
Abstract
White lupin (Lupinus albus) and narrow-leafed lupin (L. angustifolius) have special interest as high-protein feed crops but their cultivation is limited by low grain yields. This study aimed to support breeding programs targeting Italy or other climatically variable south-European regions by investigating within-species adaptation patterns across contrasting Italian environments. An additional aim was comparing species for yielding ability. Eight narrow-leafed and six white lupin cultivars featuring different origin, phenological type (Mediterranean in both species; winter in white lupin; spring in narrow-leafed lupin) and plant architecture (determinate or indeterminate in both species; tall or dwarf in white lupin) were evaluated in a Mediterranean and a subcontinental-climate site under autumn and late-winter sowing. Additive main effects and multiplicative interaction was preferable to joint regression for modeling yield responses. In both species, cross-over GE interaction was observed (P < 0.05), autumn-sown Mediterranean and subcontinental environments were the most-contrasting for GE effects, and widely adapted material included cultivars of Mediterranean phenological type with indeterminate growth. Material with determinate growth was not among the best-yielding entries in any environment, whereas a dwarf winter-type white lupin entry was specifically adapted to autumn-sown subcontinental environments. White lupin displayed larger genetic variation than narrow-leafed lupin for phenology and other traits. Relationships of morphophysiological traits with grain yield were environment-specific and were locally high for some white lupin traits (early flowering, long reproductive phase, high aerial biomass, low proportion of pod wall). White lupin exhibited higher yielding ability than narrow-leafed lupin in all environments but the late-winter sown Mediterranean one, when comparing locally top-yielding cultivars.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Feed grain legumes are strategically important to decrease the marked deficit of high-protein feedstuff and increase in various respects (safeguard of soil fertility and biodiversity; reduction of greenhouse gas emissions and nitrogen fertilizers) the sustainability of European crop-livestock systems (Carrouée et al. 2003; Jensen and Hauggaard-Nielsen 2003). Amongst them, white lupin (Lupinus albus) and narrow-leafed lupin (L. angustifolius) have outstanding grain protein content and are particularly suited to diets for ruminants, where they can replace completely the soybean meal (e.g., in traceable, GM-free feed chains) even for high-producing lactating cows (Froidmont and Bartiaux-Thill 2004). In addition, lupin grains have good potential as ingredients of functional or healthy food products (Arnoldi 2005). However, the economic sustainability of these or other feed legumes depends on the ability to increase their grain yields (Dronne 2003). Currently, the cultivation of lupins extends over just 3,000 ha in Italy, 5,000 ha in France and 6,000 ha in the Iberian peninsula (http://faostat.fao.org/).
Previous grain yield comparisons of lupin species were based on a few cultivars and provided region-specific results, e.g., the advantage of white lupin in southern Spain (Lopez-Bellido and Fuentes 1990) and that of narrow-leafed lupin in Australia (Siddique et al. 1993). Recent breeding work has considerably widened the range of lupin plant types available for cultivation, but there is little information on the adaptation of these types to south-European environments and its implications for the choice of species and plant type for local breeding targeted to these environments. The selection of definitely winter-hardy white lupins has enhanced the opportunities for autumn sowing which, in comparison with spring sowing, allows for increasing the yield potential through the longer crop cycle (Papineau and Huyghe 2004). However, winter hardiness is related to delayed floral initiation (via greater vernalization requirement) besides intrinsic frost tolerance (Huyghe and Papineau 1990) and may, therefore, increase the crop susceptibility to terminal drought. Indeed, improved adaptation of narrow-leafed lupin to autumn-sown, drought-prone environments of Australia has largely relied on the selection for earlier phenology by removing the vernalization requirement (French and Buirchell 2005). Large genotype × environment (GE) interaction in south-European regions may arise from the simultaneous diversity in: (1) phenological germplasm type, including Mediterranean (adapted to autumn sowing in mild-winter areas), European winter and European spring types (Stoddard et al. 2006); and (2) climate, ranging from subcontinental (e.g., northern Italy) to Mediterranean (e.g., southern Italy), or sowing time (autumn or late-winter sowing, the latter being adopted when unfavourable climatic conditions prevent the autumn sowing).
