade, as previously suggested in model simulations of biomass and pod yield under shade (Zuidema et al. 2005). However, crown architecture of individual species may affect light infiltration. Milicia and Albizia are characterized by a large, sparse and spreading crown, where light infiltration may be regulated over distance and, in particular for Albizia, allows for more evenly distributed sun flecks to reach the cocoa stratum (Manu and Tetteh 1987; Anim-Kwapong 2003). Newbouldia, with a narrow and dense canopy (Amanor 1994) presumably provides higher shade close to the upper canopy tree.
Soil nutrient stocks
Soil fertility status at both distances from the shade trees are given in Table 3. Available P pools in the top soil were smaller (P < 0.001) under Milicia + cocoa (19.7 kg ha−1) and Newbouldia + cocoa (19.5 kg ha−1) compared to monoculture pools (22.2 kg ha−1; Table 3), inferring depletion of this nutrient close to shade trees. Accelerated growth in agroforestry systems may increase demands on soil P supply (Ofori-Frimpong and Rowel 1999). This may result in enlarged P depletion zones, possibly inducing interspecific competition for soil P resources (Vandermeer 1989). However, P availability under Albizia + cocoa (22.7 kg ha−1) was comparable to the monoculture cocoa, indicating a lack of competition for P resources under Albizia. Nutrient transfer is often higher under N-fixing species in comparison to a non-N fixing timber species (Fassbender et al. 1991) possibly due to improved litter quality, which may accelerate nutrient cycling processes in the 5 m zone around the shade trees.
Total soil N was similar under shade for all three shade + cocoa treatments and monoculture cocoa, but decreased with distance under Newbouldia + cocoa, presumably due to Newbouldia’s narrow crown and lower structure thus impacting a smaller area. The expectation was to find elevated soil nitrogen under the N-fixing species Albizia, although no increases were detected. This result is not uncommon, as limited contribution from N-fixation in mature agroforestry systems has previously been reported (Nygren and Ramirez 1995; Beer et al. 1998). As there was no marked difference in soil nitrogen pools, improved cocoa biomass was likely associated with rapid nutrient cycling rather than N-fixation (Beer 1988).
Exchangeable K in the top soil pool was higher under Newbouldia + cocoa (214.5 kg ha−1) as compared to the monoculture (141.4 kg ha−1) and the other shade treatments (Table 3). Generally, low re-absorption of K before cocoa litterfall and the high K transfer via rainwash (Boyer 1973; Hartemink 2005), combined with Newbouldia’s particularly narrow and dense canopy may result in increased accumulation under this species. Soil exchangeable K also decreased with distance from Newbouldia, supporting our suggestion of a zone of concentrated K under Newbouldia (Table 3). As found in previous studies, larger K soil pools lead to higher concentrations of K in cocoa biomass (Boyer 1973), suggesting considerable plant uptake of potassium, and a possibility of luxury consumption.
Plant nutrition
Nutrient interactions of the cocoa plants under the different species and distances from shade trees are illustrated in Fig. 3, where the relative biomass, nutrient concentrations and nutrient content of the cocoa plant were compared in a single nomogram with monoculture cocoa normalized to 100. At <5 m distance, cocoa trees achieved the highest biomass response (35–80%) compared to a lower response (−5% to 6%) further away (5–10 m from shade tree). Concentration levels declined with P, but remained unchanged with N, and increased with K (Table 4). These changes (Fig. 3) reflect vector shifts A, B and C that signify, respectively, dilution of P, sufficiency of N, and deficiency of K in cocoa growth (Fig. 2). Under shade K uptake was stimulated most (96–140%), followed by N (43–88%), and then P (22–45%). With increasing distance from the shade tree, K uptake was enhanced most (22–34%), followed by P (9–21%), and then N (−3% to 13%).

Nomograms of relative response in dry mass and nutrient content (N, P and K) of cocoa trees under Albizia, Milicia or Newbouldia. Monoculture cocoa was normalized to 100. Both radial distances are shown on the nomogram: <5 m and 5–10 m away from the shade tree
A consistent trend of P dilution (Shift A; Fig. 2) in cocoa under each species would suggest that the next limiting nutrient on this farm would be phosphorus (Fig. 3). Near the shade tree, P uptake was least (22–45%) in comparison to the other nutrients (N = 43–88%; K = 96–140%), reflecting relatively slower cycling of this element (Fassbender et al. 1991). Phosphorous status in soils (Table 3) and in cocoa biomass (Table 4) was lower under shade trees than in monoculture. Low phosphorous status is common in tropical agricultural systems, because of widespread P deficiency in highly weathered acid tropical soils and is usually most limiting within tropical agroforestry systems (Alpizar et al. 1986). Similar to here, cocoa-shade agroforestry systems in Costa Rica reported low P accumulation in crops, attributed to relatively low phosphorus mobility within these systems (Fassbender et al. 1991; Table 4). In our case, soil P may become limiting if concentrations cannot be maintained with accelerated cocoa growth under shade.
