
Abstract
The aim of this study is to verify levels of resistance to whitefly (Bemisia argentifolii) and the two-spotted spider mite (Tetranychus urticae) in experimental hybrids of tomato plants with high acylsugar content and that bear the Mi allele, and to compare the levels of resistance to those pests according to the presence or absence of acylsugars and the Mi allele. Eleven genotypes were tested in assessment of resistance to whitefly: two inbred lines, TOM-760 (Mi/Mi), with high acylsugar content, and TOM-684 (Mi/Mi), with low acylsugar content; six experimental hybrids heterozygous for acylsugar contents, TEX-407 (Mi+/Mi), TEX-408 (Mi+/Mi), TEX-409 (Mi+/Mi), TEX-410 (Mi+/Mi), TEX-411 (Mi+/Mi), and TEX-412 (Mi/Mi); Santa Clara cultivar (Mi+/Mi+) with low acylsugar content; and two control hybrids with low acylsugar content, Ibiza (Mi+/Mi) and Carina (Mi+/Mi). In the test for repellency to the two-spotted spider mite, we used the same genotypes as in the whitefly resistance test, except for the Carina hybrid. Genotypes homozygous for acylsugars and Mi, heterozygous for Mi/Mi or Mi+/Mi acylsugars, and homozygous and heterozygous for Mi alone showed levels of resistance to the Bemisia argentifolii fly, in contrast with the genotype with low contents of acylsugars and Mi+/Mi+. Genotypes homozygous and heterozygous for acylsugars exhibit higher levels of repellency to the Tetranychus urticae mite than genotypes with low contents of acylsugar. The Mi gene, however, was not effective in providing resistance to the two-spotted spider mite. The effects of resistance to Bemisia argentifolii shown by the acylsugars are added to the effects of the Mi gene.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The Bemisia tabaci whitefly (Gennadius) (Hemiptera: Aleyrodidae) is a pest of global agricultural importance, especially for tropical and subtropical regions (Prabhaker et al. 2005; Nauen and Denholm 2005). In recent years in Brazil, problems have grown and damage has intensified due to the aggressive nature of a biotype of the whitefly, the B biotype, described as Bemisia argentifolii Bellows et al. (1994). This pest can cause direct damage by feeding from the sap, altering the vegetative and reproductive development of plants and reducing the photosynthetically active area. Tomato fruits grow irregularly, making them improper for marketing and consumption. However, the most disturbing damage caused by whiteflies is indirect, for those insects are carriers of viruses, such as geminiviridae, causing irreversible physiological disorders and crop losses of up to 100% (Tavares 2002; Villas Boas et al. 2002).
Mites include a large number of species but in the Tetranychus genus, only three are important in growing tomatoes: the two-spotted spider mite T. urticae Koch, and the red spider mites T. ludeni Zacher and T. evansi Baker Pritchard; the first is the main species found in Brazil (Flechtmann 1989). Although they are pests of secondary importance in Brazil for the tomato plant, they may might cause direct damage in environments with high infestation and lead to leaf drying and leaf drop, which makes for smaller and fewer fruits and induces early maturity (Flechtmann and Baker 1970).
Control of tomato plant pests still depends greatly on chemical control because, at this time, there are not yet cultivars with sufficiently high levels of resistance to substantially reduce the need for spraying. Consequences of indiscriminate use of insecticides might include harm to natural enemies of pests when insecticides are not selective, and they may promote selection of individuals resistant to the chemicals used (Silva et al. 2009b; Petroski and Stanley 2009).
However, strategies such as the use of cultivars with high leaf content of allelochemicals, especially acylsugars antagonistic to pests, have proven to be feasible in control via genetic resistance (Resende et al. 2006; Gonçalves Neto 2008; Maluf et al. 2010; Maciel et al. 2011). Many authors have studied and confirmed this resistance, associated with the acylsugar allelochemical, to the two-spotted spider mite (Tetranychus urticae) (Silva et al. 2009a; Marchese 2013; Oliveira 2015), the red spider mite (Tetranychus evansi) (Gonçalves 2006; Resende et al. 2008), whitefly (Bemisia argentifolii) (Resende et al. 2009; Silva et al. 2009a; Marchese 2013; Oliveira 2015), the tomato leafminer (Tuta absoluta) (Resende et al. 2006; Maciel et al. 2011), the green peach aphid (Myzus persicae), and the serpentine leafminer (Liriomyza trifolii) (Silva 2012; Silva et al. 2013).