Determinate growth (i.e., restricted branching) of white or narrow-leafed lupin, which is under monogenic control (Huyghe 1997; Adhikari et al. 2001), has been selected as a means to increase the crop earliness and harvest index. However, the advantage of this plant type has proved region-specific. Determinate white lupin germplasm outyielded conventional, indeterminate one in cool, short-season environments of UK while being outyielded in temperate environments of France (Julier et al. 1993). The advantage of determinate narrow-leafed lupins emerged in a favourable, high-rainfall site but not in two drought-prone sites of Australia (Galwey et al. 2003). No consistent advantage of determinate narrow-leafed lupins was found in areas of northern Europe (Joernsgaard et al. 2004). Dwarfism, which is controlled by two recessive genes which reduce the internode length on the main stem, also has a major impact on the white lupin plant architecture and showed the potential for increasing the crop harvest index and tolerance to lodging (Harzic et al. 1995, 1996). A novel white lupin type which combines dwarfness and determinacy has recently become available (Papineau and Huyghe 2004). However, the adaptation pattern of dwarf versus tall genotypes has not been thoroughly compared.
In this study, several narrow-leafed and white lupin cultivars featuring different phenological types and plant architectures were evaluated across Italian environments which contrasted for climate and sowing time, with the objective of: (a) investigating the adaptive response of cultivars within species in relation to their plant type and morphophysiological characteristics; (b) assessing the environment similarity for GE interaction effects; and (c) comparing the lupin species for yielding ability as determined by the locally best-performing cultivar. The generated information can support the definition of objectives and selection strategies for breeding programs targeting Italy or other climatically-variable south-European regions.
Materials and methods
Experimental data
The study included: (a) eight recent genotypes of narrow-leafed lupin released by two breeding institutions of Australia, two of Germany and one of Byelorussia, of which seven were indeterminate and one was determinate; (b) four recent genotypes of white lupin bred in France or Spain, of which two were tall determinate, one was dwarf determinate and one was dwarf indeterminate, and one Italian landrace and one Italian cultivar of white lupin with conventional plant architecture (Table 1). Phenological types as inferred from the origin of entries or other sources (e.g., Papineau and Huyghe 2004) were Mediterranean or European spring for narrow-leafed lupin, and Mediterranean or European winter for white lupin (Table 1). Australian narrow-leafed lupins were considered Mediterranean-type as they relied largely on Mediterranean genetic resources (Buirchell and Cowling 1998).
The evaluation sites were: i) Lodi (Lombardy), representative of the subcontinental climate with extended frosts in winter and moderate terminal drought which is typical of northern Italy; ii) Sanluri (Sardinia), representative of the Mediterranean climate with occasional short frost periods within mild winters and terminal drought which is widespread in southern Italy and coastal areas of central Italy (Perini et al. 2004). The soils, featuring pH values of 6.2 in Lodi and 7.4 in Sanluri and active lime below 0.4%, were suitable for lupins (Dracup et al. 1998; Papineau and Huyghe 2004). The study included five test environments defined by the combination of location and sowing time. They represented an early- and a late-autumn sowing (in the cropping season 2002–2003) and a late-winter sowing (in 2004–2005) in Lodi, and an autumn and a late-winter sowing (in 2003–2004) in Sanluri (Table 2). The autumn-sowing dates were site-specific depending on the climatic area. The adoption of two autumn sowings in the cold-prone site was justified by the fact that winter plant mortality may arise from too early floral initiation under early sowing or from insufficiently thick root parenchyma under late sowing (Huyghe and Papineau 1990). Narrow-leafed and white lupin cultivars were tested together in each trial. The two sowing dates of Lodi in 2002–2003 or Sanluri were assigned to main plots, and the cultivars grouped for species to subplots, combining features of strip-plot and group-block designs (Gomez and Gomez 1984, pp. 75–83). The trial of Lodi in 2004–2005 was designed as a group-block design, grouping cultivars of same species within each randomized complete block. There were always three replications per treatment.
Prior to sowing, seeds were treated with Germipro UFB (350 g/l Iprodione + 177 g/l Carbendazim) at the rate of 2.6 ml/kg and were inoculated with NPPL HiStick (Becker Underwood). Seed rates, defined as a compromise between Fagnano and Bozzini’s (2001), Perry et al.’s (1998) and Papineau and Huyghe’s (2004) indications, were 80 and 45 germinating seeds/m2 for narrow-leafed and white lupin, respectively. The sowing depth was 30 mm. The plots were 9 m2, and included 16 rows 3 m long, 180 mm apart for narrow-leafed lupin, and 8 rows 3 m long, 360 mm apart for white lupin. The plot harvest area excluded 8 (narrow-leafed lupin) or 4 (white lupin) edge rows. Grain yield was expressed at 13% seed moisture (after moisture determination on a random sample of 250 seeds per plot).