There was no significant difference in total N concentration in cocoa under all three species (Table 4; Fig. 3), suggesting little contribution from our shade species towards N transfer. Despite higher tree foliar N levels (Table 1), N-fixing Albizia failed to raise soil N or cocoa N status compared to the other species. Although N concentration was stable with enhanced growth under shade trees, N content increased across all treatments in comparison to the monoculture cocoa. Apparently, N uptake kept pace with growth, a common sufficiency response (Table 4; Fig. 3). Overall, nitrogen content values were comparable to a 9 year-old cocoa agroforestry system with Cordia alliodora and Erythrina poeppigiana in Costa Rica (Alpizar et al. 1986).
Potassium concentration was significantly higher in cocoa under Albizia and Newbouldia close to the shade tree, presumably due to corresponding higher K levels in soils under this tree (Table 3). However, potassium uptake by cocoa near the shade tree (<5 m) was enhanced in all multistrata treatments, especially under Albizia (140%) where K concentration rose 40%, hence signifying a typical deficiency response (Shift C; Fig. 2). On the other hand, with increasing distance from the shade tree, K contents (Albizia + cocoa = 355.6 g tree−1; sole cocoa = 265.4 g tree−1) were increased without substantially increasing N, P and biomass status (Table 4; Fig. 3). These vector patterns reflect luxury consumption (Shift D; Fig. 2) of potassium, since K accumulation occurred without increased biomass. Vector length comparisons showed that K was the most responsive element within this system, exhibiting enhanced uptake even with distance from shade tree. As noted in Table 3, the multistrata systems resulted in either a neutral or positive effect on soil exchangeable K pools, probably because of high K inputs from litter and elevated K transfer via rainwash (Boyer 1973; Hartemink 2005). Cycling of subsoil K to the surface through shade tree uptake may also occur, because after N, potassium is usually considered more readily mobile as compared to other macronutrients (Barber 1984). Specifically for K, our results illustrate a possible facilitating effect of nutrient pumping under shade trees, likely because of enhanced nutrient transfer by upper canopy trees.
Conclusions and implications
The inclusion of low-density shade trees in this 8-year-old cocoa plantation positively affected biomass of neighboring cocoa trees through nutrient and/or light manipulation. Aboveground dry matter of cocoa increased with shade, and declined along a spatial gradient away from shade trees. Light availability under all three shade tree species was within the optimal range for cocoa production. Although soil exchangeable K was increased under Newbouldia + cocoa, in general, available P decreased and total N stocks were unaffected in the soils under all shade treatments. Vector analysis showed that nutrient uptake of cocoa was highest near shade trees, and that K was the most responsive nutrient in these systems. Research trials that incorporate fertilizers (in particular K) and shade manipulation are recommended to further elucidate the specific mechanisms involved in these responses.
Our study illustrates a beneficial relationship between shade trees and enhanced cocoa biomass presumably due to provision of optimal shade and overall improved nutrient cycling capabilities near the shade trees. Nutrient uptake was maintained with accelerated cocoa growth under shade suggesting a potential for sustained productivity. We found minimal variation in the level of complementarity of each upper canopy species, however, appropriate management of upper canopy trees may vary over time. Either one of these factors may emerge more important at different points in the life of the plantation. Presumably, young cocoa trees are more sensitive to high light and require shading, however in mature plantations, the main role of upper canopy trees may shift from providing shade to enhancing nutrient stability in both soils and biomass. Therefore, farmer management of the upper canopy is critical for optimal levels of cocoa productivity and sustainability.
