
Abstract
Ricinus communis, also known as castor bean or castor plant, is a crop that produces seeds containing non-edible oil used widely in the chemical industry and the bioenergy sector. Varieties or hybrids that are suitable for mechanized harvesting may diminish the production cost boosting large scale production of the crop in new areas. In this study, we describe extensively the morphological characteristics and the ex situ performance of four newly developed R. communis hybrids (H11, H12, H13, and H14) focusing on plant architecture in order to facilitate mechanized harvesting. In addition, hybrid germination rate at low and optimal temperature was evaluated aiming to determine a minimal temperature requirement for early spring sowing in temperate climates. Seed metabolic profiling using LC–MS/MS analysis revealed the presence of more than 60 different primary and secondary metabolites, at varied levels in each hybrid, including amino acids, fatty acids, flavonols, flavones, flavanones, phenyl propanoids and catecholamines such as dopamine, many of which are reported for the first time in this species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Ricinus communis, a member of the Euphorbiaceae family, is an industrial crop producing seeds with high quality non-edible oil used in a plethora of industrial fields including biofuels, lubricants, paints, pharmaceuticals, cosmetics and textile finishing. The plant is commonly known as castor bean; although its seeds possess the shape and the size of a bean, the plant is not a legume, therefore the seeds are not true beans. The seeds are decorated with unique golden mottled designs on a dark burgundy background. Recently, it has been proposed the term “castor bean” to be replaced by the term “castor plant/seeds” (Milani and Medeiros Nobrega 2013). R. communis has a polyphyletic origin (Milani and Medeiros Nobrega 2013) with four claiming centers, all located in arid and semi-arid regions (Ethiopia, India, China and Arabia) (Anjani 2012). Further, it appears to possess a wide ability for adaptation in diverse temperate climates (Goyal et al. 2014).
Seed yield is a quantitative trait, which is the result of the genetic makeup affected by environmental conditions. Traditionally, seed selection of this species has focused on the improvement of seed yield and oil content rather than plant architecture. Nevertheless, efforts to develop mechanized harvesting of the castor seeds date back to the late 1940s in California (Knowles 1949). Modern breeding targets aim to: (i) alter plant architecture (Baldanzi et al. 2003), (ii) produce erect plants with long primary raceme, (iii) produce non-shattering fruits that mature early (biological cycle of less than 150 days), (iv) increase resistance to biotic and abiotic factors, and (v) reduce ricin content (Milani and Medeiros Nobrega 2013). Under normal conditions, the plant is monoecious with the female (pistillate) flowers positioned on the upper part of the raceme and the male (staminate) flowers on the lower part. The proportion of pistillate to staminate flowers can vary considerably within and among genotypes and it is strongly influenced by the environment (Milani and Medeiros Nobrega 2013) so as in extreme conditions dioecism occurs allowing the production of pistillate lines. The availability of pistillate lines in R. communis enabled the breeders to exploit heterosis (hybrid vigor) followed by selection (Severino et al. 2012). Evidence for hybrid vigor in seed led to the development of the first hybrids with high oil content (Lavanya and Chandramohan 2003) and disease resistance to Fusarium oxysporum (Patel and Pathak 2011) in India.
Capsule collection by the cumbersome and expensive hand-picking process is a common practice nowadays. As biofuel production reaches a commercial level, a significant shift in breeding targets has been achieved: the generation of plant lines and hybrids suitable for mechanized harvesting that is expected to reduce collection cost and contribute to the development of a competitive product with significant advantages over other fuel sources in the free market. Local liquid biofuel production from plant feedstocks can potentially reduce greenhouse gas emissions, boost rural economies, and minimize dependence of national economies on foreign fossil fuels (Bartley and Ronald 2009; Kapazoglou et al. 2013; Somerville 2007; Vega-Sanchez and Ronald 2010).
Seeds consist of the embryo and the endosperm tissue, which is filled with reserves of stored metabolites mainly oils, carbohydrates and proteins. These compounds are synthesized during seed development and are crucial for seed germination and seedling growth. Metabolic profiling is a powerful tool for detecting changes in metabolite composition in different genotypes and environments. Such an analytical approach could identify metabolites associated with processes and responses that are of agronomic importance. Only a few studies have characterized metabolites present in R. communis seeds (Campos et al. 2010; Maltman et al. 2007, 2002; Nogueira FbCS et al. 2012). Knowledge on metabolite content can be used for breeding purposes, for example in the development of genotypes that exhibit the appropriate metabolic profiles needed for industrial exploitation.
The draft genome sequence of castor (4.6-fold coverage), the first for a member of the Euphorbiaceae family, has already been reported (Chan et al. 2010), and the role of conventional and biotechnological approaches in genetic improvement of R. communis has been recently reviewed (Singh et al. 2015).
In the present study, we evaluated the performance and agronomic characteristics of four novel hybrids developed for mechanized harvesting and grown in different locations in Greece. Our comprehensive analyses on quantitative and qualitative traits provide evidence that hybrids differ significantly from crop wild relatives regarding plant architecture, germination dynamics, vigor, synchronous ripening and capsule shattering resistance. The high-throughput metabolomic analyses obtained by LC–MS/MS revealed differences in the nature and abundance of mature seed metabolites, even within a group of closely-related individuals.