Days to onset of flowering (from March 1 to when 50% of plants had the first open flower) was recorded in all experiments. The entries were characterized for the following traits across Lodi’s autumn-sown environments: i) winter plant mortality, based on plant counts at the onset and the end of winter along one linear m in each of two rows; ii) days to maturity (from March 1 to when 50% of the plants were ripe for harvesting), and length of the reproductive period; iii) percent of lodged plants at maturity, based on plant counts on two central rows; iv) aerial (grain + straw) dry matter and harvest index, weighting as straw all the residues from threshing from the combine cutting height and assessing their moisture on a sample of about 1 kg; v) proportion of seeds on the main stem, individual seed weight, and proportion of pod wall on pod biomass for pods on the main stem [as: (pod weight − seed weight)/pod weight], estimated on the oven-dried seeds or pods as average values of six random ripe plants per plot.
Statistical analysis
A combined analysis of variance (ANOVA) was performed separately for each species on grain yield data, holding environment and genotype as fixed factors and considering each environment as a randomized complete block experiment. GE interaction was partitioned by joint regression (Finlay and Wilkinson 1963) and additive main effects and multiplicative interaction (AMMI) (Gauch 1992) analysis, selecting the AMMI model whose principal component (PC) axes reached P < 0.01 significance according to the F R test recommended by Piepho (1995). AMMI-modeled yield responses, which improve the prediction of cultivar responses theoretically (Gauch 1992, pp. 134–153) and empirically (Annicchiarico et al. 2006), were graphically displayed as entry nominal yields as a function of the environment PC 1 score. Nominal yields, which sum up the estimated entry mean value and the product of the entry by the site scaled scores on PC 1 (excluding site main effects, irrelevant for entry ranking), allow for linearizing the adaptive responses (Gauch and Zobel 1997). Entry differences in a given environment were assessed according to the ANOVA’s LSD value for the relevant environment.
Within-species variation for morphophysiological traits was assessed by an ANOVA including the fixed factors: i) environment and genotype, for onset of flowering in five environments; ii) sowing time and genotype, for all traits recorded in Lodi’s autumn-sown environments (according to the split-plot lay-out of these factors). The relationship of mean grain yield or AMMI-modeled yield in each of the most-contrasting environments for GE interaction with morphophysiological traits of the cultivars (averaged across environments) was assessed by simple correlation analysis.
Other ANOVAs aimed at comparing species and assessing species × environment interaction. They included the fixed factors species, genotype within species, and environment (for grain yield or onset of flowering in five environments) or sowing time (for morphophysiological traits recorded in Lodi’s autumn-sown environments). Species × block interaction (nested into environment, when relative to five environments) acted as the error term for testing species main effects, species × environment or species × sowing time interaction, to take account of the group-block lay-out of species and cultivars (Gomez and Gomez 1984, pp. 75–83).
Species were also compared in each environment for: i) mean grain yield; ii) AMMI-modeled grain yield (inclusive of the site main effect) of the top-yielding entry of each species. Both comparisons held the ANOVA species × block interaction mean square for the specific environment as the error term. The entry comparison was based on the LSD value including this error.
Statistical Analysis System software (SAS 1999) was used for all analyses but AMMI and joint regression, which were performed by CropStat (IRRI 2008).
Results
The subcontinental-climate site (Lodi) confirmed the expected greater extent and severity of winter frosts relative to the Mediterranean site (Sanluri) and, in comparison with long-term values, exhibited a drier spring in 2002–2003 (Table 3). Sanluri had a somewhat colder winter and a wetter spring in the test year relative to the long term (Table 3). Potential evapotranspiration in spring was not available but was known to be over 50 mm greater from March 1 to June 15 in Sanluri relative to Lodi in the long term (Perini et al. 2004), justifying the fairly similar mean grain yield of Sanluri’s and Lodi’s autumn-sown environments (Table 2). Sanluri’s late winter-sown environment was the lowest-yielding environment for both species (Table 2).