References
Ahenkorah Y, Halm BJ, Appiah MR, Akrofi GS, Yirenkyi JEK (1987) Twenty years’ results from a shade and fertilizer trial on Amazon cocoa (Theobroma cacao) in Ghana. Exp Agric 23: 31–39
Allen SE (eds) (1974) Chemical analysis of ecological materials. John Wiley & Sons, New York
Alpizar L, Fassbender HW, Heuveldop J, Folster H, Enriquez G (1986) Modeling agroforestry systems of cacao (Theobroma cacao) with laurel (Cordia alliodora) and poró (Erythrina poeppigiana) in Costa Rica. I Inventory of organic matter and nutrients. Agrofor Syst 4:175–189
Amanor KS (1994). The new frontier; farmers response to land degradation. A West African study. UNRISD, London, England, pp 171
Anderson JM, Ingram JSI (eds) (1993). Tropical soil biology and fertility: a handbook of methods. CAB International, Wallingford, UK, pp 10–15
Anim-Kwapong GJ (2003) Potential of some Neotropical Albizia species as shade trees when replanting cacao in Ghana. Agrofor Syst 58: 185–193
Barber SA (1984) Soil nutrient bioavailability: a mechanistic approach. John Wiley and Sons, Inc. Canada, pp 229–256
Beer J (1988) Litter production and nutrient cycling in coffee (Coffea arabica) or cacao (Theobroma cacao) plantations with shade trees. Agrofor Syst 7:103–114
Beer J, Muschler R, Kass D, Somarriba E (1998) Shade management in coffee and cacao plantations. Agrofor Syst 38:139–164
Boyer J (1973) Cycles de la matiere organique des elements mineraux dans une cacaoyere camerounaise. Cafe´ Cacao The´ 18:3–30
Brown S (1989, 1997) Estimating biomass and biomass change of tropical forests: a primer
Cunningham RK, Arnold PW (1962) The shade and fertilizer requirements of cacao (Theobroma cacao) in Ghana. J Sci Food Agr 13: 213–221
Ewel JJ, Bigelow SW (1996) Plant life-forms and tropical ecosystem functioning. In: Orians G, Dirzo D., Cushman C (eds) Biodiversity and ecosystem processes in tropical forests, Springer-Verlag, Berlin, Germany, pp 101–123
FAO Forestry Paper 134. Food and Agriculture Organization of the United Nations, Rome, Italy
Fassbender HW, Beer J, Heuveldop J, Imback A, Enriquez G, Bonnemann A (1991) Ten year balances of organic matter and nutrients in agroforestry systems at CATIE, Costa Rica. For Ecol Manage 45:173–183
Galyuon IKA, McDavid FB, Lopez FB, Spence JA (1996) The effect of irradiance level on cocoa (Theobroma cacao L.): 1. Growth and leaf adaptations. Tropical Agriculture Trinidad 73:23–28
Gap Light Analyzer, Version 2 (1999) Simon Fraser University, British Columbia, Canada and the Institute of Ecosystem Studies, New York, USA
Haase DL, Rose R (1995) Vector analysis and its use for interpreting plant nutrient shifts in response to silvicultural treatments. For Sci 41:54–66
Hartemink AE (2005) Nutrient stocks, nutrient cycling, and soil changes in cocoa ecosystems: a review. Adv Agro 86:227–253
Hooper DU (1998) The role of complementarity and competition in ecosystem responses to variation in plant diversity. Ecology 79:704–719
Imo M, Timmer VR (1997) Vector diagnosis of nutrient dynamics in mesquite seedlings. For Sci 43: 268–273
Isaac ME, Gordon AM, Thevathasan N, Oppong SK, Quashie-Sam J (2005) Temporal changes in soil carbon and nitrogen in west African multistrata agroforestry systems: a chronosequence of pools and fluxes. Agrofor Syst 65:23–31
Kummerow J, Kummerow M, Da Silva WS (1982) Fine root growth dynamics in cacao (Theobroma cacao). Plant Soil 65:193–201
Manu M, Tetteh EK (1987) A guide to cocoa cultivation. Cocoa Research Institute of Ghana, Ghana Cocoa Board, Accra
Nygren P, Ramírez C (1995) Production and turnover of N2 fixing nodules in relation to foliage development in periodically pruned Erythrina poeppigiana (Leguminosae) trees. For Ecol Man 73:59–73
Ofori-Frimpong K, Rowell DL (1999) The decomposition of cocoa leaves and their effect on phosphorus dynamics in tropical soil. Euro J Soil Sci 50:165–172
Schroth G, Lehmann J, Rodrigues MRL, Barros E, Macedo JL (2001) Plant-soil interactions in multistrata agroforestry in the humid tropics. Agrofor Syst 53:85–102
Silver WL, Brown S, Lugo A (1996) Biodiversity and biogeochemical cycles. In: Orians G, Dirzo D, Cushman C (eds) Biodiversity and ecosystem processes in tropical forests. Springer-Verlag, Berlin, Germany, pp 49–65
Soil Survey Division. (1969). Great Soil Groups. Survey of Ghana, Accra, Ghana
Tilman D (1999). The ecological consequences of changes in biodiversity: a search for general principles. Ecology 80:1455–1474
Vandermeer J (1989) The Ecology of Intercropping. Cambridge University Press, Cambridge, UK, pp 68–85
Zuidema PA, Leffelaar PA, Gerristsma W, Mommer L, Anten NPR (2005) A physiological production model for cocoa (Theobroma cacao): Model presentation, validation, application. Agri Syst 84: 95–225
Acknowledgements
We would like to thank farmer collaborators in the Sefwi Wiawso District, Ghana. We are also grateful to the Faculty of Renewable Natural Resources, Kwame Nkrumah University of Science and Technology, Ghana, for research support. We would like to acknowledge field and laboratory assistance by J. Hagan, Y. Teng, and E. Benczkowski and constructive critical comments by two anonymous journal reviewers. Financial support for this study was provided by the Natural Science and Engineering Research Council of Canada.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Isaac, M.E., Timmer, V.R. & Quashie-Sam, S.J. Shade tree effects in an 8-year-old cocoa agroforestry system: biomass and nutrient diagnosis of Theobroma cacao by vector analysis. Nutr Cycl Agroecosyst 78, 155–165 (2007). https://doi.org/10.1007/s10705-006-9081-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10705-006-9081-3