Materials and methods
Plant material, growth conditions and DNA isolation
F1 seeds of R. communis hybrids H11, H12, H13 and H14 were developed in Israel (Nof Ayalon, 31°52′N, 34°59′E) as part of a breeding program aiming to improve plant characteristics for large-scale mechanized harvesting. The performance of the newly developed hybrids was evaluated ex situ in initial field trials in northern Greece (Institute of Applied Biosciences, Thermi, Thessaloniki, 23°59′N, 40°31′E, and Krithia 22°59′E, 40°50′N) on a deep sandy-clay soil, which was slightly acidic (pH 5.0–6.5) and had good drainage. A complete randomized design was used. To prepare the soil for sowing, deep plowing was initially applied followed by fine tillage. At the developmental stage of four to six leaves, herbicide application was performed to control the growth of weeds. Morphological characteristics and other agronomic traits, including plant height (height from ground to the base of the primary raceme), stem architecture, germination and flowering time, length of primary raceme, and number of capsules per raceme, were recorded. In addition, the 1000-seed weight and the oil content were measured. Plant specimens were visualized using a high-performance stereomicroscope (Leica MZ10F).
ISSR-based genetic analysis
To genotype the four hybrids and to evaluate their genetic relationship, PCR reactions were performed using the following five Inter-Simple Sequence Repeats (ISSR) primers: UBC-827, 835, 841, 860, and 881. The PCR reactions were carried out in a final volume of 25 µL, containing 30 ng of DNA, 0.5 U Taq polymerase (Kapa Taq), 2.5 µL of 10X reaction buffer, 3.0 mM MgCl2, 0.2 mM of each dNTP and 0.8 µM primer (Life Technologies). DNA amplifications were performed in My Cycler of Bio-Rad thermo cycler, under the following conditions: initial denaturation at 94 °C for 3 min, followed by 40 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 20 s and extension at 72 °C for 40 s, final extension at 72 °C for 10 min. PCR products were resolved by electrophoresis in 1.5 % agarose-TAE gels stained with ethidium bromide. A 100 bp ladder was used to compare the sizes of the amplicons. To ensure reproducibility of the DNA profiles, PCR reactions were performed in triplicates. DNA band scoring was performed recording all the DNA products that had been produced by the PCR reactions. The DNA band records were then processed in a binary matrix, where values ‘1’ and ‘0’ indicated the presence or the absence of a PCR product, respectively. The matrix was analysed by the Genetic Analysis in Excell (GenAlEX) version 6.501 software (Peakall and Smouse 2012). Subsequently, the MEGA5 software (Tamura et al. 2011) was used for the construction of the dendrogram from an UPGMA cluster analysis of the ISSR results.
Germination under different temperatures
Seeds of the four hybrids were sown in pots containing potting mix. The containers were covered with transparent covers and were regularly watered. Two temperature regimes (16 or 24 °C) were tested in a growth chamber (16/8 light/dark cycle; relative humidity 60 %) for the efficiency of germination in white light. More than 60 seeds were shown per treatment in three replicates. Germination observations were taken for 28 days post sowing.
LC-MC/MS profiles of metabolites in R. communis seeds
Castor seed oil was extracted by cold pressing using an automatic spiral oil expeller machine (BT Biopresser, Denmark).
Wild type and hybrid seeds originating from the same location were ground in liquid nitrogen and stored at −80 °C. Fifty milligram of mature seed tissue were extracted in 200 μl of 80 % methanol (High Performance Liquid Chromatography grade) and then vortexed for about 20 s. Subsequently, the samples were centrifuged at 11,000 g for 15 min at 4 °C to remove protein pellets. The cleared supernatant containing extracted metabolites was transferred into aliquots for liquid chromatography coupled with tandem mass spectrometry (LC–MS/MS) analyses. Metabolites were identified by triplicate analysis of single seed extracts.
Results and discussion
Phylogenetic relationships of the hybrids and the wild-types
The use of ISSR molecular markers to evaluate the genetic diversity offers a non-radioactive method that is easy, fast and with low cost to apply leading to reproducible results (Reddy et al. 2002) that are used to differentiate between related genotypes in many species. In the case of R. communis, ISSRs have been used recently to assess the genetic diversity of genotypes from India (Gajera et al. 2010; Kulshrestha and Talati 2014; Tomar Rukam et al. 2014), China (Wang et al. 2013) and other countries (Kallamadi et al. 2015). In all cases, dendrograms have been produced revealing the genetic closeness of the selected genotypes.