There are reports of other mechanisms related to pest resistance in tomato plants. The Mi gene grants effective resistance to three important species of cotton root-knot nematodes, Meloidogyne incognita, M. javanicae, and M. arenaria (Roberts and Thomason 1989). However, studies show that the Mi gene also offers resistance to other pests, such as the potato aphid (Macrosiphum euphorbiae) (Rossi et al. 1998), the two-spotted spider mite (T. urticae) (Godzina et al. 2011), and whitefly (Bemisia argentifolii) (Nombela et al. 2000; Marchese 2013; Oliveira 2015). For whitefly, according to Nombela et al. (2000), the presence of the Mi gene in tomato cultivars reduced the number of females and adults up to 50%. Marchese (2013) and Oliveira (2015) also noticed a reduction in the number of eggs and nymphs of Bemisia argentifolii because of the Mi gene, even though this effect had a smaller magnitude than the one promoted by the action of leaf allelochemicals. However, there are no studies in the literature showing the effect of simultaneous use of high allelochemical contents and the Mi gene on resistance to whitefly.
In regard to the two-spotted spider mite, Godzina et al. (2011) found a smaller population density in tomato plants with the presence of the dominant allele in homozygosity for the Mi gene locus than in those that did not have the allele. However, Marchese (2013) and Oliveira (2015) did not notice any effect of the Mi gene on repellency to the two-spotted spider mite (T. urticae).
The aim of this study is to verify levels of resistance to whitefly and two-spotted spider mites in experimental tomato hybrids with high acylsugar content and that bear the Mi allele, and to compare the levels of resistance of the tomato plant to those pests according to the presence or absence of acylsugars and the Mi allele.
Materials and methods
Genotypes evaluated
Two inbred lines (TOM-760 and TOM-684), six experimental hybrids (TEX-407, TEX-408, TEX-409, TEX-410, TEX-411, and TEX-412), the Santa Clara cultivar, and two commercial-control hybrids (Ibiza and Carina) were tested in assessment of resistance to whitefly. In the test of repellency to the two-spotted spider mite, we used the same genotypes as in the whitefly resistance test, except for the Carina hybrid (Table 1).
The TOM-684 line bears (in homozygosity) the Mi gene, which grants resistance to nematodes of the Meloidogyne spp. genus. Mi allele is linked in the repulsion phase to the Ty-1 allele, which grants resistance to begomovirus. It has low leaf contents of acylsugars (Marchese 2013). The TOM-760 line, also Mi/Mi homozygous, has the same genotypic background as TOM-684, but it differs in acylsugar content; TOM-760 has acylsugar content comparable to TOM-687 and TOM-688 (information acquired personally from Wilson Roberto Maluf), which have high leaf content (Marchese 2013).
The six experimental hybrids were obtained by crossing elite lines from the breeding program of Hortiagro Sementes S.A. (which have low acylsugar content) with the TOM-760 line. Hybrids between a parent with high acylsugar content and a parent with low content have intermediate allelochemical levels between those from the parent lines (Maciel 2008; Maluf et al. 2010). Therefore, they are higher than those of the low content parent. Consequently, all the experimental hybrids have intermediary acylsugar contents. TEX-407, TEX-408, TEX-409, TEX-410, and TEX-411 are heterozygous (Mi+/Mi) in the Mi locus, whereas TEX-412 is homozygous (Mi/Mi).
Ibiza from Hortiagro Sementes S.A. and Carina from Sakata Seed Sudamerica Ltda are commercial hybrids of indeterminate growth habit. Both have low acylsugar content and the Mi locus in heterozygosity.
Santa Clara is a commercial cultivar of indeterminate growth habit with low acylsugar leaf content, susceptible to nematodes (does not carry the Mi allele), and with known susceptibility to whitefly and the two-spotted spider mite (Marchese 2013; Oliveira 2015).
Assessment of whitefly resistance (Bemisia argentifolii)
The genotypes were sown at the Experimental Station of the Hortiagro Sementes S.A. company in 162-cell plastic trays containing Tropstrato® substrate on August 23, 2015, and then transplanted 30 days after in polyethylene pots with a capacity of 5 L of substrate and kept in greenhouses.