Entry mean values for grain yield and onset of flowering across environments are given in Table 1. In narrow-leafed lupin, top-yielding material had indeterminate habit and was mostly bred in Australia. The range of flowering time was fairly narrow (about 7 days) and unrelated to phenological type or mean yield (r = − 0.34, P > 0.10) of the genotypes. In white lupin, the Italian cultivars with conventional plant architecture were top-yielding and early-flowering, whereas the bottom-yielding entry was dwarf determinate and notably late-flowering. The entry range for onset of flowering was large (over 25 days), associated with phenological type (early in Mediterranean entries, and intermediate or late in winter-type germplasm: Table 1), and inversely related to entry mean yield (r = − 0.88, P < 0.05).
For both species, GE interaction for grain yield was highly significant (P < 0.001) and the selected AMMI model, including one PC axis, was preferable to joint regression on the basis of: i) its greater GE interaction variation accounted for (R 2 = 85% vs. 12% for narrow-leafed lupin, and 89% vs. 3% for white lupin); ii) the highly significant deviations from regression term (P < 0.001) (Table 4). The autumn-sown Mediterranean environment and the autumn-sown subcontinental environments were the most-contrasting for GE effects in both species according to the environment ordination on the first GE interaction PC axis (Figs. 1, 2). Spring-sown environments of the two sites exhibited similar entry responses, and were more similar to Lodi’s autumn-sown environments in narrow-leafed lupin (Fig. 1) and to Sanluri’s autumn-sown environment in white lupin (Fig. 2).

Nominal grain yield of eight narrow-leafed lupin cultivars as a function of the score on the first genotype × environment interaction principal component (PC) axis of five environments (average LSD at P < 0.05 = 0.61; LEA, LLA and LW Lodi’s early-autumn, late-autumn and late-winter sowings, respectively; SA and SW Sanluri’s autumn and late-winter sowings, respectively)

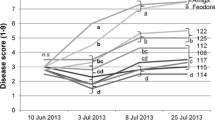
Nominal grain yield of six white lupin cultivars as a function of the score on the first genotype × environment interaction principal component (PC) axis of five environments (average LSD at P < 0.05 = 0.78; LEA, LLA and LW Lodi’s early-autumn, late-autumn and late-winter sowings, respectively; SA and SW Sanluri’s autumn and late-winter sowings, respectively)
Tall determinate genotypes, namely ‘Walan 2023’ in narrow-leafed lupin and ‘AB 47’ and ‘Ludet’ in white lupin, were markedly unadapted to Lodi’s autumn-sown environments (Figs. 1, 2) but failed to enter the top-yielding set (P < 0.05) also in the other environments. The Byelorussian narrow-leafed line ‘HP 39-1’ and the dwarf indeterminate white lupin ‘Luxe’ tended to the opposite adaptive response, being specifically adapted to Lodi’s cold-prone environments and poorly adapted to the other environments (Figs. 1, 2). Cross-over GE interaction (P < 0.05) occurred between ‘HP 39-1’ and ‘Walan 2023’ in narrow-leafed lupin, and between ‘Luxe’ and ‘AB 47’ in white lupin. The sizeable extent of GE effects was also supported by the fact that the two most-contrasting environments for PC 1 score tended towards inverse correlation in narrow-leafed lupin (r = − 0.66, P < 0.10) and were not correlated in white lupin (r = 0.27, P > 0.30) for AMMI-modeled entry yields. However, widely adapted material could also be found, such as ‘Jindalee’ in narrow-leafed lupin (Fig. 1) and ‘Multitalia’ and the Italian ecotype in white lupin (Fig. 2).
Genetic variation for morphophysiological traits (P < 0.10) was detected for all traits but susceptibility to lodging in narrow-leafed lupin. This species displayed narrower genetic variation than white lupin also for other traits besides onset of flowering and lodging, such as maturity time, length of the reproductive phase, proportion of seeds on the main stem, dry seed weight and proportion of pod wall, while exhibiting wider variation than white lupin only for winter plant mortality, according to range values reported in Table 5. White lupin also showed genotype × sowing time interaction (P < 0.05) for some morphophysiological traits across Lodi’s autumn-sown environments, i.e., onset of flowering, maturity date, length of the reproductive phase, aerial dry matter and winter plant mortality. GE interaction for onset of flowering across five environments was sizeable in narrow-leafed lupin (P < 0.01) and large in white lupin (P < 0.001), where it was largely due to quantitative interaction of phenological type with location. In particular, the flowering delay of winter white lupins relative to Mediterranean ones was much wider in Sanluri’s environments (averaging 25 days) than in Lodi’s environments (averaging 12 days).
