
Abstract
Aspen woodland is an important ecosystem in the western United States. Aspen is currently declining in western mountains; stressors include conifer expansion due to fire suppression, drought, disease, heavy wildlife and livestock use, and human development. Forecasting of tree species distributions under future climate scenarios predicts severe losses of western aspen within the next 50 years. As a result, aspen has been selected as one of 14 vital signs for long-term monitoring by the National Park Service Upper Columbia Basin Network. This article describes the development of a monitoring protocol for aspen including inventory mapping, selection of sampling locations, statistical considerations, a method for accounting for spatial dependence, field sampling strategies, and data management. We emphasize the importance of collecting pilot data for use in statistical power analysis and semi-variogram analysis prior to protocol implementation. Given the spatial and temporal variability within aspen stem size classes, we recommend implementing permanent plots that are distributed spatially within and among stands. Because of our careful statistical design, we were able to detect change between sampling periods with desired confidence and power. Engaging a protocol development and implementation team with necessary and complementary knowledge and skills is critical for success. Besides the project leader, we engaged field sampling personnel, GIS specialists, statisticians, and a data management specialist. We underline the importance of frequent communication with park personnel and network coordinators.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Importance of aspen in western mountains
Quaking aspen (Populus tremuloides; hereafter, aspen) woodland is an important community type in semi-arid mountains and woodlands of the western USA. Although they constitute only a small portion of the landscape, aspen communities represent extremely biologically diverse areas (Winternitz 1980; Rumble et al. 2001), second only to riparian ecosystems. Aspen decline, observed over the past few decades, has resulted in losses of vertebrate species, vascular plants (Campbell and Bartos 2001), and other organismal groups. Aspen is therefore considered an indicator of ecosystem integrity (Woodley 1993) and has been portrayed as a keystone community (Bartos 2001). Aspen woodlands are highly valued by various human interest groups, including recreationists, artists, and naturalists. Aspen in North American semi-arid mountains is rarely appreciated for its timber value due to poor harvesting economy; however, the plant communities produce high-quality forage for wildlife and livestock (Beck et al. 2006).
Aspen ecology
The life cycle of aspen in the mountains of the western USA is unique. Although aspen is a prolific producer of viable seed, conditions required for successful germination and establishment are rare (Kemperman and Barnes 1976; Romme 1982; Mitton and Grant 1996). Botanists have argued that significant sexual reproduction in aspen has not occurred in the western USA since the last glacial retreat (Einspahr and Winton 1976; McDonough 1985). Recent research, however, indicates that genetic variability within aspen clones is common, suggesting that sexual reproduction in xeric aspen may occur more frequently than previously suggested (Mock et al. 2008). An example of more recent successful establishment of aspen followed the severe fires in Yellowstone National Park in 1988, which apparently provided a “window of opportunity” of suitable substrate and climate conditions for seed germination (Romme et al. 2005). Although aspen clones are long lived, possibly thousands of years (Kemperman and Barnes 1976), individual stems normally live 100–150 years (Mitton and Grant 1996; Shepperd et al. 2001). Since sexual regeneration requires prolonged moist conditions and is thought to be rare for xeric aspen, it is possible that an aspen clone lost from the landscape will not regenerate from seed (Mitton and Grant 1996).
Aspen stressors
Aspen stands in the western mountains commonly occur in conjunction with conifer species but have also been observed as uneven-aged aspen stands where aspen appears to persist as a stable, self-regenerating ecosystem (Rogers et al. 2010). These stable aspen systems are unsuitable for conifers or are located far away from conifer seed sources (Mueggler 1989). In biophysical settings where aspen is seral to conifer species, slow-growing shade-tolerant conifers begin to overtop aspen late in succession, eventually outcompeting the aspen, leading to aspen loss (Shepperd et al. 2001).
Browsing by wildlife and livestock has been shown to inhibit successful regeneration in aspen stands (Bartos and Campbell 1998; Kay and Bartos 2000; Kaye et al. 2005; Jones et al. 2009). Growth of aspen suckers is reduced when browsing results in the removal of terminal leaders and large amounts of branch biomass (Jones et al. 2009). Consecutive years of mid-season browsing and repeated browsing in the same growing season should be avoided in areas where aspen regeneration is a management goal (Jones et al. 2009). Aspen regeneration is particularly affected within elk (Cervus elaphus) winter range in areas when elk populations are high (Hart and Hart 2001).
