
Abstract
Barley is one of the most important cereal crops grown for the livelihoods of the poor farmers of Tigray region in northern Ethiopia. As many low input and marginal environments it has benefited less from the yield increases achieved by modern breeding. This has been largely attributed due to genotype × environment intraction (GEI). To investigate the causes of GEI, ten barley varieties including local checks (two farmers developed varieties, four modern varieties and three rare local varieties) were tested over 21 environments. Participatory methods were applied to sample an adequate number of environments spanning the regional diversity. The yielding ability and stability of the varieties was graphically depicted by GGE and PLSR biplot. There were two major groups of environments, the central and northern highlands, the latter with less rainfall and poorer soils. Rainfall per month and total nitrogen level were the environmental variables that differentiated these two groups. In Tigray, rainfall in June and July were negatively correlated with yield, reflecting waterlogging problems. The different varieties were either specifically or widely adapted across the two environments. The variety ‘Himblil’, originating in Tigray, was the highest yielding and also most stable in the region of origin. However, it was inferior to improved varieties (Shege and Dimtu) at high yield levels. The association of earliness with grain yield indicates that the trait can be effectively manipulated within the existing materials. We recommend breeding for drought/water logging resistance based on selection in the target environment as the best strategy to provide stable and high yielding varieties for Tigray.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Barley is one of the most important cereal crops grown for the livelihoods of the poor farmers of Tigray region in northern Ethiopia. Its productivity is low, ranging from 0.8 to 1.8 t/ha, depending on the season and applied inputs (Fekadu and Skjelvag 2002). As many low input and marginal environments it has benefited less from the yield increases achieved by modern breeding. This has generally been ascribed to several reasons: (a) that the varietal development and testing has been done in conditions not representative of those of resource-poor farmers, implying a low or negative genetic correlation, (b) that breeding materials evaluated may lack genetic variance (c) a higher experimental inaccuracy leading to lower heritability in marginal conditions, and (d) stronger spatial and temporal fluctuations among such environments.
This case study—based on data from Tigray—addresses the general issue of varieties ˝specifically adapted˝ to low input target environments as advocated by Simmonds (1991). His term ‘local adaptation’ was critically defined by Atlin et al. (2001) to mean ‘ecologically specific adaptation’, implying that truly ‘local’ adaptation is rare, such as the case of photoperiod-sensitive ‘Rayada’ varieties of flooding rice in Bangladesh (Vaughan and Chang 1992). In contrast photoperiod insensitive rice will give a wide adaptation.
To delineate distinct groups of ‘target environments’ has, however, been an important issue in multi-environment trials (MET) research, and a prerequisite for any meaningful cultivar evaluation and recommendation (Yan et al. 2000). A significant GEI can be positively exploited to identify stable genotypes adapted for specific target areas (Ceccarelli 1996), or effectively removed—once the interacting factor is identified—e.g., through selection for resistance or low pH tolerance. In general exposing varieties to contrasting sets of environments (year × locations) is an effective way of identifying genotypes with high average yield and low GEI, but at the expense of the specific adaptation (Annicchiarico 2002; Holland et al. 2001; Ceccarelli 1989).
The analysis and interpretation of GEI range from simple analysis of variance to more specific analyses of genotype performance, from the univariate linear regression analysis of Finlay and Wilkinson (1963) and Eberhart and Russell (1966) or Perkins and Jinks (1968), to the multivariate AMMI (Zobel et al. 1988), GGE biplot (Yan 2001) and PLSR (Partial Least Squares Regression) models (Vargas et al. 1998). The commonly used linear regression analysis typically explains only a portion of the interactions. The AMMI model analysis is useful in displaying the main effects of genotypes and environments and their interactions. It can estimate the genotype responses and separate noise from real sources of variation through partitioning of the GEI. This model has contributed to improved cultivar evaluations, recommendations and selection of test sites (Gauch and Zobel 1997). Although the AMMI biplot can be very effective in summarizing the variation and in visualizing main effects, it does not show which variety is high yielding in which environment (Yan 2001), nor the discrimination and representativeness of the environments (Yan et al. 2000). Yan and Rajcan (2002) demonstrated the effectiveness of GGE biplot for trait × environment and factor × environment relationships in soybean. High yielding and stable varieties as well as representative and discriminating environments could be identified. In addition to GGE, PLSR has been used successfully by Aastveit and Martens (1986) and Vargas et al. (1998) in bilinear modeling seeking common structures (‘latent variables’) in both the of X matrix (agronomic variables, rainfall, temperature and soil factors) and the Y matrix (yield data in one or more environments).