In the current study, molecular phylogeny was used to infer genetic relationships among a wild-type from Israel (105 N), four hybrid (H11, H12, H13, and H14- genotypes bred in Israel for traits suitable for mechanized harvesting), and also six wild-type genotypes (Amorgos, Fotolivos, Karatassou, Lefkada, Mikrokampos, Sitagroi) from Greece, using five ISSRs molecular markers that had been chosen from the University of British Colum-bia collection. The application of these markers resulted in the generation of multiple banding profiles. The number of bands that had been produced per ISSR was ranged from 12 (ISSR primer UBC881) to 17 bands (ISSR primer UBC841), with a mean of 14.8. Totally, 74 DNA bands were amplified. In more details, the UBC-827 produced 16 products in total, all within the range of 550–3300 bp. Similarly, UBC-835 produced 14 products of 350–2950 bp, UBC-841 17 products of 300–2800 bp, UBC-860 15 products of 640–2500 bp, and UBC-881 12 products of 700–3150 bp. The results of the ISSR analysis were used to construct a dendrogram visualizing the genetic closeness between the 11 genotypes that have been analyzed. Two distinctive clades were formed (Fig. 1a); four of the Greek genotypes (Mikrocampos, Fotolivos, Amorgos, and Lefkada) were classified in one clade, whereas the other clade consisted of the four hybrids plus the remaining two Greek genotypes (Sitagroi and Karatassou) plus the wild Israeli genotype 105 N (Fig. 1b). The phylogenetic data indicate that the Israeli genotypes (the four hybrids and the wild-type 105 N) shared high degree of homology for the ISSR loci, revealing a narrow genetic background which is in agreement with their breeding history. The majority (four out of six) of the wild-type Greek genotypes that had been used as references, were not grouped together with the four hybrids. Having, however, two Greek genotypes in the same clade together with the five Israeli genotypes indicates the occurrence of a genetic link between the genotypes of the two countries, both located at the eastern part of the Mediterranean Sea.

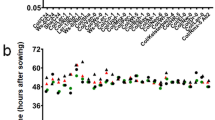
Genetic relatedness of R. communis hybrids and germination responses. a Phylogenetic analysis involving the four newly developed hybrids (H11, H12, H13, and H14), an Israeli wild-type genotype (105 N), and six Greek wild-type genotypes (Amorgos, Fotolivos, Karatassou, Lefkada, Mikrokampos, Sitagroi) that were used as references. The dendrogram is obtained from UPGMA cluster analysis of the ISSR-based results. b Leaf morphology of the six Greek wild-type genotypes used-detail of the glandular structure at the point where the petiole joins the lamella on the abaxial site. c Germination dynamics at 16 °C (circles) and 24 °C (squares). Seeds from hybrids H11, H12, H13 and H14 were evaluated for synchrony and efficiency of germination for 28 days after sowing
Temperature-dependent germination dynamics differ in hybrids
Early spring germination can be critical for the development of R. communis as commercial crop (Severino and Auld 2013). Successful early and rapid growth of the plants to escape the summer drought conditions may advance harvesting time thus reducing the risk of damage related to fall diseases and frost which may potentially threaten yield. Therefore, the effect of temperature on plant seed germination was investigated comprehensively in order to uncover possible genetic variation which could be useful for selecting the best genotypes for consistent performance. The hypothesis that R. communis seeds need a minimum of 16 °C soil temperature for germination was tested in all four hybrids, as well as to the optimal germination temperature of 24 °C (Fig. 1c). The results of this study showed that a minimum of eight days was required before the first seedlings of each hybrid germinated. All four hybrids performed better at 24 °C reaching 100 % germination either within 22 (H11 and H12) or 28 days (H13 and H14) post sowing. H13 performed exceedingly well at 24 °C as evidenced by nearly 80 % seed germination within one week post sowing. On the other hand, seeds of H11 and H12 needed an extra week to reach the same germination level. The germination pattern changed at 16 °C as the percentage of germinating seeds was severely reduced. Specifically, the highest final germination rate (80 %) was recorded for H11, while H12, H13 and H14 displayed lower rates (50–60 %). Germination of H12 and H14 seemed to be more tolerant to temperature since these two hybrids performed similarly at 16 and 24 °C during the first three weeks post sowing. Overall, the low temperature of 16 °C significantly impaired the potential of the hybrids to germinate whereas the temperature of 24 °C led to 100 % germination within 3–4 weeks after sowing. Our results are in agreement with previous studies, as reviewed recently (Severino and Auld 2013), correlating germination of R. communis seeds with temperature. Our results suggest that seed emergence under low temperature is a genotype-dependent response as the four hybrids responded differently regardless of their genetic closeness.
Ex-situ morphological characterization of plant architecture
Ex-situ phenotypic characterization of the novel hybrids in different environments in Greece was important to determine the genotype–environment interactions and the suitability of the hybrids for these environments. Macroscopic observations showed that all four hybrids possess seeds similar in shape and size (Fig. 2a). However, variations in the size of the protruding structure in seeds, named caruncle, were observed. It is believed that caruncle plays a role in seed dispersal by insects (Severino and Auld 2013). This structure is larger in H11 and H13 compared to that of H12 and H14 (Fig. 2b). At the early stages of development, when the seedlings emerged from the soil, the hypocotyl hooks in hybrids H12, H13 and H14 exhibited an anthocyanin shade of red (Fig. 2i). The anthocyanin pigmentation of stem is frequently observed in R. communis inbred lines (Baldanzi et al. 2015). Later in development, hybrid H11 developed solid green stems, whereas the remaining three hybrids possessed hollow stems with a reddish-green tinge (Fig. 2h). Interestingly, only hybrid H11 was sensitive to stem lodging. The greater bending strengths of the other three hybrids with hollow stem suggest that the hollowness of the stem is one genotype-related characteristic that must be taken into account during selection of varieties or hybrids with stem bending resistance. Therefore, breeding efforts should be channeled towards increasing stem hollowness to avoid stem lodging.