Drought has been reported to cause mortality in aspen in Colorado (Worrall et al. 2008) and in the Canadian parklands (Brandt et al. 2003; Frey et al. 2004). In Colorado, mature stands on south-facing slopes at low elevations were found to be particularly susceptible to disease and insects as a result of acute drought and high temperatures. Aspen dieback in Canada has been correlated to factors such as stand age, drought and freeze-thaw events, defoliation, wood-boring insects, and fungal pathogens (Brandt et al. 2003; Frey et al. 2004). A relatively recent phenomenon affecting western aspen is referred to as sudden aspen decline (SAD) and is different from the slow decline in aspen populations that has been occurring over the past century. In cases of SAD, mature aspen stems are dying with no apparent regeneration in the form of suckering. Scientists and managers are currently researching the causes and trying to find solutions to this sudden die-off. Rising temperatures and drought have been correlated with this increased forest mortality in the southwestern USA (van Mantgem and Stephenson 2007; van Mantgem et al. 2009; Huang and Anderegg 2012), potentially caused by hydraulic failure of roots (Anderegg et al. 2012). Climate modeling predicts that in 50 years, approximately 40 % of aspen stands currently in the western USA will no longer have a suitable climate regime to support their existence (Rehfeldt et al. 2009), due to moisture stress and the balance between temperature and precipitation throughout the year.
Rationale of monitoring aspen in the upper Columbia basin
The Upper Columbia Basin Network (UCBN) within the U.S. National Park Service has identified aspen as 1 of 14 priority vital signs to monitor (Garrett et al. 2007). Information gained from monitoring aspen will contribute to the weight of ecological knowledge about natural resources and contribute to regional management strategies for the conservation of aspen. Aspen monitoring is currently implemented in two southern Idaho parks, City of Rocks National Reserve (CIRO) and Craters of the Moon National Monument and Preserve (CRMO). These parks are situated in areas located near or below the precipitation threshold of 400 mm per year (Perala 1990), below which upland aspen does not persist long term. A changing climate may result in a reduced snow pack, which could affect the availability of water for aspen stands in the parks. Aspen in these parks may be one of the first species to respond to locally changing climate, and the response may be a decline in regeneration or a die-off of water-stressed clones. It is not currently known how aspen will react to a changing climate locally, and the trends observed, as part of this monitoring protocol, will contribute timely, valuable data to future aspen management in the western USA.
Objectives of ecological monitoring
The primary object of ecological monitoring is to determine changes in characteristics of one or more species or other site characteristics over a relevant time period (Archaux and Bergès 2008; Bonham 1989). Monitoring programs are an essential tool providing information to the land manager on whether the ecosystem is departing from a desired state, measuring the success of land management actions, and detecting the effects of disturbance and environmental change (Legg and Nagy 2006). Specific objectives must be carefully determined for monitoring to be successful; otherwise, the success of the monitoring process will be fortuitous at best (West 2003; Carlsson et al. 2005). To be effective, a monitoring program should address the needs of the land manager by providing both short-term and long-term information on the condition of the monitoring site (Holm et al. 1987).
Once the objectives are identified, the specific characteristics to measure and the sampling method can be determined. It is also important to consider the magnitude of change desired to detect. For example, if a 1 % change is considered biologically significant, then a method that provides data at this or a finer scale must be used (Bonham 1989). Generally, quantitative data are more likely to provide adequate accuracy than qualitative data, but the choice of quantitative data does not ensure success (Legg and Nagy 2006). When evaluating time series data, sampling precision is more important than accuracy (Gotfryd and Hansell 1985). In addition, the ecological monitoring questions often change over time (Wildi et al. 2004). Because monitoring requires several periods of data collection, it is tempting to use data whose intended purpose was different than that to which it is applied. There is no method that adequately serves multi-purpose monitoring objectives (Wildi et al. 2004).