The latter is very important, since there is usually not one single stress, and certain stresses like drought actually affect yield differently depending on the crop stage. This has led Ceccarelli et al. (1998) to propose that breeding for drought tolerance has to be based on direct selection for grain yield in the target environment, since drought-prone conditions, tend to be more different from each other than favorable environments (Ceccarelli and Grando 2007) and produce repeatable cross-over interactions (Ceccarelli 1996).
Factors affecting GEI include phenology (van Oosterom et al. 1993; Abay and Cahalan 1995), adaptation to cool temperatures (Annicchiarico et al. 2005), nutrient availability such as nitrogen (Atlin and Frey 1989; Górny 2001; Sinebo et al. 2002; Muruli and Paulsen 1981) and phosphorus (Atlin and Frey 1989; Ahokas and Manninen 2001).
Usually marginal farmers face complex biotic and abiotic stresses, and avoiding a crop failure in drought seasons (i.e., avoiding GEI) is more important than achieving maximum yield potential in good seasons. Moreover, the yield is not only grain, but also feed value of the straw and the quality attributes. Hence to fix desirable GEI effects may be particularly relevant to them The objective of this study was to investigate the reasons why the officially recommended released varieties ‘HB-42’ and ‘Shege’ have not been adopted by farmers in Tigray and to identify the traits required for adaptation in this part of Ethiopia.
We wished to investigate the causes of this and the traits required for adaptation in this part of Ethiopia. Our hypothesis was that specific adaptation is needed and a regional breeding “programme is required to produce varieties acceptable by farmers”. We employed participatory methods to sample an adequate number of environments spanning the regional diversity. The formal breeding often fails to realize such complexity, and for this reason Participatory Varietal Selection was investigated as a complementary strategy.
Materials and methods
Plant materials
A participatory search process for varieties that matched farmers’ criteria led to the selection of nine varieties (Table 1) including four improved varieties (IV), two farmers developed varieties and three local varieties. Of the four improved varieties ‘Dimtu’ and ‘Misrach’ are intended for both early and late sowings and for both high and low input conditions, while ‘HB-42’ and ‘Shege’ are released for high input, but deliberately included because of their official recommendation for cultivation in Tigray. ‘Himblil’ and ‘Demhay’ are selections from landraces developed by the farmer Kahsay Negash of Bolenta, who also hosted one of the experiments. Three other landraces, the medium to late maturing ‘Rie’, ‘Sihumay’ and ‘Atona’, were included on farmer request since they were becomingrare. For more detailed description of rare and farmers developed varieties see Abay et al. (2008). Local checks used were different across locations but similar across years two row barley varieties in the drier sites (Adinefas, Melfa, Habes FTC, Habes on farm, Mugulat, Buket, Neksege and Mekele) and six row varieties in the wetter (Menkere, Mekhan, Bolenta, Fala, Holleta and Debrebrhan). The research addresses only the highland barley growing areas of Tigray, not the most drought stressed zones, since the test materials were six rowed and appropriate to highlands with longer growth period. The location of the test sites (Fig. 1) and their characteristics are presented in Table 2. The experiments were conducted for two production main seasons in 2004 (seven locations) and 2005 (14 locations); they were unbalanced in terms of locations × years, but balanced for varieties × year and varieties × locations. In the second year three research stations were included: Mekelle in Tigray and Holleta and Debrebrhan in central and north Shewa region, respectively. These are research sites from where the IVs were released.

Map of Tigray and location of test sites
Decentralized participatory trials
These trials were conducted for 2 years in four villages of Bolenta, Habes, Buket and Mugulat. The villages were selected to represent existing biodiversity and constraints of the area, e.g., soil fertility, water logging and disease pressure, and participating farmers included experienced seed selectors and women. (Since 30% of the households are female headed, on average 20% of the experimenter farmers were women).
Experimental design
The trials followed the “mother and baby” design (Snapp 1999). In this paper only the data from the mother trials were considered for analysis. The mother trials in which two farmers in each village received the complete set of varieties were considered for analysis. A randomized complete block with two replications (farmers) was used. In all locations the soil was ploughed 2–3 times and seeds were planted manually in six rows 2.5 m long and 20 cm apart. A seed rate of 100 kg/ha was used uniformly. The trials were managed as per farmers’ practice; including soil preparation, weeding and no fertilizer was applied.
On station and farmers training center (FTC) trials
These trials were, respectively, managed by researchers and development agents. Three trials were conducted on station and ten trials in Tigray were conducted at FTCs. Trials on station received DAP fertilizer before planting at a rate of 100 and 50 kg/ha for urea. The planting dates ranged from 27 June to 6 July, in both years. The FTC and the trials on station trials were managed by the researchers, Development Agents (DAs) and technical staffs of the research sites.