For whitefly (B. argentifolii) infestation, insects were previously raised in a greenhouse. Whiteflies were collected from tomato crops; they were released in the greenhouse, where kale plants (Brassica oleracea var. acephala) served as a substrate for multiplication. When the whitefly population was high, the tomato plants were transferred to a greenhouse on the UFLA Campus to be assessed, where the resistance test was conducted. The experiment was set up in a randomized block design (RBD) with 11 treatments (genotypes) and seven replications; a pot with a single plant constituted a plot.
The genotypes were assessed in regard to oviposition from 4 to 14 days after infestation by counting the number of eggs, taking samples of four leaflets of the third completely expanded leaf from the upper third of each plant and examining them with the aid of an entomological magnifying glass of 10 times magnification; the numbers of eggs in 1 cm2 of leaf area were recorded. From 19 to 26 days after infestation, the number of nymphs was assessed in the same sample leaflets, which had been identified with a wool thread, once more with the aid of an entomological magnifying glass of 10 times magnification and in an area of 1 cm2.
Test of repellency to the two-spotted spider mite (Tetranychus urticae)
Resistance to the two-spotted spider mite was measured through the test of repellency developed by Weston and Snyder (1990). A randomized block design was used, with 6 replications. Similar sized expanded leaflets were taken as samples from the upper third of the plant. The mite population was obtained from the mite collection raised by EPAMIG Sul de Minas; mites identified as Tetranychus urticae were used, which were multiplied in jack bean plants (Canavalia ensiformis).
The test was conducted in a climate controlled chamber at 16 ± 1 °C and 64 ± 4% relative humidity. Leaflets of each one of the genotypes were fastened in the central region of a sheet of sulfite paper on a polystyrene board with a thumbtack (9 mm diameter), with the adaxial surface turned upward. The leaflets were placed at random on the polystyrene board. Two female mites were transferred to the center of each thumbtack with the aid of a thin brush. The mean distances (mm) covered by the mites over the surface of each leaflet were measured as of the outer extremity of the thumbtack after 20, 40, and 60 min of exposure. For those who remained on the thumbtack, the distance was considered as zero. Smaller distances covered by the mites were considered signs of greater levels of repellency (Marchese 2013; Oliveira 2015).
Statistical analysis
Analysis of variance was performed for the resistance data of whitefly and two-spotted spider mites, and the means of the genotypes were compared by the Tukey test at the 5% level of probability. Estimates of contrasts selected between groups of genotypes with different contents of acylsugars and/or different genotypic constitutions in the Mi locus were obtained in order to characterize possible differences in levels of pest resistance, in accordance with those two characteristics. Analyses were made through the statistical application Sisvar (Ferreira 2000).
Results and discussion
Resistance to whitefly
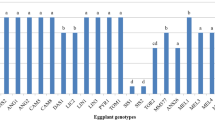
A significant difference, at 5% by the Tukey test, was observed between the genotypes for number of eggs and number of nymphs (Table 2). TOM-760 and TOM-684 had smaller average values in number of eggs and nymphs than the ‘Santa Clara’ control (Table 2). Both homozygosity for Mi alone (Table 3, C3) and homozygosity for Mi and for high acylsugar content (Table 3, C2) are effective against whitefly, but the effects of acylsugars add to the Mi gene, such that TOM-760 is more resistant than TOM-684, with the tendency toward decline in oviposition of the insect (Table 3, C1). Both homozygosity for Mi and high acylsugar content (Table 3, C4) and homozygosity for Mi alone (Table 3, C5) are likewise more effective against whitefly than heterozygosity in Mi-, which, in turn, is more effective than the genotype susceptible to nematodes and with low acylsugar (Table 3, C6). This shows that both Mi and acylsugars are effective in resistance to whitefly, and that the effect of Mi is more accentuated in homozygosity.