Indicators, for example, vegetation cover and composition, are frequently used as measurable surrogates in monitoring programs to increase sampling efficiency (Godínez-Alvarez et al. 2009). Noss (1990) stated that indicators should be sufficiently sensitive to provide early information regarding change, occur over a broad geographical area, provide data over a wide spectrum of environmental change, not be critically sensitive to sample size, be relatively easy to collect and analyze, and exhibit low annual and residual variation relative to true change due to management or environmental change. Since no single indicator typically possesses all these qualities, a complementary set of indicators is used.
The ability of a statistical test to detect trend in time with nominal test size is related to the variability of the data, the magnitude of the trend, and the type I error level at which testing is conducted (Cohen 1988). Therefore, power analysis is essential when planning monitoring programs (Legg and Nagy 2006). Statistical power can be increased by increasing sample size; it can also be increased by changing other sampling design characteristics such as the size of the sampling unit, stratification of samples, and the use of permanent plots (Archaux and Bergès 2008; Legg and Nagy 2006). It is important to note that many ecological variables vary with changing spatial grain and temporal scale (Archaux and Bergès 2008).
Permanently located plots are commonly used to increase statistical power in the presence of site-to-site variability (Legg and Nagy 2006). Milberg et al. (2008) identified reasons why there may be disagreement in the data when doing repeated surveys. These include (1) inability to relocate the plot, (2) variation in the ability to consistently quantify the plot’s characteristics, (3) random failure to observe some species occurrences, (4) disagreement on species identity, and (5) variation in taxonomic skills of observers over time. Vittoz and Guisan (2007) observed that only 45–63 % of the species were consistently detected by all observers regardless of the sample size. However, most missed species had cover values that were less than 0.1 %. Obviously, the uncommon species present the greatest problem when sampling because the sample sizes are often inadequate for statistical analysis. Careful location and documentation of plot locations, proper training of monitoring personnel, and monitoring efforts focused on a subset of target species can help to mitigate these problems.
Most monitoring programs have financial constraints limiting the time that can be invested. A common monitoring conundrum balances monitoring fewer sites where more detailed and time-consuming data collection is required against monitoring more sites using less intensive data collection methods. This decision varies on a case-by-case basis and is subject to several factors, including (1) monitoring objectives, (2) the spatial scale and homogeneity of the area, and (3) the magnitude of change that is desired to be detected.
Monitoring protocol development
In this section, we outline a process for the long-term aspen monitoring protocol and implementation (Strand et al. 2009) within UCBN parks. The protocol includes standard operating procedures (SOPs), according to suggestions by Oakley et al. (2003), for (1) preparation for the field season, (2) training observers, (3) finding GPS waypoints, (4) locating and establishing sampling transects, (5) measuring vegetation along transects, (6) data management, (7) data summary, analysis, and reporting, (8) protocol revision, and (9) field safety. The protocol development process is summarized in a conceptual flow diagram describing the components: objectives, resource mapping, pilot data collection, sample design, and data management considerations (Fig. 1).

Process for development of the aspen monitoring protocol
Measurable objectives
The overarching programmatic goal of the UCBN aspen vital signs monitoring program is to obtain data to inform management decisions pertaining to the perpetuation of quaking aspen populations at the two parks, CIRO and CRMO, where aspen is an important component resource. Following Beck et al. (2010), we have summarized monitoring program design components in Table 1. The monitoring protocol addresses the following specific measurable objectives (Strand et al. 2009): (1) estimate current status and long-term trend in regeneration of aspen populations in the parks and in individual stands; (2) estimate status and trend in aspen abundance, as measured by stem density of live and dead trees; specifically, live aspen stems were divided into five size classes (Table 2); (3) estimate the status and trend in dead standing aspen stems; (4) estimate the status and trend of conifer stem density by size class (Table 2) within aspen stands.
The decision to focus on stem counts in different size classes of aspen and conifer species, including dead aspen stems, originated in two existing aspen monitoring protocols by Kilpatrick et al. (2003) and Jones et al. (2005). Aspen sprout (often referred to as suckers) survival is important for the continuous regeneration of aspen clones. Aspen stems that are beyond the reach of browsers but are not yet mature trees reaching the crown (1.5–5 m tall) are considered new regeneration into the aspen population (Bartos and Campbell 1998). Information about stem density in size classes therefore provides necessary ecological data to determine if aspen suckering and regeneration is adequate and if the clone is deteriorating, indicated by a decreasing trend in live stems and an increasing trend in dead stems.