Data collection and statistical analysis
Days to heading (DH) and maturity (DM) were recorded as the number of days from date of sowing to 50% appearance of and 75% discoloration of the spikes in a plot, respectively. Plant height (PH) was measured in cm from ground level to the base of the main spike at maturity. The central four rows (2 m2) were hand harvested and hand threshed, in order to record the grain yield (in grams and later converted into kg ha−1). Thousand grain weight (TGWT) was recorded in grams as the average of three samples of 1,000 grains from a plot after the harvest. Rainfall, minimum and maximum temperatures were recorded from meteorological stations. Data were recorded at the site in the case of Neksege, Menkere, Mekele, Holleta and Debrebrhan while for the others data were obtained from meteorological stations located in the closest district headquarters.
Quantitative and qualitative data were recorded from mother trials. Mutual visits within villages were organized for participatory evaluation at three stages of crop growth (vegetative, flowering and grain filling). Household preferences were assessed based on household level questionnaires (HLQ), their matrix ranking level of introduced variety. The reasons for preference of a given variety were recorded to be considered as criteria for ranking Following the methods explained by Christinck et al. (2005), the farmers’ preference criteria and varieties were listed in a matrix to understand the choices between the varieties.
Analysis of variance for the 14 locations in 2005 was performed as for the single-year-multi-location trial (SYMLT) using the GLM procedures in SAS version 9.2 (SAS 2001) and for AMMI using IRRISTAT version 4.3 software (IRRI 2002). Coefficients of variation (CV) were calculated to measure the precision of experiments and the mean standard error differences to compare the differences between varieties. For joint regression analysis, the combination of years and locations was considered as 21 environments (MET) and partitioned into components of linearity and deviation from regression. Pooled error was obtained from the error means square divided by replications. The relative performance of varieties across environments was further examined through stability analysis. The analysis was performed using SASG × ESTAB: a SAS program developed by Hussein et al. (2000). GGE biplot analysis was performed to graphically display the relations among genotypes, environments and their interactions (Yan 2001). The underlying influence of the year variation was explored further by visualizing the ‘‘which won where’’ feature in the GGE biplots (Yan et al. 2000), to visualize target environments sharing the same winner genotype.
Environmental variables were used to characterise the main effects and their interactions. The data obtained from these variables were used as input for both GGE and PLSR models (Martens and Naes 1989).
GGE biplot analysis
GGE biplot provides a capacity to evaluate cultivars for their yielding ability and stability. As described by Yan and Tinker (2006), in a GGE biplot based on genotype-focused singular value partitioning, the length of the genotype vector measures its contribution to either G or GEI or both. Genotypes located near the origin have little contribution to GEI. In the ‘‘polygon’’ view of the GGE biplot, the ‘vertex’ genotypes are the most responsive genotypes, because they have the longest distance from the origin in their directions (Yan and Tinker 2006). Depending on the length and direction of the vector, both good and poor performing genotypes can be considered as responsive. In a GGE biplot based on environment-focused singular value partitioning, the correlation (grouping) of two environments in their ranking of the genotypes can be visualized by the angle between them. An angle of 90° indicates independence, an angle >90° a negative correlation and an angle <90 a positive correlatio The GGE biplot analysis was performed for genotype × environment, factor × environment and trait × environment interactions (Yan 2001; Yan and Rajcan 2002). PLSR was applied in order to visualize the relationship of yield with external covariables of soil and rainfall with full cross-validation to test significance of traits associated with PLS factors using the Unscrambler version 9.6. (camo@camo.no).
Results
Rainfall data
The rainfall amount and distribution differed clearly between the central sites Holleta/Debrebrhan and Tigray (Fig. 2). The main season started in May in Holleta and lasted until September/October, whereas the locations in Tigray had highest rainfall (around 54–70% of the total) in July and August, which corresponded to the sowing and vegetative stages of the crop. A lack of rain in May is crucial for timely sowing of six rowed varieties, while deficient rainfall in September may subject the crop to drought stress during the grain filling stage (Fig. 2). The coefficients of variation across locations were also markedly higher in Tigray than in the central locations. Moreover, the years were contrasting, 2004 being dry (especially in May and September) and 2005 unusually wet especially during grain filling. Rainfall varied more than twofold between locations in Tigray in 2005 (the driest location Neksege had only 330 mm in 2004). In 2004 the Atsbi site failed completely. The average grain yield ranged from a minimum of 180 kg/ha in Neksege to a maximum 2,428 kg/ha in Holetta. As shown in Figs. 3 and 4, yield was not determined by rainfall alone. Although the rainfall at Debrebrhan was higher than Holleta, the latter gave 63% higher yields. In both years rainfall at Menkere was about twice as much as at Neksege—the driest sites. However, the yield at Menkere was only 53 and 65% of that at Bolenta and Mugulat, respectively. This implies that there are other factors that determines yield in addition to rainfall as shown by Ceccarelli et al. (2007) in Mediterranean environments.