TOM-760 had a smaller number of eggs and nymphs than TEX-412 (Table 3, C9), whereas TOM-684 had higher numbers than TEX-412 (Table 3, C10). Though the contrasts were not significant in these cases, the joint interpretation of C9 and C10 is that the hybrid homozygous for Mi and heterozygous for acylsugar (TEX-412) has intermediate levels of resistance, between the levels of TOM-760 (Mi homozygous and high acylsugar content) and TOM-684 (Mi homozygous and low acylsugar content). Therefore, even in the presence of Mi in homozygosity, heterozygosity (intermediate content) for acylsugar is effective in respect to promoting additional resistance to whitefly. The C7 contrast (Table 3) shows that the combination of Mi in homozygosity associated with high contents of acylsugar also in homozygosity is more effective than both Mi and acylsugar content in heterozygosity; hybrids with this last combination were similar to TOM-684, homozygous in the Mi locus (Table 3, C8). The C14 contrast (Table 3) shows that Mi and acylsugar content, both in association and in heterozygosity, are highly effective in granting resistance to Bemisia argentifolii. The C12 contrast (Table 3) indicated that use of the technology of higher acylsugar content (in heterozygosity) in backgrounds that are also heterozygous for the Mi gene have an additional effect in promoting resistance to whitefly in comparison to use of Mi alone in heterozygosity (Table 1 and Table 3, C12).
In the presence of acylsugar content in heterozygosity, the difference in resistance granted by Mi in homozygosity compared to Mi in heterozygosity is negligible (Table 3, C11) but may not be null, which is shown by C7 and C9, Table 3: the hybrids heterozygous for the Mi locus had lower resistance to whitefly, indicated by the number of nymphs, than TOM-760 (homozygous for Mi and for high acylsugar content) (Table 3, C7), whereas the Mi homozygous hybrid (TEX-412) exhibited levels of resistance similar to TOM-760 (Table 3, C9). Yet both genotypic constitutions are effective in granting resistance to whitefly, and they have higher resistance to Bemisia argentifolii than genotypes with lower acylsugar content, whether they are resistant to nematodes (Carina and Ibiza) (Table 3, C12 and C13) or not (Santa Clara) (Table 3, C14 and C15).
Maluf et al. (2010) documented resistance to whitefly in hybrids between acylsugar rich parent lines [TOM-687 (Mi+/Mi+), TOM-688 (Mi+/Mi+), and TOM-689 (Mi+/Mi+)] and acylsugar poor parent lines [TOM-684 (Mi/Mi), TOM-690 (Mi+/Mi+)]. The results of the present study, similar to those presented by Maluf et al. (2010), show that in using only one parental line with high content of this alellochemical, it is possible to obtain hybrids with high contents that have satisfactory levels of resistance to Bemisia argentifolii. However, the Mi homozygous line (TOM-684) had a higher number of eggs than the Mi+ homozygous line (TOM-690) in Maluf et al. (2010) and was not different in relation to the number of nymphs, indicating that, at least in that test, the Mi gene was not very effective in granting non-preference for oviposition.
Kaloshian et al. (1995) report that, even though a reaction of complete dominance of the Mi allele for resistance to nematodes occurs, studies regarding resistance to the Macrosiphum euphorbiae aphid show that there is different behavior for the aphid between homozygous (Mi+/Mi+) and heterozygous (Mi/Mi+) genotypes, indicating that there is possible incomplete dominance of this gene or of a linked gene that grants this resistance. Data of the present study indicate that the effect of the Mi gene for whitefly resistance also has incomplete dominance, corroborating Kaloshian et al. (1995).
Nombela et al. (2003) report that the presence of the Mi gene reduced the average number of nymphs per leaflet by 50% compared to plants that did not have the resistance allele. Marchese (2013) found a reduction for eggs and nymphs of approximately 20% in plants with the Mi gene compared to the ‘Santa Clara’ control. The reduction in the number of eggs and nymphs (around 70%) in the line homozygous for the Mi allele (TOM-684) compared to the ‘Santa Clara’ susceptible genotype was bigger in the present study than the reduction reported in papers by Nombela et al. (2003) and Marchese (2013). The results of this study agree with Nombela et al. (2000) in showing that the Mi gene, or another gene closely linked to it, grants resistance to whitefly (B. argentifolii).
Our results confirm that high leaf acylsugar contents are effective in granting resistance to whitefly, as described by other authors (Resende et al. 2008; Silva et al. 2009a; Maluf et al. 2010; Marchese 2013), and suggest that the effects of acylsugars and the Mi gene are greater in homozygosity than in heterozygosity. They also demonstrate that the effects of the resistance granted by acylsugars are added to the effects of the Mi gene. Therefore, the technology characterized by the presence of the Mi gene and high acylsugar contents can be effectively used to grant resistance to Bemisia argentifolii.