Statistical sampling objectives include the following:
-
Obtain an estimated mean, \( \widehat{\mu} \), within ± 25 % of the true mean, μ, with 90 % confidence, for aspen stem density (stems/ha) of suckers (class I + II), regeneration (class III + IV), mature trees (class V), and dead stems (class VI) within aspen clones. We are using the sample mean to estimate the true mean, μ.
-
Detect with >80 % power (1-β) a change ≥25 % between any two sequential time periods (5 years) of mean live or dead aspen stem density estimates with a 90 % confidence.
Identification of stands using remote sensing, field reconnaissance, and GIS
The first step in the monitoring protocol development process was to identify aspen stands in the sampling frame, which we accomplished via a combination of image classification of recent aerial photographs, on-screen digitizing, and field reconnaissance. We chose to use aerial photos rather than satellite images because of the fine resolution (1 m pixels) of these photos. Most aspen stands in the area are 1 ha or less in size, which would be difficult to detect on commercially available Landsat satellite imagery (30 m pixels). In monitoring vegetation types that occur in larger patches, Landsat images would likely be the preferred choice of imagery. During these initial steps toward development of a monitoring protocol, personnel skilled in the use of geographic information systems (GIS) and remote sensing were essential. We allocated one field season to the identification and mapping of aspen stands and evaluation of the variability in stem density by size class. The number of personnel required will depend on the size of the study areas, but in general one GIS/remote sensing specialist, one plant ecologist, and at least one field assistant are recommended for completing the initial mapping of the aspen resources within the area of interest. In CIRO and CRMO, aspen stands identified via classification of aerial photographs were visited in the field to confirm classification accuracy. During field reconnaissance, we visited all stands classified as aspen and removed cover types mistakenly classified as aspen. Misclassified cover types included broadleaf shrubs (Prunus spp.) and willow (Salix spp.).
Importance of statistical power analysis
Pilot data necessary for development of a sound sampling design were collected in 4-m radius plots in 13 stands in the parks 1 year prior to protocol implementation. We used 4-m radius plots based on suggestions for assessment and monitoring of aspen developed by Kilpatrick et al. (2003). Similar to Kilpatrick et al. (2003), our protocol allows for variable radius plots, which would be particularly useful following vigorous suckering after a disturbance. To date, we have only used the 4-m radius. These pilot data provided information about stem count variability and growth rates. Measurements in each plot included GPS coordinates, a stem count of all aspen and non-aspen trees, and assignment of each stem into predetermined size classes (Table 2). With permission from park personnel, we estimated growth rates in aspen saplings by destructively sampling four stems in each park. These saplings were cut in 30-cm increments and the growth rings for each segment were counted to estimate growth rates.
Power analyses were conducted at the stand level and at the park level to determine (1) the number of plots per stand to estimate stem counts within size classes with a confidence of 0.9 and a power of 0.8 and (2) the power to detect a 25 % change in either direction over 5 years within each park. Following Elzinga et al. (1998), we computed the required number of 4-m plots (n) for detecting a 25 % difference in the true mean within aspen size classes within aspen stands as follows:
where n = sample size, s = sample standard deviation, Z a = standard normal distribution quantile at the type I error of 0.1, Z b = standard normal distribution quantile at the type II error rate of 0.2, d = minimum detectable change in stem count since the initial sampling in year 1, \( s=\left({s}_1\right)\left(\sqrt{\left(2\left(1-\rho \right)\right)}\right) \), s 1 = sampling standard deviation among sampling units at the first time period, and ρ = correlation coefficient between sampling unit values in the first and second time period.
Analysis of pilot data revealed that the standard deviation of stem counts was of the same magnitude as the mean for each stand; in the aspen regeneration class (class III + IV), this was around 600 stems per hectare. We computed the sample size required for an adequate statistical test at different levels of confidence and power for both temporary and permanent plots. In both cases, we estimated the number of plots necessary for a 25 % minimum change detection using 600 stems per hectare as the standard deviation between plots. In the case of permanent sampling units, we estimated the correlation between time periods as 0.8. The monitoring frequency may be adjusted in the future if the correlation between time periods is found to differ substantially from 0.8 and subsequent power analysis indicates inadequate temporal replication. In stands that are 2–3 ha in size, 14–15 plots are required to determine a change in mean stem density of 25 % with a confidence of 0.90 and a power of 0.8. In the case of temporary plots, the number of plots required for the same confidence and power is 60–70, which is practically unreasonable. The results from this power analysis helped guide us to select permanent sampling locations to maximize the statistical power and confidence while minimizing the sampling effort.