Means and CVs of monthly rainfall of experimental sites

Total rainfall, average grain yield and coefficient of variation by test environments, HO Holleta; DB Debrebrhan; HB Habes; HBF Habes FTC; BU Buket; MG Mugulat; BO Bolenta; MH Mekhan; MR Menkere; NK Neksege; FA FALA; Ad Adinefas; MF Melfa; 1 and 2 stands for the experimental years of 2004 and 2005, respectively

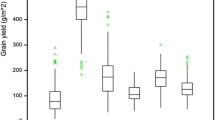
Grain yields of high and low input trials by variety
Analysis of soil fertility
Sites varied in organic matter (OM%)”, total nitrogen (%) and soil texture and pH value (Table 2). Using the rating of Mitiku et al. (2003), where >0.2% means low, 0.1–0.2% means very low and <0.1% extremely low N, 90% of the experimental plots had extremely low, while 10% (notably Holleta and Debrebrhan) had low N content. The OM was also higher in the research stations, but less than on the well manured farm in Bolenta. The low pH in the central highlands was striking, approaching the tolerance level of barley, whereas it was rather high in Tigray.
Analysis of variance
Significant components of genotypic, environment, and GEI for grain yield were revealed for all trials (Table 3). Mean grain yield varied among trials and ranged from 1,052 kg/ha for SYMLT and 915 kg/ha for MET (Table 4). The lower yields of the latter trials shows the large differences between years (an average yield advantage of 265 kg/ha in 2005 over 2004). Analysis of SYMLT revealed that location effects were 3.35 and 20.5 larger than genotype and interaction effects, respectively. The ANOVA for AMMI analysis for the yield of ten varieties evaluated in 21 environments also indicated that Environments (E), Genotypes (G) and GEI were highly significant and accounted for 60, 23 and 17% of the total sum squares (SS), respectively.
The over all mean yields (Table 4) indicated highly significant differences between the genotypes. The coefficients of variation (CV) ranged from 6 to 16% indicating the consistency of the experiments over the 2 years and locations. ‘Himblil’ was the best variety in Tigray and significantly different from other varieties, not only in the high yielding environments of Bolenta and Habes, but also at the driest site (Neksege) and in the water logged soils of Menkere. ‘Misrach’ was superior to the two-rowed checks at some dry sites, but not to the six-rowed in the wetter sites. ‘Dimtu’ was either inferior or not different from the local checks. ‘HB-42’ and ‘Shege’ were consistently the lowest yielding at all sites (Table 4).
The varieties ranked differently at different yield levels (Fig. 5). The average genotypic correlation of yields of varieties between groups was negative (r = − 0.21). Within Tigray the baby trials were highest yielding, followed by on-farm and last the FTCs (970, 889 vs. 710 kg/ha). This was consistent over all the varieties and was probably due to the fact that the demonstration plots were located on poorly managed soils, too poor to be representative of farmers’ management conditions.

Regression of mean grain yield (kg/ha) of ten varieties on the mean grain yield of on station and on farm trials 2004–2005
Stability analysis
In the Eberhart and Russell (1966) joint regression analysis, both linear and non linear components of the GEI were significant (Table 3). 50% of the GEI sum of squares was accounted for by the deviation from regression and 50% was accounted for by the linear regression on the means of environment. This suggests that both components were equally important for describing stability. However, the higher efficiency of AMMI in partitioning SS of GE interaction was demonstrated, since the first principal component (IPC1) alone accounted for 53%. Further partitioning of the GEI showed significant heterogeneity among slopes of regressions and residual non linear component.
There was clear evidence of cross-over interactions. Given the analytical limitations of linear interactions (Fig. 5; Table 4), ‘Himblil’ yielded more than the other varieties, when yield levels were less than about 1,200 kg ha−1, and had a βi < 1 typical of a variety adapted to unfavorable conditions. It was followed by ‘Local’, ‘Misrach’ and ‘Dimtu’ with βi-estimates close to one, indication a wide adaptation across environments. The varieties ‘Shege’, ‘Sihumay’, ‘Rie’ and HB42 have the opposite behavior being adapted to high input conditions.