Comparison of the morphological features of the 4 R. communis hybrids. a Seeds of the hybrids. Left, side with the hilum; bars, 4 mm. b The caruncle. c Mature seed-bearing inflorescences. d Capsules. e Adaxial surface of the leaves. f Abaxial surface of the leaves. g Glandular structures at the junction of the petiole to the lamina. h Cross section of the stems. i Differences in hypocotyl color among genotypes during germination. Lower row H11; Upper row H12. H13 and H14 are similar to H12 (not shown). j Droplets of clear exudates on glandular structures of the petiole. k Exudates on abaxial side of the leaf. l Exudates from the hydathode-like structures. m Trichome morphology on the abaxial side of the leaves. n Seed oil color differentiation among the H11, H12, H13 and H14 hybrids
The pattern of plant architecture of castor hybrids H11 and H12 was characterized by two secondary branches, whereas the pattern of plant architecture in hybrids H13 and H14 was characterized by four to five secondary branches (Table 1). In nature, R. communis is highly branched. Unlike the R. communis plants found in nature, the fully grown hybrids of the current study are short (about 1–1.5 m) (Fig. 3a), in contrast to the R. communis plants in nature which reach 3–4 m in height. The dwarf hybrids of the current study exhibit a branched architecture of two (Fig. 3b, c) to five secondary stems (Fig. 3d). In this architecture, the primary raceme bears the main yield and the secondary branches bear smaller racemes that supply the pollen for the delayed flowers of the primary raceme. Importantly, the main stem bearing the primary raceme is within the same zone with the secondary racemes facilitating concentrated mechanized harvesting. Breeding efforts for the generation of R. communis varieties suitable for mechanized harvesting focused on the development of hybrids with only pistillate flowers across the raceme, which has greater number of capsules and consequently higher oil yield over their inbred parents. In general, branching in plants is influenced by genetic and environmental factors (Baldanzi and Pugliesi 1998). The wild-types of R. communis are usually multi-branched, with a main stem bearing a raceme usually at high level from ground while secondary branches bearing secondary racemes develop over time making harvesting laborious. ‘Plant ideotype for mechanized crop harvesting includes short stature, determinate growth habit with reduced number of racemes per plant, dense racemes and indehiscent capsules that are easy to hull’ (Fernandez-Martinez and Velasco 1998). Indeed, evaluation of five Italic and 19 Greek cultivars reported a mean of 1.3 (Laureti et al. 1998), and 1.9–4.0 (Koutroubas et al. 2000) racemes per plant, whereas one or two racemes per plant was suggested by Filho (Savy Filho et al. 2007) as optimal for mechanical harvesting. Among the four hybrids, H11 possessed the longest pedicel in contrast to H14 that possessed the shortest.

Plant height and architecture of H11, H12, H13 and H14 hybrids improved for mechanized harvesting. a plant height of up to 150 cm; b graphical representation of plant architecture—a primary raceme (about 18 cm from ground level) and four secondary racemes; c plant architecture consisting of a primary raceme and 2 secondary ones; d plant architecture consisting of a primary raceme and four to five secondary ones
In general, at the stage of maturity the height of the plants ranged between 101 and 125 cm, with the length of the primary racemes ranging between 45 and 70 cm (Table 1). Notably, H14 produced more capsules compared to the other hybrids but at the expense of seed weight (Table 1). All four hybrids possessed large alternately arranged palmate leaves with nine lobes and finely serrated margins (Fig. 2e, f); with the exception of H11, their petiole was reddish. Depending on the genotype, one or two glandular structures were visible on the underside of the leaf at the lamina–petiole junction (Fig. 2g). Spontaneous exudation from structures present on the petioles close to the stem junction was observed under high humidity conditions while further stereoscopic examination revealed the presence of clear gluey exudates on glands (Fig. 2j). Glandular structures with spontaneous clear exudates were also observed on the abaxial surface of the lamina close to the margins (Fig. 2k) and also on the serrated periphery of the lamina in structures that seem similar to hydathodes (Fig. 2l). The role of these exudates in the physiology of the plant is not well understood. Stereoscopic observation also revealed the presence of spiky trichomes (Fig. 2m) on the lamina’s abaxial surface.
Notably, distinct degrees of disease susceptibility among the genotypes were observed in the different locations in late fall so far. The disease severity was greater on short flower pedicel genotypes than on those with longer pedicels. It is likely that the compact shape of the raceme in combination with high density predisposed it to pathogen attack in high humidity conditions. The disease development in the Krithia location (north of Thessaloniki, Greece) was greater than in Thermi (east of Thessaloniki, Greece), presumably due to the rainy season and the higher plant density (70 × 30 vs 75 × 43 cm) that generated a disease favoring micro-climate. These results suggest that R. communis should not be planted in high densities. In general, high density in raceme may favour infection by bacteria in high humidity locations. On the contrary, a dense raceme in dry areas is less likely to be infected by diseases.