Correlation results between morphophysiological traits over Lodi’s autumn-sown environments and grain yield of entries in the most contrasting environments for GE interaction, i.e., autumn sowing in Sanluri (for both species) and early-autumn (for narrow-leafed lupin) or late-autumn (for white lupin) sowing in Lodi, are reported in Table 5. Higher yield in the Mediterranean environment was a feature of entries with earlier onset of flowering and longer reproductive phase in white lupin (P < 0.05), whereas no distinct relationship emerged in narrow-leafed lupin (where the trend to inverse correlation with harvest index was associated with the low level of genetic variation for this trait) (Table 5). Higher yield in the relevant autumn-sown subcontinental environment was related (P < 0.10) to later maturity and greater aerial biomass in both species, and to lower proportions of pod wall and of seed on the main stem in white lupin (Table 5). As expected, determinate lines had higher proportion of seed on main stem than indeterminate lines in both species (P < 0.05). ‘Luxe’ and ‘Ludet’ in white lupin, and ‘Walan 2053’ and ‘HP 39-1’ in narrow-leafed lupin, were top-ranking for harvest index.
Species comparison for grain yield was affected by species × environment interaction (P < 0.05). On average, white lupin was higher yielding in Lodi and lower yielding in Sanluri than narrow-leafed lupin under late-winter sowing (P > 0.10), whereas the two species did not differ in the autumn-sown environments (Table 2). The comparison of greater practical interest, based on material which maximized locally the species yielding ability (i.e., ‘Multitalia’ in all cases for white lupin, and ‘Quilinock’ or ‘Jindalee’ for narrow-leafed lupin), highlighted a trend towards higher yield of white lupin, as: i) ‘Multitalia’ outyielded the top-yielding narrow-leafed entry also in all autumn-sown environments besides the late winter-sown in Lodi (P < 0.10); ii) the two species did not differ in Sanluri under late-winter sowing (Table 2). On average, white lupin had later onset of flowering and maturity, longer reproductive phase, lower winter mortality, somewhat greater lodging, greater proportion of seed on the main stem, lower proportion of pod wall and heavier seed than narrow-leafed lupin over Lodi’s autumn-sown environments (Table 5).
Discussion
The present indications on species adaptation and within-species GE interaction patterns should not be considered as conclusive, given the limited sampling of germplasm (especially for some plant types within species) and of environments within each climatic area. The sharp contrast between subcontinental-climate and Mediterranean-climate environments under autumn sowing which emerged for both species in the AMMI ordination for GE effects agrees with previous studies performed in Italy on faba bean (Annicchiarico and Iannucci 2008) and small-grain cereals such as bread wheat, durum wheat or oat (Annicchiarico 1997), where genotype × location (GL) interaction effects for autumn-sown environments were large and mainly accounted for by the two climatic areas. The substantial repeatability across years of GL effects in these studies (whose trials were repeated also in time at each site) could not be verified here but suggests, on the whole, the overwhelming impact of the climatic area on the genotype adaptive response of autumn-sown grain legume or small-grain cereal crops in Italy. Indeed, the climatic differences between lupin test locations were less marked in the test years than the long term, especially for rainfall amount, suggesting that even larger GL interaction may emerge on test years whose rainfall amount in each site represented more faithfully the average difference between the two climatic areas.
Although preliminary, our findings have implications for Italian lupin breeding programs and may support the breeding work in other south-European regions which include a Mediterranean and a subcontinental (or oceanic) climatic area (such Spain, Portugal, southern France or the Balkan peninsula). The limited cropping area of lupins in south-European regions may lead breeding programs to concentrate resources on the improvement of only one lupin species. White lupin may be preferable to narrow-leafed lupin, as its lack of superiority according to top-yielding material (and its inferiority based on species mean yield) emerged only in the Mediterranean environment including late-winter sowing. This sowing time as determined by unfavourable climatic conditions for autumn sowing is expected to be much less frequent in Mediterranean-climate than subcontinental-climate areas. However, novel narrow-leafed lupin varieties selected in southern Europe (which may become available within a few years: Crinò and Saccardo 2008) may widen the adaptation of this species and reduce its disadvantage relative to white lupin in this region. An additional advantage of white lupin over narrow-leafed lupin is its higher grain quality for energy value (Petterson 1998) and protein content (averaging 39.5% vs. 31.2% for the present sets of cultivars over Lodi’s autumn-sown environments: Colombini et al. 2004). On the other hand, the greater earliness of maturity of narrow-leafed lupin may allow for the earlier sowing of a summer crop in irrigated areas.