Spatial autocorrelation—considerations and solutions
We anticipated problems with pseudoreplication (sensu Hurlbert 1984) from spatial dependence between adjacent plots, which would lead to underestimation of standard errors. Plots located closer together are more alike than plots farther apart, so variance increases with distance between plots. The variance of the difference between outcomes at two locations can be plotted in a semi-variogram as a function of distance between plots (Cressie 1991). To develop such a semi-variogram, we collected data in four stands as part of the pilot study. We placed twenty 4-m radius plots at variable distances in each stand. The plots were placed in three transects with an equal distance between plots in each transect. The distance between plots and transects was allowed to vary depending on the size of the aspen stand. In each plot, we recorded the GPS coordinates at the center point and the stem counts within size class (Table 2). We estimated minimum distances of spatially independent plots by examining semi-variograms for each size class in pilot data and used these distances to design the transect and plot layouts (Cressie 1991). Semi-variogram diagnostics indicated that the variance increased up to about 15–25 m (Fig. 2) which suggested that plots that were at least 25 m apart could be considered spatially independent. Spatially independent plots enable unbiased estimates of precision in mean and trend estimates.

Examples of semi-variograms of aspen class I (a) and aspen class II (b). The semi-variograms indicate that the spatial dependence in stem count between plots level out at a distance of 15–25 m. Plots that are spaced >25 m apart can therefore be considered spatially independent
Sampling design
Sampling design and objectives were developed through a process that involved site reconnaissance, pilot data collection and analysis, and thoughtful consideration of park and network information requirements. The development team consisted of the protocol authors, as well as other UCBN information management staff and park interpretive staff. Personnel required for developing and implementing this protocol and their responsibilities are summarized in Table 3.
Aspen stands within UCBN served as the sampling unit. The sampling frame consisted of the aspen stands within CIRO and CRMO because of the sensitive aspen resources in these parks, see “Rationale of monitoring aspen in the upper Columbia basin” section. The sampled population, which was also the target population for monitoring, included all upland aspen stands located on public land within the park boundaries (Table 1).
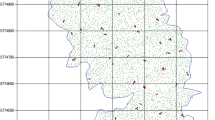
Aspen in a riparian corridor, short shrubby snow-damaged aspen, stands which were smaller than 0.3 ha in area, and aspen stands on private lands were excluded from the sampling frame. Aspen in the riparian corridor were excluded from sampling because they are less sensitive to climate change than upland aspen stands due to higher water accessibility. Snow-damaged aspen stands were excluded because they experience change due to mechanical damage induced by snow pack: stem density therefore cannot be expected to be a good indicator of change caused by climate conditions. One other stand was excluded from sampling because of difficult access and crew safety. Aspen stand perimeters were mapped and stored in a GIS (see example map for CIRO in Fig. 3). Notice that all delineated stands on the map (Fig. 3) were not sampled for the reasons described above.

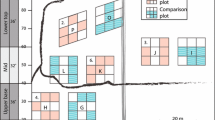
Stands and plots in the City of Rocks National Reserve
The number of plots within each aspen stand was determined via the power analysis. Following advice from statistics experts about random placement of plots while also considering field sampling limitations, we decided to place plots along transects with a random starting point. The random start location allows all parts of the stand to be available for sampling while placement along transects enables easier relocation of plots in the field. The random starting points were generated using the GIS software package ArcGIS (ESRI 2012) with transects oriented north–south or east–west depending on the axis of the stand. For small stands oriented in a different azimuth than north–south or east–west, transects were oriented along the longest axis of the stand. Four plots were placed along each transect with a distance of 25 m between plot centers (Fig. 4) as determined by the semi-variogram analysis. Altogether, 102 plots were placed within CRMO, arranged in 21 stands, and 122 plots were placed within CIRO, arranged in 26 stands. Logistically, we determined that it took the field crew 2 weeks for each sampling period to collect necessary data in these stands, a time period which was within the budget constraint for this project.