Field observations showed earliness in flowering for H11 and H14 at high rate (86 %). Flowering in castor plant is promoted by longer photo-periods, which result in higher yield of the primary raceme (Kumar et al. 1997). All genotypes developed capsules with spines. Capsule shatter at maturity can lead to considerable yield loss of about 20–50 % (field observations) prior to harvest. This character is critical for mechanized harvesting and the hybrids under investigation were genetically improved for shatter resistance. In the field trials, the hybrids showed very good resistance to capsule shattering. Regarding seed weight, the 1000-seed weight was determined for the four hybrids: H14 was found to be lighter (239.9 g) than the remaining three hybrids (H11: 290.6 g, H12: 300.1 g, H13: 290.4 g) (Table 1). Seed oil content (w/w) in hybrids ranged between 49–53 % (Table 1). Oil produced by the hybrids had distinct color as it can be seen in Fig. 2n.
Metabolite screening in mature R. communis seeds
Mature seeds of the four hybrids and also from a wild-type Greek genotype (Mikrocampos, hereafter Mkp) were analyzed for the content of extractable metabolites using LC–MS/MS analyses. A total of 65 metabolites were identified covering most of the primary metabolism (lipids, amino acids, hormones) and some of the secondary metabolism (phenolics, alkaloids, sterols, catecholamines and lignins). The top 20 metabolites were present in all genetic backgrounds and exhibited high abundance variation (Fig. 4). Out of a total of 65 metabolites, 35 were organic acids. One of those, namely ricinoleic acid, is the major (>90 %) lipid of the total fatty acids in R. communis and was particularly abundant in the hybrids compared to the Mkp wild-type genotype. In contrast, the hybrids appeared to have lower levels of the amino acids proline and tyrosine. Particularly, proline displayed 100-fold change between the wild-type genotype (Mkp) and the four hybrids (Fig. 4). Tyrosine, in turn, had the highest abundance in the wild-type genotype while it was undetectable in hybrid H11. Maslinic acid, a pentacyclic triterpene with antioxidant, anti-inflammatory and antiproliferative properties (Mokhtari et al. 2015) was detected only in the wild-type Mkp genotype.

Representative Ricinus metabolites identified in mature seeds based on LC–MS/MS data. Relative amounts of metabolites in mature seeds from different genetic backgrounds expressed as % of the highest amount, which was arbitrarily designated as ‘100’
The abundance of proline and tyrosine amino acid metabolites in certain genotypes suggests a dominant role of these amino acids in the regulation of R. communis seed metabolism. In plants, proline, a low molecular weight osmolyte that has been largely observed to accumulate under abiotic stress conditions is derived from nitrogen compounds such as glutamate and polyamines and can accumulate under diverse type of stresses such as oxidative stresses (Paschalidis et al. 2009; Skopelitis et al. 2006), water deficit (Paleg and Aspinall 1981; Paschalidis et al. 2010), tropospheric ozone (Goumenaki E, Karidis Z, Paschalidis K Assessment of Tropospheric Ozone Impact on Crops in Crete (Greece) Using Snap Bean as Bioindicator. In: XXVIII International Horticultural Congress on Science and Horticulture for People (IHC 2010), salinity (Ashraf and Harris 2004), low temperature (Naidu et al. 1991), high temperature (Kaushal et al. 2011), and heavy metals (Bassi and Sharma 1993a, b; Schat et al. 1997). Proline has a protective function against stress conditions acting as an osmolyte, radical scavenger, electron sink, stabilizer of macromolecules, and a cell wall component (Matysik et al. 2002). Therefore, a response to the aforementioned stresses may entail alterations in proline content which in turn may impact the ability of the plant to tolerate stress. Tyrosine was more abundant in H12 and Mkp compared to the other genotypes (Fig. 4). Tyrosine is not only converted into flavonoids in plants, but is also a precursor of dopamine. The physiological role of dopamine in plants is poorly understood. A brief overview of biosynthetic inter-relations between plant metabolites stemming from tyrosine has been recently reviewed (Trantas et al. 2015). Furthermore, hydroxytyrosol, a phenethyl alcohol, is presumably produced from tyrosine-derived dopamine (Alagna et al. 2012). Our study revealed the presence of dopamine in R. communis hybrid seeds, a finding that has never been reported previously. Dopamine was detected in all genotypes with H12 having the highest levels. However, the hydroxytyrosol content in H12 was moderate, suggesting that high tyrosine levels in this hybrid gave rise to dopamine but dopamine, in turn, was not subsequently converted to hydroxytyrosol. On the contrary, the wild-type high tyrosine content observed in our study reflected also high hydroxytyrosol, but low dopamine, contents. Surprisingly, H14 presented moderate levels of both tyrosine and dopamine levels, but the highest content of hydroxytyrosol, suggesting that both tyrosine and dopamine serve as main precursors of hydroxytyrosol in this seed. The remarkable compositional changes that were revealed by LC–MS/MS analysis advance our background knowledge on R. communis metabolic phenotypes in relation to their genetic makeup. Linking the metabolic changes to genomic changes would be valuable to R. communis breeding programs targeting different end products.