Despite the large GE interaction effects observed in white lupin, the wide adaptation displayed by two cultivars having Mediterranean phenological type and conventional plant architecture (‘Multitalia’ and the Italian ecotype) suggests that breeding for wide adaptation may be feasible through the exploitation of Mediterranean-type genetic resources and the parallel selection of novel germplasm across a subcontinental-climate and a Mediterranean-climate site under autumn sowing (i.e., across two selection environments which contrast for GE effects and are representative of major target environments). The potential usefulness of a dwarf plant type requires further assessment, as one such entry (‘Luxe’) was as yielding as the best tall material in the cold-prone environments and may have failed to respond well to the other environments because of its winter phenological type (implying late cycle) rather than its dwarf stature. Indeed, the markedly delayed flowering of ‘Luxe’ and other winter varieties in Sanluri could be attributed to the difficulty of this mild-winter site to satisfy the high vernalization that is required by winter-type white lupins (Huyghe and Papineau 1990). Spring-type white lupins were not evaluated here, because earlier results highlighted their susceptibility to winter cold in subcontinental-climate environments and late frosts in Mediterranean environments of Italy (Annicchiarico and Iannucci 2007). The substantial lack of adaptation across environments of determinate white lupins of Mediterranean or winter phenological type suggests the greater interest of an indeterminate habit for south-European environments. Correlation results revealed no morphophysiological trait consistently related to higher white lupin yield across autumn-sown Mediterranean and subcontinental environments. However, they suggested that the features of the Mediterranean, conventional-type material (early onset of flowering, long reproductive phase, high aerial biomass), as well as low proportion of pod wall on pod biomass, were useful in one of these environments without being detrimental in the other. The usefulness of low proportion of pod wall agrees with results reported by Lagunes-Espinoza et al. (1999) for a larger set of white lupin genotypes.
The similarity between spring and Mediterranean narrow-leafed lupins for flowering and maturity time could be expected on the ground of selection for low or nil vernalization which Mediterranean germplasm bred in Australia usually underwent (French and Buirchell 2005). The slight advantage of Australian over central-European germplasm in terms of overall adaptation was probably due to better tolerance of this material to the terminal drought which features south-European environments. GE interaction in this species was mainly due to the marked specific-adaptation response of the Byelorussian entry ‘HP 39-1’. The great climatic contrast between selection and test environment may account for its very poor adaptation to Mediterranean environments, although no recorded morphophysiological trait could be related to its adaptation pattern. The adaptive response of the determinate Mediterranean narrow-leafed line ‘Walan 2023’ paralleled that of the determinate Mediterranean white lupin line ‘AB 47’, namely, a positive GE interaction effect in the autumn-sown Mediterranean environment insufficient to attain the yield level of the top-yielding material even in this environment (Figs. 1, 2). Our results suggest that also narrow-leafed lupin selection for wide adaptation may conveniently rely on Mediterranean germplasm with indeterminate habit. Within this germplasm, the limited GE interaction currently observed would allow to limit the evaluation across climatically contrasting environments to late selection stages. The identification of useful morphophysiological traits was hindered in this species by the only moderate genetic variation and the lack of high and consistent correlation with grain yield of these traits. Proportion of pod wall was not related to grain yield in any environment, despite the potential usefulness of this trait advocated also for this species (Clements et al. 2005).
In conclusion, this study supports the somewhat greater interest of white lupin over narrow-leafed lupin across Italian environments and, within both species, the usefulness of indeterminate, Mediterranean-type germplasm in breeding for wide adaptation. Breeding specifically for a Mediterranean- and a subcontinental-climate area may also be of interest for white lupin, where it could exploit different phenological types (e.g., winter material for subcontinental areas) and specific adaptive traits (e.g., early flowering for Mediterranean areas). Further work based on a large germplasm sample is required to verify whether the advantage in terms of yield gains provided by specific adaptation over wide adaptation is large enough to justify this adaptation strategy for a species whose seed market is fairly limited. The scope for specific breeding may be widened when targeting a Mediterranean and a subcontinental area across south-European countries, as suggested for faba bean by a tentative definition of variety adaptive zones (Metayer 2004).