The biplot divided the locations in two sectors (Fig. 6). The first sector contains the high input on station trials of Holetta and Debrebrhan and the varieties ‘Shege, ‘Dimtu’ and ‘Misrach’. ‘Misrach’ was the ‘winner’ at Debrebrhan, ‘Shege’ and ‘Dimtu’ at Holleta, coinciding with their selection environments. The second sector includes the low input trials and FTCs of Tigray locations with cultivar ‘Himblil’ being a winner in 19 location × year combinations. In conclusion the biplot shows that (1) the IVs were associated with the relatively high yielding environments, both outside Tigray, (2) ‘Himblil’ was the best variety in both years in Tigray locations, but among the poorest yielding at Debrebrhan and Holleta (3) ‘HB-42’ had the opposite pattern. The right angles between the environments indicated that some varieties were consistent in high (‘Misrach’) or low ranking (‘Demhay’) while others changed markedly.

Polygon view for yield data from 2005 in all locations. The names of the test sites are spelled out in upper case, BO Bolenta; HAB Habes; BUK Buket; MKH Mekan; MG Mugulat; NK Neksege; MR Menkere; HO Holleta; DB Debrebrhan; AD Adinefas; MF Melfa; HBFT Habes FTC; MK Mekelle; FA FALA
Relationship of GEI to environmental factors
The relationships between the yield levels in various environments and various environmental factors were visually investigated using bilinear decomposition obtained by PLSR (Fig. 7a, b). Here, the GEI matrix (Y matrix) is regressed on the environmental variables (X matrix) to arrive at the hidden ‘latent’ variables, also called PCs or ‘PLS’ factors. The first PLS factor explained 33% and the second about 8% of the variation in GY (Fig. 7a), again separating the two high yielding sites (Holleta and Debrebrhan) from the 12 low yielding sites of Tigray. A high GY is associated with high TN (%), high rainfall in all months and low maximum temperatures in July and August (opposite positions in the plot). In Fig. 7b, only Tigray locations are displayed. Rainfall in June and to some extent July and high pH now had a negative impact on GY. This may reflect the effects of water logging, just as the positive associations of early and late rains reflect water limitation.

Biplot of the first and second partial least squares factors representing the X scores of ten genotypes and the Y loadings of 21 environments; Abbreviations for a and b: Temperature (MAXTM maximum temperature in May; MAXTJ in June; MAXTJUL in July; MAXTAG in August; MINTM minimum temperature in May; MINTJ in June; MINTJL in July; MINTAG in August); Rainfall (RM rainfall in May; RJ in June; RJL in July; RAG in August; RSEP in September); Elevation (ALT altitude) and soil factors (TN(%) total nitrogen; pH soil pH, OM Organic Matter). Variables marked with a circle are significant at P < 0.05 probability
Associations among days to flowering (DAFL), days to maturity (DAM), plant height (PH), thousand grain weight (TGWT) and grain yield (GY)
The correlation among these five traits was graphically analyzed using genotype by trait biplot (Fig. 8a). GY and PH were positively correlated, and they were negatively correlated with DAM and DAFL. The highest yielding varieties ‘Himblil’, ‘Local’, and ‘Misrach’ were all relatively tall and early varieties. Variety Demhay had the lowest TGWT. The location by trait biplot (Fig. 8b) shows the environmental correlations among traits. Again GY and PH were positively correlated and both had high values in Holleta. DAM and DAFL were positively correlated and they had high values at the high altitudes of Debrebrhan but low in TGWT. Therefore, late varieties are better adapted to Debrebrhan and Holleta.

Genotype by trait and environment by trait biplot showing the relation among traits across environments and genotypes, GY grain yield; TGWT thousand grain weight; DAM days to maturity, PH plant height, and DAFL days to flowering
Farmer’s preferences and perceptions
The overall farmers’ preferences across the seven sites are presented in Fig. 9. The preference for high grain yield was only significant for ‘Himblil’. Over all, the preference was only significant for ‘Himblil’ and ‘Misrach’, the latter for its overall merit, indicating the multiple criteria and flexibility of farmers in maintaining varieties (Table 5). ‘HB-42’ was rejected by 98% of experimenter farmers (Fig. 9). ‘Dimtu’ was preferred by farmers in Mugulat because they observed its ‘stay green’ trait during late season drought. Thus the preference ranking corresponds well with the conventional experimental data. The experiments also allowed information exchange among farmers and increased the familiarity with new varieties of barley. The expansion of the area planted with ‘Himblil and Misrach’ in 2005 indicated a fast adoption by farmers. The variation between villages for the area allocation to each variety was associated with the adaptation of varieties. Thus the PVS showed tangible results already in its second year.