The phenolics identified in seeds classified into flavonols including quercetin and kaempferol, flavones including luteolin and apigenin which act as signaling molecules, and flavanons such as narigenin. Out of 15 detected secondary metabolites in R. communis hybrid seeds, 11 were flavonoids that could function as antioxidants. For instance, the antioxidant quercetin had the highest levels in H12. In the hybrid H11 many secondary metabolites displayed higher abundance compared to the other hybrids. For instance, the secondary metabolites cinnamic acid isomer, p-coumaric truxillic acid, caffeic acid, kampherol 3-O-glucoside, vanillin and apigenin 7-O-glucoside had higher levels in H11. Caffeic acid and its derivatives caffeic acid hexoside and caffeoyl hexose I, were present in all genotypes and particularly abundant in H11. Finally, only hybrids H12 and H13 were found enriched in some individual metabolites such as dopamine.
Our analyses identified numerous compounds of high value revealing the potential of R. communis hybrids to be used both for production of energy and high-value products. Previous metabolomic studies of R. communis seeds were mainly focused on oil-related metabolism. The acquisition of metabolite data from whole seeds expands our understanding of seed metabolism. The results from LC–MS/MS-based analysis provided evidence for distinct metabolical profiles of R. communis genotypes grown in the same area. The current LC–MS/MS study apart from detecting variations in known metabolites among hybrids, identified compounds not previously reported for this species. Our results on the relative amounts of metabolites may have implications for the genetic contribution to metabolite profiles. The phenolic compounds identified in hybrids were mainly involved in defense, amino acid, carbohydrate, and lipid metabolism during seed development. The detection of low-molecular weight phenolic acids, generated from cinnamic acid such as p-coumaric acid, ferulic acid, o-coumaric acids, syringic acid, in hybrid seeds is in agreement with a previous R. communis study (Chakravartula and Guttarla 2007).
Based on the initial information obtained by LC–MS/MS analysis, hybrid H11 had a five-fold increase in oleic acid content over the other genotypes. In the same hybrid, the ricinoleic acid content was 1.6 fold higher compared to the other genotypes. Thus, hybrid 11 may have evolved an oleic acid hydroxylase to synthesize ricinoleic acid, as it has also developed the capacity to efficiently accumulate high ricinoleic acid levels in its seed oil. The oleic acid hydroxylase gene, which produces ricinoleic acid, has evolved from the widely occurring FAD2 gene for the ∆12-oleic acid desaturase (Van De Loo et al. 1995). Among the key enzymes involved in the incorporation of ricinoleic acid into oils are diacylglycerol acyltransferases (DGATs), which catalyze the final step in triacylglycerol assembly (Chan et al. 2010). Castor oil for biodiesel production is characterized by high viscosity due to its high content in ricinoleic acid and low in oleic acid (Knothe 2008). As a result of this, castor oil for biofuel production must be blended with petrodiesel and not used in its pure form. An improvement of the oil viscosity for biofuel production can be the reduction of ricinoleic acid content with the simultaneous increase in oleic acid as it was found in a natural R. communis mutant (Rojas-Barros et al. 2004). Additionally, because of its high proportion of the unusual fatty acid ricinoleic acid, oil from R. communis could be used further as high-quality lubricant (Goodrum and Geller 2005), thus helping to replace sulfur lubricants in petroleum diesel and subsequentlyt reduce sulfur atmospheric emissions. Our results also reflect the modest production of various metabolic compounds that have been transiently expressed in seeds of R. communis hybrids. Oil composition is a complex seed quality trait functionally related to seed dormancy and seedling vigor (Snider et al. 2014). Because of the metabolic complexity in seeds, a synthetic approach to reproduce the oil composition seems rather a challenging task. Rather, screening of diverse genetic backgrounds by metabolic profiling is a forward approach that can serve as an important resource for the exploitation of seed-derived end products with great promise for the chemical and bioenergy industries. The natural variation in castor seed metabolites can direct future breeding strategies and metabolic engineering for crop improvement.
Conclusions
In the current study four R. communis hybrids improved for mechanized harvesting were characterized phenotypically, genetically and chemically. The hybrids possess short stature, with two or four to five secondary branches bearing racemes that ripe synchronously to the primary raceme. In the context of large-scale production with low cost, optimal plant architecture and robust performance are key determinants. The findings of this research also improve our understanding of the seed metabolomic response to genetic variation and can potentially broaden breeding strategies for crop improvement.