Repellency to the two-spotted spider mite
The genotype tests exhibit significant differences at 5% probability according to the Tukey test concerning repellency to the two-spotted spider mite at 20, 40, and 60 min (Table 4).
The walking distances (displacement) of the spider mites over the leaflets at 20 min in the line with high acylsugar content (TOM-760) were significantly smaller than in the TOM-684 line (Table 5, C1) and in the experimental hybrids heterozygous (Table 5, C7) and homozygous (Table 5, C9) for the Mi gene (and heterozygous for acylsugars). The same happened at 20 and 40 min in comparison to Ibiza (Table 5, C4), and in every time assessed in comparison to the ‘Santa Clara’ control (Table 5, C2). These results demonstrate the efficiency of the acylsugar allelochemical in resistance and/or repellency to spider mites of the Tetranychus genus, as already documented by other authors (Silva et al. 2009a; Maluf et al. 2010; Marchese 2013).
Among the genotypes with low acylsugar content (Santa Clara, TOM-684, and Ibiza), there were no significant differences, regardless of the genotypic constitution for the Mi gene (Table 4 and Table 5, C3, C5, and C6). In a similar study, Marchese (2013) and Oliveira (2015) verified that homozygous lineages for Mi did not change repellency to the two-spotted spider mite compared to susceptible controls. However, Godzina et al. (2011) reported that the spider mite (T. urticae) developed slowly in tomato plants with the Mi gene. The authors noted incomplete dominance of the resistance under field conditions, in which the heterozygous genotype for the Mi gene locus showed intermediate levels of resistance to the spider mite. The results of this study show that the Mi gene does not have an effect on repellency to the two-spotted spider mite, corroborating Marchese (2013) and Oliveira (2015) and contrasting with Godzina et al. (2011). Studies suggest the Mi gene influences the structure of the phloematic content of the plant, which repels sucking arthropods (Kaloshian et al. 2000). Marchese (2013) suggests that one of the reasons for which T. urticae spider mites are less sensitive to plants with the Mi gene is that they basically feed from the leaf mesophyll and are not sucking insects.
The average repellency to the T. urticae two-spotted spider mite granted by the experimental hybrids heterozygous for acylsugar contents and resistance to nematodes were greater than the ‘Santa Clara’ control at every time assessed (Table 5, C14), greater than Ibiza at 40 and 60 min (Table 5, C12), and greater than TOM-684 at 40 min (Table 5, C8). TEX-412, heterozygous for acylsugar contents and Mi/Mi homozygous, was not significantly different from TOM-684 (Table 5, C10), Ibiza (Table 5, C13), or Mi+/Mi experimental hybrids (Table 5, C11) at any of the times assessed, but it exhibited shorter displacement on the part of the spider mites than the ‘Santa Clara’ control (Table 5, C15). This shows that acylsugar contents, even when in heterozygosity (and therefore smaller than when in homozygosity), are effective in repelling the spider mite. The results of this study are similar to those of Maluf et al. (2010), who found moderate resistance to the two-spotted spider mite (Tetranychus urticae) in pre-commercial hybrids of the tomato plant, with intermediate acylsugar contents (hybrids between acylsugar rich parent lines—TOM-687, TOM-688—and acylsugar poor parent lines—TOM-684, TOM-690).
Conclusions
The genotypes homozygous for acylsugars and Mi, heterozygous for Mi/Mi or Mi+/Mi acylsugars, and homozygous and heterozygous for Mi alone showed levels of resistance to the Bemisia argentifolii fly in contrast with the genotype with low acylsugar contents and Mi+/Mi+. The homozygous and heterozygous genotypes for acylsugars show levels of repellency to the Tetranychus urticae mite in contrast with genotypes with low acylsugar contents.
The Mi gene, or another closely linked to it, provided a certain level of resistance to whitefly and seems to have incomplete dominance for resistance to this pest. However, this gene was not effective in granting resistance to the two-spotted spider mite.
The effects of resistance to Bemisia argentifolii provided by the acylsugars are added to the effects of the Mi gene.