Overall preferences ranking of barley varieties in three districts of Tigray
Discussion
If mega-environments are defined as “a group of locations that consistently share the same best cultivars (s)” (Yan et al. 2000), the results obtained here suggests that Tigray and the central highlands are two different mega-environments. The correlations between performance under stress and under favourable growing conditions were poor, indicating high genotype environment interactions (Ceccarelli et al. 1992; Ceccarelli 1994, 1996).
The nature of GEI that cause the crossovers among winning genotypes can be positively exploited by selecting different cultivars for the two mega-environments (Ceccarelli et al. 1998; Annicchiarico et al. 2005; Yan et al. 2007). Once the target regions are clarified, the knowledge of GEI can reduce the cost of extensive genotype evaluation by eliminating unnecessary testing sites and by fine-tuning breeding programs (Yan and Tinker 2006). Annicchiarico et al. (2005) found a positive advantage of selection for specific adaptation to winter temperatures in durum in Algeria, with a genetic correlation between the two sub regions of 0.66. In our case the correlation was slightly negative. The advantages of specific adaptation for barley in Syria observed by Ceccarelli et al. (1998) correspond to our results and may reflect the differences in environments as well as the well-known greater interactions in a diploid like barley versus, polyploid wheats.
The superior performance of ‘Himblil’ under water logged soils indicates a potential genotypic variability for water logging tolerance also observed in other trials (Fekadu, 2006, personal communication). Efficient nurseries for screening for waterlogging tolerance will be an important next step, given the complexity of water stresses, involving both deficiency and excess in the phenology of the crop. As shown in Chinese barleys, waterlogging tolerance may have a high heritability and is controlled mainly by GCA effects (Zhou et al. 2007). The genetics of waterlogging tolerance in ‘Himblil’ is being investigated and compared with the Chinese and Japanese sources. Waterlogging is a widespread problem in Ethiopia and not confined to barley. In durum wheat the landraces are superior in this trait than introduced germplasm. Any introduced germplasm from CIMMYT needs to have this specific adaptation (Rajaram, pers. comm.)
Drought in Ethiopia is associated with late season drought, but the situation described in this paper is more complex. The significant positive relation of yield with rainfall in May, and the negative association in June is associated with early season drought and temporary water logging problems and underdeveloped roots, respectively. An indication of the latter is found in the ‘residual moisture’ cultivation of barley widespread during the dry season. This needs to be considered when designing breeding strategies for the area. The large and significant genotypic component of variation in days-to-heading and maturity together with the association with grain yield indicates that the trait can be effectively manipulated within the existing materials in Ethiopia. To combine grain yield and long stature with earliness in heading and maturity is consistent with earlier studies of barley in Tigray (Abay and Cahalan 1995) and in Syria (van Oosterom et al. 1993; van Oosterom and Ceccarelli 1993), showing that flowering date shows relatively simple genetic control for dry land cereal crops. Sinebo (2002) identified a positive influence of early maturity and early shoot vigor under low nitrogen. The association of high throusand grain weights with low yielding environments is in agreement with the earlier study by Abay and Cahalan (1995) and Hadjichristodoulou (1987).
Choice of breeding strategies and genetic diversity
The combination of high rainfall and high N levels seem unrepresentative of the environments in Tigray. The response of ‘Himblil’ to low input and low yielding environments and of ‘Shege’ to high nitrogen and high rainfall soils can be associated to their respective selection environments. The inferiority of ‘HB-42’—obtained from a cross with exotic germplasm in addition reflects a misconceived germplasm strategy.
‘Misrach’ and ‘Dimtu’, the best performing varieties in Holleta and Debrebrhan, though selected there, reflect a major positive shift in barley breeding using relevant germplasm and testing methods performed by Ethiopian Institute of Agricultural Research (EIAR) since 1988 (Birhanu et al. 2005; Sinebo 2002). ‘Misrach’ is one of the >1,200 pure lines extracted from 180 tested landraces tested across three selection environments, of which one is Debrebrhan. That it comes from a landrace in Arsi and promoted in Debrebrhan due to a superior water logging tolerance, puts the ‘local’ adaptation hypothesis into doubt. Indeed, the regression coefficients of ‘Misrach’ and ‘Dimtu’ imply that the repeated testing over different environments has given them a wide adaptation and stability. However, the expected yield advantages of ‘Misrach’ and ‘Dimtu’ based on the previous breeding were not met in Tigray. Though only based on 1 year data, the high rainfall received in 2005 might be the best environment that the IVs developed at Debrebrhan and Holleta can expect. If these varieties did not excel then, it is doubtful that they would do better in the normal occurrence of drought. A reason why ‘Misrach’ did best of the IVs may be the selection for water logging tolerance at Debrebrhan. Still it was significantly inferior to the local check at the most waterlogged site Menkere. Also it is still surprising that ‘Himblil’—selected with much lower selection intensity from one landrace by the farmer in Bolenta—performed better than ‘Misrach’. This indicates that with a wider genetic diversity of germplasm from Tigray, even better varieties would have been produced. The selection environment of ‘Himblil’ (and ‘Demhay’) had unusually high N and OM (and P, not shown) content for Tigray, having been richly manured over years. The selector expressed that this allowed a better differentiation between plants. His primary selection goal was a quality comparable to tef for making thetolerance to water logging as well as the short rainy season in Tigray.