References
Alagna F, Mariotti R, Panara F, Caporali S, Urbani S, Veneziani G, Esposto S, Taticchi A, Rosati A, Rao R (2012) Olive phenolic compounds: metabolic and transcriptional profiling during fruit development. BMC Plant Biol 12(1):162
Anjani K (2012) Castor genetic resources: a primary gene pool for exploitation. Ind Crops Prod 35(1):1–14
Ashraf M, Harris PJC (2004) Potential biochemical indicators of salinity tolerance in plants. Plant Sci 166(1):3–16
Baldanzi M, Pugliesi C (1998) Selection for non-branching in castor Ricinus communis L. Plant Breed 117(4):392–394
Baldanzi M, Fambrini M, Pugliesi C (2003) Redesign of the castorbean plant body plan for optimal combine harvesting. Ann Appl Biol 142(3):299–306
Baldanzi M, Myczkowski ML, Salvini M, Macchia M (2015) Description of 90 inbred lines of castor plant (Ricinus communis L.). Euphytica 202(1):13–33
Bartley L, Ronald P (2009) Plant and microbial research seeks biofuel production from lignocellulose. Calif Agric 63(4):178–184
Bassi R, Sharma SS (1993a) Changes in proline content accompanying the uptake of zinc and copper by Lemna minor. Ann Bot 72(2):151–154
Bassi R, Sharma SS (1993b) Proline accumulation in wheat seedlings exposed to zinc and copper. Phytochemistry 33(6):1339–1342
Campos FAP, Nogueira F, Cardoso KC, Costa GCL, Del Bem LEV, Domont GB, Da Silva MJ, Moreira RC, Soares AA, Jucà TL (2010) Proteome analysis of castor bean seeds. Pure Appl Chem 82(1):259–267
Chakravartula SV, Guttarla N (2007) Identification and characterization of phenolic compounds in castor seed. Nat Prod Res 21(12):1073–1077
Chan AP, Crabtree J, Zhao Q, Lorenzi H, Orvis J, Puiu D, Melake-Berhan A, Jones KM, Redman J, Chen G (2010) Draft genome sequence of the oilseed species Ricinus communis. Nat Biotechnol 28(9):951–956
Fernandez-Martinez JM, Velasco L (1998) Castor. Technological innovations in major world oil crops, vol 1. Springer, New York, pp 237–265
Gajera BB, Kumar N, Singh AS, Punvar BS, Ravikiran R, Subhash N, Jadeja GC (2010) Assessment of genetic diversity in castor (Ricinus communis L.) using RAPD and ISSR markers. Ind Crops Prod 32(3):491–498
Goodrum JW, Geller DP (2005) Influence of fatty acid methyl esters from hydroxylated vegetable oils on diesel fuel lubricity. Bioresour Technol 96(7):851–855
Goumenaki E, Karidis Z, Paschalidis K (2010) Assessment of tropospheric ozone impact on crops in crete (Greece) using snap bean as bioindicator. In: XXVIII international horticultural congress on science and horticulture for people (IHC2010): International symposium on 938, pp 401–407
Goyal N, Pardha-Saradhi P, Sharma GP (2014) Can adaptive modulation of traits to urban environments facilitate Ricinus communis L. invasiveness? Environ Monit Assess 186(11):7941–7948
Kallamadi PR, Nadigatla VPRGR, Mulpuri S (2015) Molecular diversity in castor (Ricinus communis L.). Ind Crops Prod 66:271–281
Kapazoglou A, Drosou V, Nitsos CK, Bossis I, Tsaftaris A (2013) Biofuels get in the fast lane: developments in plant feedstock production and processing. Adv Crop Sci Technol 1:117
Kaushal N, Gupta K, Bhandhari K, Kumar S, Thakur P, Nayyar H (2011) Proline induces heat tolerance in chickpea (Cicer arietinum L.) plants by protecting vital enzymes of carbon and antioxidative metabolism. Physiol Mol Biol Plants 17(3):203–213
Knothe G (2008) “Designer” biodiesel: optimizing fatty ester composition to improve fuel properties. Energy Fuels 22:1358–1364
Knowles P (1949) Castor beans: studied for potential values as oil crop for California production. Calif Agric 3(3):13–14
Koutroubas SD, Papakosta DK, Doitsinis A (2000) Water requirements for castor oil crop. J Agron Crop Sci 184(1):33–41
Kulshrestha K, Talati JG (2014) Molecular characterization of castor (Ricinus communis L.) genotypes through SSR and ISSR markers. Indian J Agric Biochem 27(1):20–24
Kumar PV, Ramakrishna YS, Rao BVR, Victor US, Srivastava NN, Subba Rao AVM (1997) Influence of moisture, thermal and photoperiodic regimes on the productivity of castor beans (Ricinus communis L.). Agric Meteorol 88:279–289
Laureti D, Fedeli AM, Scarpa GM, Marras GF (1998) Performance of castor (Ricinus communis L.) cultivars in Italy. Ind Crops Prod 7(2):91–93
Lavanya C, Chandramohan Y (2003) Combining ability and heterosis for seed yield and yield components in castor. J Oilseeds Res 20:220–224
Maltman DJ, Simon WJ, Wheeler CH, Dunn MJ, Wait R, Slabas AR (2002) Proteomic analysis of the endoplasmic reticulum from developing and germinating seed of castor (Ricinus communis). Electrophoresis 23(4):626–639
Maltman DJ, Gadd SM, Simon WJ, Slabas AR (2007) Differential proteomic analysis of the endoplasmic reticulum from developing and germinating seeds of castor (Ricinus communis) identifies seed protein precursors as significant components of the endoplasmic reticulum. Proteomics 7(9):1513–1528
Matysik J, Bhalu B, Mohanty P (2002) Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Curr Sci 82(5):525–532