The technology characterized by the presence of higher leaf acylsugar contents can be effectively used to grant resistance to the Tetranychus urticae mite and to the Bemisia argentifolii whitefly, and, in combination with the Mi gene, action against whiteflies is optimized.
References
Bellows TS Jr, Perring TM, Gill RJ, Headrick DH (1994) Description of a species of Bemisia (Homoptera: Aleyrodidae). Ann Entomol Soc Am. https://doi.org/10.1093/aesa/87.2.195
Ferreira DF (2000) Sistemas de análise estatística para dados balanceados. Universidade Federal de Lavras/DEX/SISVAR
Flechtmann CHW (1989) Ácaros de importância agrícola. Nobel, São Paulo
Flechtmann CHW, Baker EW (1970) A preliminary report on the Tetranychidae (Acarina) of Brazil. Ann Entomol Soc Am. https://doi.org/10.1093/aesa/63.1.156
Godzina M, Kielkiewicz M, Szymczykiewicz K (2011) Varying abundance and dispersal of the two-spotted spider mite (Tetranychus urticae Koch, 1836, Acari: Prostigmata: Tetranychidae) on Mi-tomato plants differing in allelic combination. Biol Lett. https://doi.org/10.2478/v10120-011-0022-8
Gonçalves LD (2006) Herança do teor de acilaçúcares em genótipos de tomateiro e sua relação com tricomas foliares e repelência ao ácaro Tetranychus evansi. Dissertation, Universidade Federal de Lavras
Gonçalves Neto AC (2008) Seleção para teor de acilaçucar nas folhas em tomateiros com qualidade comercial confere resistência à traça (Tuta absoluta). Dissertation, Universidade Federal de Lavras
Kaloshian I, Lange WH, Williamson VM (1995) An aphid-resistance locus is tightly linked to the nematode-resistance gene, Mi, in tomato. Proc Natl Acad Sci. https://doi.org/10.1073/pnas.92.2.622
Kaloshian I, Kinsey MG, Williamson VM, Ullman DE (2000) Mi-mediated resistance against the potato aphid Macrosiphum euphorbiae (Hemiptera: Aphididae) Limits Sieve Element Ingestion. Environ Entomol. https://doi.org/10.1603/0046-225X-29.4.690
Maciel GM (2008) Broad spectrum arthropod resistance mediated by leaf acylsugar contents in tomatoes. Dissertation, Universidade Federal de Lavras
Maciel GM, Maluf WR, Silva VF, Gonçalves NETOAC, Gomes LAA (2011) Híbridos pré-comerciais resistentes a Tuta absoluta obtidos de linhagem de tomateiro rica em acilaçúcares. Hortic Bras. https://doi.org/10.1590/S0102-05362011000200003
Maluf WR, Maciel GM, Gomes LAA, Cardoso MG, Gonçalves LD, Silva EC, Knapp M (2010) Broad-spectrum arthropod resistance in hybrids between high- and low-acylsugar tomato lines. Crop Sci Soc Am. https://doi.org/10.2135/cropsci2009.01.0045
Marchese A (2013) Resistência à mosca-branca e ao ácaro-rajado mediada por acilaçúcares e pelo gene Mi em tomateiro. Dissertation, Universidade Federal de Lavras
Nauen R, Denholm I (2005) Resistance of insect pest to neonicotinoid insecticides: current status and future prospects. Arch Insect Biochem Physiol. https://doi.org/10.1002/arch.20043
Nombela G, Beitia F, Muñiz M (2000) Variation in tomato host response to Bemisia tabaci (Hemiptera: Aleyrodidae) in relation to acyl sugar content and presence of the nematode and potato aphid resistance gene Mi. Bull Entomol Res. https://doi.org/10.1017/S0007485300000274
Nombela G, Williamson VM, Muniz M (2003) The root-knot nematode resistance gene Mi-1.2 of tomato is responsible for resistance against the whitefly Bemisia tabaci. Mol Plant-Microbe Interact. https://doi.org/10.1094/MPMI.2003.16.7.6450
Oliveira CM (2015) Efeito do gene Mi e dos altos teores foliares de acilaçúcares e de zingibereno na resistência do tomateiro a artrópodes-praga. Dissertation, Universidade Federal de Lavras
Petroski RJ, Stanley DW (2009) Natural compounds for pest and weed control. J Agric Food Chem. https://doi.org/10.1021/jf803828w