The merit of PVS for barley improvement in Tigray
The breeding history of ‘Himblil’ and these experiments have shown that PVS is a viable method for identifying preferences, constraints and the potential of varieties. The PRA methods corresponded well with the data from the mother and baby trials, and the analysis showed that the level of accuracy was acceptable. Also the preliminary experiences of variety dissemination are promising. For further work, strong collaborative networks have been established between the farming communities and regional extension systems. The genetics of waterlogging tolerance in ‘Himblil’ is being investigated in a cross with ‘Saesa’, representing the very early two-rowed barleys, hopefully also elucidating the adaptation in the latter group of barleys not dealt with in this paper.
References
Aastveit AH, Martens H (1986) ANOVA interactions interpreted by partial least squaresregression. Biometrics 42:829–844. doi:10.2307/2530697
Abay F, Cahalan C (1995) Evaluation of response of some barley landraces in drought prone sites of Tigray (northern Ethiopia). Crop Improv 22:125–132
Abay F, Waters-Bayer A, Bjørnstad A (2008) Farmers’ seed management and Innovation in varietal selection: implications for barley breeding in Tigray, northern Ethiopia. Ambio 37(4):312–320. doi:10.1579/0044-7447(2008)37[312:FSMAII]2.0.CO;2
Ahokas H, Manninen M-L (2001) Polymorphisms of phosphate acquisition parameters in barley (Hordeum vulgare) landraces: selected acid phosphatase and milieu acidification of roots after germination in vitro. Biol Agric Hortic 18:385–399
Annicchiarico P (2002) Genotype × environment interactions: challenges and opportunities for plant breeding and cultivar recommendations. FAO Plant Production and Protection Paper No. 174. FAO, Rome
Annicchiarico P, Bellah A, Chiari T (2005) Defining sub regions and estimating benefits for a specific-adaptation strategy by breeding programs: a case study. Crop Sci 45:1741–1749. doi:10.2135/cropsci2004.0524
Atlin GN, Frey KJ (1989) Predicting the relative effectiveness of direct versus indirect selection for oat yield in three types of stress environments. Euphytica 44:137–142. doi:10.1007/BF00022608
Atlin GN, Cooper M, Bjørnstad Å (2001) A comparison of formal and participatory breeding approaches using selection theory. Euphytica 122:463–475. doi:10.1023/A:1017557307800
Birhanu B, Fekadu A, Berhane L (2005) Food barley in Ethiopia, pp 53–81. In: Grando S, Helena GM (eds) Food barley importance, uses and local knowledge. Proceedings of the international workshop on food barley improvement, Hammamet, Tunisia, ICARDA, Aleppo, Syria
Ceccarelli S (1989) Wide adaptation. How wide? Euphytica 40:197–205
Ceccarelli S (1994) Specific adaptation and breeding for marginal conditions. Euphytica 77:205–219. doi:10.1007/BF02262633
Ceccarelli S (1996) Positive interpretation of genotype by environment interactions in relation to sustainability and biodiversity. In: Cooper M, Hammer GL (eds) Plant adaptation and crop improvement. CAB International, Wallingford, pp 467–486
Ceccarelli S, Grando S (2007) Decentralized-participatory plant breeding: an example of demand driven research. Euphytica 155:349–360. doi:10.1007/s10681-006-9336-8
Ceccarelli S, Grando S, Hamblin J (1992) Relationships between barley grain yield measured in low and high yielding environments. Euphytica 64:49–58
Ceccarelli S, Grando S, Impiglia A (1998) Choice of selection strategy in breeding barely for stress environments. Euphytica 103:307–318. doi:10.1023/A:1018647001429
Ceccarelli S, Grando S, Baum M (2007) Participatory plant breeding in water-limited environment. Exp Agric 43:411–435. doi:10.1017/S0014479707005327
Christinck A, Weltzien E, Hoffmann V (eds) (2005) Setting breeding objectives and developing seed systems with farmers. A handbook for practical use in participatory plant breeding projects. Margraf Publishers, Weikersheim
Eberhart SA, Russell WA (1966) Stability parameters for comparing varieties. Crop Sci 6:36–40