The start location for each transect is located in the center of plot 1 of the transect. Each plot along the transect is located 25 m apart, center to center. The direction of a transect is determined by the pre-defined azimuth (true north). Trees in different size classes (Table 2) are counted within each plots. The plot ID number is the stand number followed by the transect number followed by the plot number
Long-term monitoring by definition requires revisits of sampling locations at regular time intervals. Based on the power analysis of pilot data, we determined that permanently marked plot locations were necessary to detect change with the desired confidence and power. When determining the temporal sampling interval, the expected change between two sequential sampling periods must be considered. In the case of aspen stem counts of different size classes, we expected the largest temporal change in the smaller size classes. Pilot data indicated that 4–15 years was needed for an aspen seedling to reach a height of 150 cm. Therefore, a sampling interval of 5 years was deemed sufficient to detect change in aspen regeneration. In the developmental stages of this protocol (the first two time periods), we collected samples 3 years apart. Currently, we estimate a 5-year sampling interval for the final long-term protocol. The sampling interval will be reviewed by the UCBN team following each sampling period and may be adjusted up or down as determined necessary for adequate detection of change while conserving resources.
The monitoring protocol also requires a distance photo of each aspen stand to be obtained from a location with a good vantage point that overlooks the stand (see location of photo point in Fig. 3). The map coordinates, map datum, and true north azimuth in the direction the photo is oriented are recorded for repeatability.
Precision and reproducibility
Precision and reproducibility are important qualities in sampling design. Precision, defined as “the closeness of agreement between independent test results obtained under prescribed stipulated conditions,” was periodically assessed by each field team during each sampling occasion following recommended procedures outlined by Irwin (2008) and the Environmental Data Standards Council (2006). Duplicate counts of aspen and conifer stems were made periodically during sampling in order to provide estimates of precision, reproducibility, and measurement error. Specifically, each field team independently assessed relative percent difference (RPD) on 10 % of transects, which involved one duplicate density measurement taken by an alternate observer. Duplicate values were recorded in the data entry form for each transect’s RPD measure.
Data management
Strategic data management, critical to long-term monitoring, was led by the data management specialist on our protocol development team. Field data were collected on paper sheets or entered directly into a mobile field computer by the field crew. Data sheets (paper or digital) were inspected by team leaders and the project leader at the end of each field day, as a key step in the quality assurance and quality control process (QA/QC). Data from all paper data sheets were entered into the working copy of the UCBN Aspen database, a Microsoft Access application, as soon after data collection as practical; that entry was treated as an additional opportunity to conduct QA/QC. Validation rules programmed into the database help detect logical inconsistencies. The Microsoft Access database contains forms for simple data entry and reports summarizing the data are easy to produce and print. To reduce errors in data handling, data can be exported in custom reports that feed directly into scripts developed for statistical analysis.
What have we learned about aspen monitoring?
Aspen stands are variable and dynamic
Aspen stands are dynamic with high variability in suckering from year to year, in growth leading to change in size class, and in mortality occurring in all size classes. Sucker density varied by several thousand stems per hectare between time periods while the density in the regeneration class rarely varied more than 1000 stems per hectares (see summary statistics in Table 4). Stem counts for mature and dead classes were generally fewer and less variable between time periods (Table 4). Aspen stands were also observed to be spatially variable with suckering and regeneration occurring in one portion of the stand while non-existent in other portions. Conifer populations were encroaching on aspen in some parts of stands while conifers were completely absent in other parts of stands. Given this variability in both time and space, our sampling design (Strand et al. 2009) focused on distributing transects spatially among stands within the parks. We established a larger number of small sampling plots distributed across each stand rather than a single large plot within each stand. Following two time periods of repeated sampling, we have observed that each stand is unique in its dynamic between growth and presence of stressors. Initial observations suggest that conifer expansion is likely reducing aspen regeneration in a few stands, while wildlife browsing is the main stressor in other stands.