References
Adhikari KN, Galwey NW, Dracup M (2001) The genetic control of highly restricted branching in narrow-leafed lupin (Lupinus angustifolius L.). Euphytica 117:261–274. doi:10.1023/A:1026571416075
Annicchiarico P (1997) Joint regression vs. AMMI analysis of genotype–environment interactions for cereals in Italy. Euphytica 94:53–62. doi:10.1023/A:1002954824178
Annicchiarico P, Iannucci A (2007) Winter survival of pea, faba bean and white lupin cultivars across contrasting Italian locations and sowing times, and implications for selection. J Agric Sci 145:611–622. doi:10.1017/S0021859607007289
Annicchiarico P, Iannucci A (2008) Breeding strategy for faba bean in southern Europe based on cultivar responses across climatically-contrasting environments. Crop Sci 48:983–991. doi:10.2135/cropsci2007.09.0501
Annicchiarico P, Bellah F, Chiari T (2006) Repeatable genotype × location interaction and its exploitation by conventional and GIS-based cultivar recommendation for durum wheat in Algeria. Eur J Agron 24:70–81. doi:10.1016/j.eja.2005.05.003
Arnoldi A (2005) Optimized processes for preparing healthy and added value food ingredients from lupin kernels, the European protein-rich grain legume. Aracne, Rome
Buirchell BJ, Cowling WA (1998) Genetic resources of lupins. In: Gladstones JS, Atkins C, Hamblin J (eds) Lupins as crop plants: biology, production and utilization. CABI, Wallingford, pp 41–66
Carrouée B, Crépon K, Peyronnet C (2003) Les protéagineux: intérêt dans les systèmes de production fourragers français et européens. Fourrages 174:163–182
Clements JC, Dracup M, Buirchell BJ, Smith CG (2005) Variation for seed coat and pod wall percentage and other traits in a germplasm collection and historical cultivars of lupin. Aust J Agric Res 56:75–83. doi:10.1071/AR03114
Colombini S, Annicchiarico P, Odoardi M (2004) Resa proteica di alcune varietà di pisello, fava e lupino nell’Italia settentrionale. Inf Agr 60(39):73–75
Crinò P, Saccardo F (2008) Cece e lupino italiani ancora una risorsa. Inf Agr 64(17):47–49
Dracup M, Turner NC, Tang C, Reader M, Palta J (1998) Responses to abiotic stresses. In: Gladstones JS, Atkins C, Hamblin J (eds) Lupins as crop plants: biology, production and utilization. CABI, Wallingford, pp 385–409
Dronne Y (2003) L’approvisionnement en protéines de la France dans son contexte européen et mondial. Fourrages 174:107–128
Fagnano M, Bozzini A (2001) Lupino (Lupinus albus L.). In: Ranalli P (ed) Leguminose e agricoltura sostenibile—specie da granella e cover crops. Calderini Edagricole, Bologna, pp 591–614
Finlay KW, Wilkinson GN (1963) The analysis of adaptation in a plant-breeding programme. Aust J Agric Res 14:742–754. doi:10.1071/AR9630742
French RJ, Buirchell BJ (2005) Lupin: the largest grain legume crop in western Australia, its adaptation and improvement through plant breeding. Aust J Agric Res 56:1169–1180. doi:10.1071/AR05088
Froidmont E, Bartiaux-Thill N (2004) Suitability of lupin and pea seeds as a substitute for soybean mean in high-producing dairy cow feed. Anim Res 53:475–487. doi:10.1051/animres:2004034
Galwey NW, Adhikari K, Dracup M, Thomson R (2003) Agronomic potential of genetically diverse narrow-leafed lupins (Lupinus angustifolius L.) with restricted branching. Aust J Agric Res 54:649–661. doi:10.1071/AR02192
Gauch HG (1992) Statistical analysis of regional yield trials: AMMI analysis of factorial designs. Elsevier, Amsterdam
Gauch HG, Zobel RW (1997) Identifying mega-environments and targeting genotypes. Crop Sci 37:311–326
Gomez KA, Gomez AA (1984) Statistical procedures for agricultural research, 2nd edn. Wiley, New York
Harzic N, Huyghe C, Papineau J (1995) Dry matter accumulation and seed yield of dwarf autumn-sown white lupin (Lupinus albus L.). Can J Plant Sci 75:549–555