Fekadu W, Skjelvag AS (2002) Sowing date effects on growth rate and straw yield of local cultivars. NLH PhD thesis 36: 1–20
Finlay KW, Wilkinson GN (1963) The analysis of adaptation in a plant-breeding program. Aust J Agric Res 14:742–754. doi:10.1071/AR9630742
Gauch HG, Zobel RW (1997) Identifying mega-environments and targeting genotypes. Crop Sci 37:311–326
Górny AG (2001) Variation in utilisation efficiency and tolerance to reduced water and nitrogen supply among wild and cultivated barleys. Euphytica 117:59–66. doi:10.1023/A:1004061709964
Hadjichristodoulou A (1987) Stability of 1,000 grain weight and its relation with other traits of barley in dry land areas. Euphytica 51:11–17
Holland JB, Bjørnstad Å, Frey KJ, Gullord M, Wesenberg DM (2001) Reccurent selection for broad adaptation affects stability of oat. Euphytica 126:265–274
Hussein MA, Bjørnstad Å, Aastveit AH (2000) SASG × ESTAB: a SAS program for computing genotype × environment stability statistics. Agron J 92:454–459
IRRI (2002) International Rice Research Institute. IRRISTAT 4.3 for Windows. www.irri.org
Martens H, Naes T (1989) Multivariate calibration. Wiley, New York
Mitiku H, Berhanu G, Amare B (2003) The status of soil fertility in Tigray. In: Gebremedhin B, Pender J, Endu SK (eds) Policies for sustainable land management in the highlands of Tigray, northern Ethiopia. Socio-economics and policy research working paper 54. ILRI, Nairobi
Muruli BJ, Paulsen GM (1981) Improvement of nitrogen use efficiency and its relationship to other traits in maize. Maydica 26:63–73
Perkins JM, Jinks JL (1968) Environmental and genotype-environmental components of variability IV. Non-linear interactions for multiple inbred lines. Heredity 23:525–535. doi:10.1038/hdy.1968.71
SAS (2001) SAS user’s guide, released 9th edn. SAS Institute Inc., Cary
Simmonds NW (1991) Selection for local adaptation in plant breeding program. Theor Appl Genet 82:363–367. doi:10.1007/BF02190624
Sinebo W (2002) Yield relationships of barleys grown in a tropical Highland environment. Crop Sci 42:428–437
Sinebo W, Gretzmacher R, Edelbauer A (2002) Environment of selection for grain yield in low fertilizer input barley. Field Crops Res 74:151–162. doi:10.1016/S0378-4290(01)00207-6
Snapp S (1999) Mother and baby trials: a novel trial design being tried out in Malawi. Target Newsl South Afr Soil Fertil Netw 17:8–10
van Oosterom EJ, Ceccarelli S (1993) Indirect selection for grain yield of barley in harsh Mediterranean environments. Crop Sci 33:1127–1131
van Oosterom EJ, Ceccarelli S, Peacock JM (1993) Yield response of barley to rainfall and temperature in Mediterranean environments. J Agric Sci Camb 121:307–314
Vargas M, Crossa J, Sayre K, Reynolds M, Ramirez ME, Talbot M (1998) Interpreting genotype × enviornment interaction in wheat using partial least squares regression. Crop Sci 38:679–689
Vaughan DA, Chang TT (1992) In situ conservation of rice genetic resources. Econ Bot 46:368–383
Yan W (2001) GGEbiplot a windows application for graphical analysis of multi-environment trial data and other types of two-way data. Agronomy 93:1111–1118
Yan W, Rajcan I (2002) Biplot analysis of test sites and Trait relations of soyabean in Ontario. Crop Sci 42:11–20
Yan W, Tinker NA (2006) Biplot analysis of multi-environment trial data: principles and applications. Can J Plant Sci 86:623–645
Yan W, Hunt LA, Sheng Q, Szlavnics Z (2000) Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Sci 40:597–605
Yan W, Kang MS, Ma B, Woods S, Cornelius PL (2007) GGE Biplot vs. AMMI analysis of genotype-by-environment data. Crop Sci 47:641–653
Zhou MX, Li HB, Mendham NJ (2007) Combining ability of waterlogging tolerance in barley. Crop Sci 47:278–284. doi:10.2135/cropsci2006.02.0065
Zobel RW, Wright MJ, Gauch G (1988) Statistical analysis of a yield trial. Agron J 80:388–393
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Abay, F., Bjørnstad, A. Specific adaptation of barley varieties in different locations in Ethiopia. Euphytica 167, 181–195 (2009). https://doi.org/10.1007/s10681-008-9858-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-008-9858-3