GPS considerations and relocation of plots
In a sampling design with permanent plots, such as in this protocol, the plot locations must be easy to relocate during subsequent sampling occasions. We used 50-cm-long rebar with a yellow plastic cap to mark the plot centers. The rebar was inserted into the ground with only the 3-cm-tall cap above ground. The marker locations were recorded using a high-accuracy Trimble GeoXT GPS unit with <1-m spatial accuracy after differential correction. The importance of using a high-accuracy GPS unit cannot be over-emphasized. We compared plots marked with the high-accuracy Trimble unit and a unit of lower accuracy (3–5 m). Permanent plot markers originally set with a lower accuracy GPS unit were extremely difficult to relocate 3 years later. Plots located with the lower accuracy unit also took much longer to find because of the larger search radius, and they were sometimes not found at all. Re-establishing plot monuments introduced error, particularly in this case, where the plots are relatively small (4-m radius) and in the same size range as the GPS error. We also determined that the lower accuracy GPS units are sufficient for locating permanent markers so long as the marker was originally recorded using a high-accuracy unit, hereby avoiding the compounding spatial error of both setting and searching for the plot using a low-accuracy GPS. Locating plots with GPS rather than stretching tapes between plot centers was much more time efficient. We tested both methods while designing the monitoring strategy. Overall, we achieved an 85 % plot center relocation rate, which would have been higher if we had initially used the high-accuracy GPS unit to monument plot centers. In general, we relocated plots within only a few minutes, although occasionally we searched as long as 15 min. Searching longer than 15 min rarely resulted in finding the plot. In cases where the plot marker was not found, we suspected that it had been removed by humans and animals or there had been a disturbance to the soil.
Field crew training
In all monitoring, personnel training is extremely important; aspen monitoring is no exception. During training, we found that observers were uncertain how to determine the size class of an aspen stem and if a stem should be classified as falling within the plot, live or dead, and standing or down. According to the standard operating procedures of this monitoring protocol (Strand et al. 2009), we found that the recommended half day of training was generally sufficient. During this half day of training, the trainers sample one plot while explaining each sampling step to the trainees. The trainees then sample several plots through the half day with the trainers available for guidance as needed. During training, the size breaks between aspen size classes and counting and recording of stems are important to emphasize. Clear communication between the plot reader and the data recorder is critical. Finally, we recommend that the project leader or an experienced ecologist or manager is always present when the permanent plots are set the first time. It is important that the permanent plot is set in a location that is within the aspen stand, not located in a stream or other unstable area, and otherwise within the constraints of the defined target population. Such long-term important decisions must be made by the monitoring the leadership team.
Database management and custom queries
Diligent field data handling and database management are of great importance. We recommend that field data be entered into the project master database each day if possible. Ideally, the project leader and the data manager review the data as soon as possible after it has been entered into the database. Many potential errors can be avoided by communication among the field crew members, project manager, and data manager. As time goes by, it is difficult for all parties to remember the plot, the data, and any issues during sampling that may shed light on apparent inconsistencies, outliers, and/or errors. Our aspen database has evolved over time, with the needs of the project leader, statisticians, and field crew directing revisions in database design and data summary output that were accomplished by the data manager. For example, the data manager has developed specific queries for the output of data in a format ready for summary tables and statistical analysis. These specific queries have been extremely helpful in reducing the risk of making errors during data handling, summarization, analysis, and reporting.
Statistical analysis
The aspen stem density data were analyzed by evaluating the difference in stand-level means for each age classification: suckers (seedlings), regeneration, mature trees, and dead trees. To test for evidence that the average change in stand level trees is non-zero, we used a non-parametric permutation test. Pairwise comparisons for each age class were performed with a blocked multi-response permutation procedure (blocked MRPP) with a Euclidean distance measure in the PC-ORD software (McCune and Grace 2002). MRPP compares the observed data to the expected distribution computed via the permutation procedure. Statistical results are interpreted via the “A” statistic of agreement and the p value. The A statistic is a measure of agreement between groups, where: A = 1 for complete within-group homogeneity; A = 0 when the heterogeneity within groups are equal to the expectation; and A < 0 if there is less agreement within groups than expected by chance (McCune and Grace 2002). The p value represents the probability of a type I error under the null hypothesis that there is no difference between groups and time periods in this analysis. We selected a non-parametric method because our data were not normally distributed. The overall change between the two time periods was computed, rather than a linear trend, since data existed for only two time periods at the time of the analysis. In the future, a methodology for trend analysis will be developed.