Prabhaker N, Castle S, Henneberry TJ, Toscano NC (2005) Assessment of cross-resistance potential to neonicotinoid insecticides in Bemisia tabaci (Hemiptera: Aleyrodidae). Bull Entomol Res. https://doi.org/10.1079/BER2005385
Resende JTV, Maluf WR, Faria MV, Pfann AZ, Nascimento IR (2006) Acylsugars in tomato leaflets confer resistance to the South American tomato pinworm, Tuta absoluta Meyr. Sci Agricola. https://doi.org/10.1590/S0103-90162006000100004
Resende JTV, Maluf WR, Cardoso MG, Faria MV, Gonçalves LD, Nascimento IR (2008) Resistance of tomato genotypes with high level of acylsugars to Tetranychus evansi Backer & Pritchard. Sci Agricola. https://doi.org/10.1590/S0103-90162008000100005
Resende JTV, Maluf WR, Cardoso MG, Gonçalves LD, Faria MV, Nascimento IR (2009) Resistance of tomato genotypes to the silver leaf whitefly mediated by acylsugars. Hortic Bras 27:345–348. https://doi.org/10.1590/S0102-05362009000300015
Roberts PA, Thomason IJ (1989) A review of variability in four Meloidogyne spp. measured by reproduction on several hosts including Lycopersicon. In: Russel GE (ed) Agricultural zoology reviews, vol 3. Intercept, Andover, pp 225–252
Rossi M, Goggin FL, Milligan SB, Kaloshian I, Ullman DE, Williamson VM (1998) The nematode resistance gene Mi of tomato confers resistance against the potato aphid. Proc Natl Acad Sci. https://doi.org/10.1073/pnas.95.17.9750
Silva AA (2012) Resistência de genótipos de tomateiro com teores foliares contrastantes de aleloquímicos a Liriomyza trifolii (Burgess) (Diptera: Agromyzidae) e a Myzus persicae (Sulzer) (Hemiptera: Aphididae). Dissertation, Universidade Federal de Lavras
Silva VF, Maluf WR, Cardoso MG, Gonçalves Neto AC, Maciel GM, Nízio DAC, Silva VA (2009a) Resistência mediada por aleloquímicos de genótipos de tomateiro à mosca-branca e ao ácaro-rajado. Pesqui Agropecu Bras. https://doi.org/10.1590/S0100-204X2009001000008
Silva LD, Omoto C, Bleicher E, Dourado PM (2009b) Monitoramento da suscetibilidade a inseticidas em populações de Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) no Brasil. Neotrop Entomol. https://doi.org/10.1590/S1519-566X2009000100013
Silva AA, Maluf WR, Moraes JC, Alvarenga R, Costa EMR (2013) Resistência a Myzus persicae em genótipos de tomateiro com altos teores foliares de aleloquímicos. Bragantia. https://doi.org/10.1590/S0006-87052013005000022
Tavares CAM (2002) Perspectivas econômicas da tomaticultura frente aos problemas causados pelo geminivirus. Biológico 64: 157–158. http://www.biologico.sp.gov.br/uploads/docs/bio/v64_2/Tavares.pdf. Accessed 18 April 2016
Villas Boas GL, Franca FH, Macedo N (2002) Potencial biótico da mosca-branca Bemisia argentifolii a diferentes plantas hospedeiras. Hortic Bras. https://doi.org/10.1590/S0102-05362002000100014
Weston PA, Snyder JC (1990) Thumbtack bioassay: a quick method of measuring plant resistance to two spotted spider mites (Acari: Tetranychidae). J Econ Entomol. https://doi.org/10.1093/jee/83.2.500
Acknowledgements
Our thanks to the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), to the Fundação de Amparo a Pesquisa do Estado de Minas Gerais (FAPEMIG), to the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and to Hortiagro Sementes S.A., for financial support, scholarships, and providing infrastructure.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Gouveia, B.T., de Oliveira, A.M.S., Ribeiro, G.H.M.R. et al. Resistance to whitefly (Bemisia argentifolii) and repellency to the two-spotted spider mite (Tetranychus urticae) in tomato plant hybrids with high leaf contents of acylsugar and the Mi gene. Euphytica 214, 140 (2018). https://doi.org/10.1007/s10681-018-2224-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10681-018-2224-1