Harzic N, Huyghe C, Papineau J, Billot C, Esnault R, Deroo C (1996) Genotypic variation of seed yield and architectural traits in dwarf autumn-sown white lupin. Agronomie 16:309–316. doi:10.1051/agro:19960504
Huyghe C (1997) White lupin (Lupinus albus L.). Field Crops Res 53:147–160. doi:10.1016/S0378-4290(97)00028-2
Huyghe C, Papineau J (1990) Winter development of autumn-sown white lupin: agronomic and breeding consequences. Agronomie 10:709–716. doi:10.1051/agro:19900902
IRRI (2008) Cropstat for Windows Version 5. International Rice Research Institute, Los Baños, Philippines
Jensen ES, Hauggaard-Nielsen H (2003) How can increased use of biological N2 fixation in agriculture benefit the environment? Plant Soil 252:177–186. doi:10.1023/A:1024189029226
Joernsgaard B, Christiansen JL, Kuptsov N 2004 Adaptation of lupins for northern European maritime conditions. In: Proceedings of 10th International Lupin Conference, Int. Lupin Assoc., Canterbury, NZ, pp 105–110
Julier B, Huyghe C, Papineau J, Milford GFJ, Day JM, Billot C, Mangin P (1993) Seed yield and yield stability of determinate and indeterminate autumn-sown white lupins (Lupinus albus) grown at different locations in France and UK. J Agric Sci Camb 121:177–186
Lagunes-Espinoza L, Huyghe C, Papineau J, Pacault D (1999) Effect of genotype and environment on pod wall proportion in white lupin: consequences to seed yield. Aust J Agric Res 50:575–582. doi:10.1071/A98151
Lopez-Bellido L, Fuentes M (1990) Growth, yield and yield components of lupin cultivars. Agron J 82:1050–1056
Metayer N (2004) Vicia faba breeding for sustainable agriculture in Europe—identification of regional priorities and definition of target genotypes. GIE Févérole, Paris
Papineau J, Huyghe C (2004) Le lupin doux protéagineux. Editions France Agricole, Paris
Perini L, Beltrano MC, Dal Monte G, Esposito S, Caruso T, Motisi A, Marra FP (2004) Atlante agroclimatico—agroclimatologia, pedologia, fenologia del territorio italiano. UCEA, Rome
Perry MW, Dracup M, Nelson P, Jarvis R, Rowland I, French RJ (1998) Agronomy and farming systems. In: Gladstones JS, Atkins C, Hamblin J (eds) Lupins as crop plants: biology, production and utilization. CABI, Wallingford, pp 291–338
Petterson DS (1998) Composition and food uses of lupin. In: Gladstones JS, Atkins C, Hamblin J (eds) Lupins as crop plants: biology, production and utilization. CABI, Wallingford, pp 353–384
Piepho H-P (1995) Robustness of statistical tests for multiplicative terms in the additive main effects and multiplicative interaction model for cultivar trials. Theor Appl Genet 90:438–443
SAS (1999) SAS/STAT User’s guide, Version 8. SAS Institute, Cary, NC
Siddique KHM, Walton GH, Seymour M (1993) A comparison of seed yields of winter grain legumes in western Australia. Aust J Exp Agric 33:915–922. doi:10.1071/EA9930915
Stoddard FL, Balko C, Erskine W, Khan HR, Link W, Sarker A (2006) Screening techniques and sources of resistance to abiotic stresses in cool-season food legumes. Euphytica 147:167–186. doi:10.1007/s10681-006-4723-8
Acknowledgments
The work was carried out within the project “Increase of protein feed production” funded by the Ministry of Agricultural and Forestry Policies of Italy. We thank Sandro Proietti and Lino Fraschini for excellent technical assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Annicchiarico, P., Carroni, A.M. Diversity of white and narrow-leafed lupin genotype adaptive response across climatically-contrasting Italian environments and implications for selection. Euphytica 166, 71–81 (2009). https://doi.org/10.1007/s10681-008-9836-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-008-9836-9