Milani M, Medeiros Nobrega MB (2013) Castor breeding. Agricultural and biological sciences. In: Andersen SB (ed) Plant breeding from laboratories to fields. doi:10.5772/56216
Mokhtari K, Rufino-Palomares EE, Reyes-Zurita FJ, Pérez-Jiménez A, Figuera C, García-Salguero L, Medina PP, Peragón J, Lupiáñez JA (2015) Maslinic acid, a triterpene from olive, affects the antioxidant and mitochondrial status of B16F10 cells grown under stressful conditions. Evid-Based Complem Altern Med
Naidu BP, Paleg LG, Aspinall D, Jennings AC, Jones GP (1991) Amino acid and glycine betaine accumulation in cold-stressed wheat seedlings. Phytochemistry 30(2):407–409
Nogueira FbCS, Palmisano G, Soares EL, Shah M, Soares AA, Roepstorff P, Campos FAP, Domont GB (2012) Proteomic profile of the nucellus of castor bean (Ricinus communis L.) seeds during development. J Proteom 75(6):1933–1939
Paleg LG, Aspinall D (1981) The physiology and biochemistry of drought resistance in plants. Academic Press
Paschalidis KA, Moschou PN, Toumi I, Roubelakis-Angelakis KA (2009) Polyamine anabolic/catabolic regulation along the woody grapevine plant axis. J Plant Physiol 166(14):1508–1519
Paschalidis KA, Imene T, Panagiotis MN, Roubelakis-Angelakis KA (2010) ABA-dependent amine oxidases-derived H2O2 affects stomata conductance. Plant signal behav 5(9):1153–1156
Patel P, Pathak H (2011) Genetics of resistance to wilt in castor caused by Fusarium Oxysporum F. Sp. Ricini Nanda and Prasad. Agric Sci Dig 31(1):30–34
Peakall R, Smouse PE (2012) GenAlEx 6.5: genetic analysis in excel. Population genetic software for teaching and research: an update. Bioinformatics 28(19):2537–2539
Reddy MP, Sarla N, Siddiq EA (2002) Inter simple sequence repeat (ISSR) polymorphism and its application in plant breeding. Euphytica 128(1):9–17
Rojas-Barros P, de Haro A, Muñoz J, Fernández-Martínez JM (2004) Isolation of a natural mutant in castor with high oleic/low ricinoleic acid content in the oil. Crop Sci 44(1):76–80
Savy Filho A, Amorim EP, Ramos NP, Martins ALM, Cavichioli JC (2007) IAC-2028: new castor bean cultivar. Pesqui Agropecu Bras 42(3):449–452
Schat H, Sharma SS, Vooijs R (1997) Heavy metalal-induced accumulation of free proline in a metal-tolerant and a nontolerant ecotype of Silene vulgaris. Physiol Plant 101(3):477–482
Severino LS, Auld DL (2013) A framework for the study of the growth and development of castor plant. Ind Crops Prod 46:25–38
Severino LS, Auld DL, Baldanzi M, Cândido MJ, Chen G, Crosby W, Tan D, He X, Lakshmamma P, Lavanya C (2012) A review on the challenges for increased production of castor. Agron j 104(4):853–880
Singh AS, Kumari S, Modi AR, Gajera BB, Narayanan S, Kumar N (2015) Role of conventional and biotechnological approaches in genetic improvement of castor (Ricinus communis L.). Ind Crops Prod 74:55–62
Skopelitis DS, Paranychianakis NV, Paschalidis KA, Pliakonis ED, Delis ID, Yakoumakis DI, Kouvarakis A, Papadakis AK, Stephanou EG, Roubelakis-Angelakis KA (2006) Abiotic stress generates ROS that signal expression of anionic glutamate dehydrogenases to form glutamate for proline synthesis in tobacco and grapevine. Plant Cell 18(10):2767–2781
Snider JL, Collins GD, Whitaker J, Chapman KD, Horn P, Grey TL (2014) Seed size and oil content are key determinants of seedling vigor in Gossypium hirsutum. J Cotton Sci 18:1–9
Somerville C (2007) Biofuels. Curr biol 17(4):R115–R119
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28(10):2731–2739
Tomar Rukam S, Parakhia MV, Kavani RH, Dobariya KL, Thakkar JR, Rathod VM, Dhingani RM, Golakiya BA (2014) Characterization of castor (Ricinus communis L.) genotypes using different markers. Res J Biotechnol 9(2):6–13
Trantas EA, Koffas MA, Xu P, Ververidis F (2015) When plants produce not enough or at all: metabolic engineering of flavonoids in microbial hosts. Front Plant Sci 6
Van De Loo FJ, Broun P, Turner S, Somerville C (1995) An oleate 12-hydroxylase from Ricinus communis L. is a fatty acyl desaturase homolog. Proc Natl Acad Sci 92(15):6743–6747
Vega-Sanchez ME, Ronald PC (2010) Genetic and biotechnological approaches for biofuel crop improvement. Curr Opin Biotechnol 21(2):218–224
Wang C, Li G-r, Zhang Z-y, Peng M, Y-s Shang, Luo R, Chen Y-s (2013) Genetic diversity of castor bean (Ricinus communis L.) in Northeast China revealed by ISSR markers. Biochem Syst Ecol 51:301–307
Acknowledgments
We gratefully acknowledge financial support of the JONAH-FUEL (3176) by Greece and the European Regional Development Fund within the program ‘Bilateral R&D cooperation between Greece and Israel 2013–2015’.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Merkouropoulos, G., Kapazoglou, A., Drosou, V. et al. Dwarf hybrids of the bioenergy crop Ricinus communis suitable for mechanized harvesting reveal differences in morpho-physiological characteristics and seed metabolic profiles. Euphytica 210, 207–219 (2016). https://doi.org/10.1007/s10681-016-1702-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-016-1702-6