Summary statistics describing the average difference between the two sampling periods and standard deviation in aspen and conifer stem density within stands and by size class are provided in Table 4 for CIRO and CRMO. Summary statistics from the MRPP analysis are provided in Table 5 by park, species, and size class.
A significant increase in mature aspen stems (A = 0.123, p = 0.004) with a mean difference of 334 stems/ha was observed in CIRO, but no difference was detected in the other size classes. A significant increase in conifer seedlings (A = 0.064, p = 0.030), with a mean difference of 43 stems/ha, was also detected in CIRO. Although the analysis shows significance at the 0.05 alpha level, the agreement statistic A was low.
Statistical analysis indicated a significant (alpha = 0.05) increase in aspen regeneration with a mean difference in 750 stems/ha (A = 0.188, p = 0.002) between 2007 and 2010 in CRMO. No significant change was detected for suckers, mature, or dead trees. A significant increase in conifer seedlings (A = 0.073, p = 0.033), with a mean difference of 41 stems/ha, was also detected in CRMO. Similar to the observations in CIRO, the agreement statistic for the increase in conifer seedlings was low.
While stands may be considered somewhat independent within the park, repeated observations within each stand are not temporally independent. Thus, from a statistical viewpoint, based on only two points in time for each site and four calendar years total for all data, long-term trends cannot yet be assessed. Observed temporal changes may reflect either short-term variability or long-term temporal trends. Additional sampling data will help determine whether such temporal differences are indicative of long-term trends. The next sampling period begins in 2015. Scripts used for statistical analysis are stored as part of the datasets and will be revisited and updated for each sampling period.
Conclusions
The long-term aspen monitoring protocol fulfills the objectives of sound ecological monitoring as outlined in the introduction (Legg and Nagy 2006; West 2003; Carlsson et al. 2005). Monitoring objectives are clearly specified as suggested by West (2003), Carlsson et al. (2005), and many others. Our statistical design enables detection of disturbance and environmental change with necessary confidence and power and will not fall into the categories of protocols that are a “waste” of time, as pointed out by Legg and Nagy (2006).
We emphasize precision, as Gotfryd and Hansell (1985) state that sampling precision is more important than accuracy when evaluating time series data. Specific efforts to ensure precision in this protocol include (1) implementing permanent rather than temporary plot locations, (2) use of accurate GPS for relocating plots, (3) taking measures to avoid spatial autocorrelation between nearby plots by reviewing semi-variograms, (4) testing for relative percent difference in 10 % of the plots, (5) proper training of field crews, (6) diligent QA/QC, data entry, data storage, and data analysis, and (7) continuity in analysis methods through storage and documentation of statistical analysis for each time period.
Aspen monitoring addresses an immediate land management need as aspen is declining region-wide; short- and long-term monitoring results are made available to the land managers via the NPS database and reports produced after each monitoring event, as recommended by Holm et al. (1987). Data provided by this vital signs monitoring protocol will enable the NPS to engage in adaptive and proactive management with the goal of long-term preservation of aspen habitats within the parks. Monitoring aspen regeneration and stem density of live and dead aspen stems and conifers is important for determining when active management could be considered for restoration and long-term maintenance of aspen stands.
In the event that management action should become desirable for long-term aspen maintenance, Miller et al. (2005) emphasize the importance of asking the right questions before selecting a management technique. What is the desired future condition? What factors are affecting proper ecological function? What is the current state of the site? What is the predicted outcome of a treatment? A long-term monitoring program for detecting status and trends in aspen regeneration and population density at CIRO and CRMO will directly measure the population characteristics most important to park mission, visitor experience, and desired future conditions and will provide necessary data to provide feedback to adaptive management programs.
References
Anderegg, W. R., Berry, J. A., Smith, D. D., Sperry, J. S., Anderegg, L. D., & Field, C. B. (2012). The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proceedings of the National Academy of Sciences, 109(1), 233–237.
Archaux, F., & Bergès, L. (2008). Optimising vegetation monitoring. A case study in a french lowland forest. Environmental Monitoring and Assessment, 141(1–3), 19–25.
Bartos, D. L. (2001). Landscape dynamics of aspen and conifer forests. In W. D. Shepperd, D. Binkley, D. L. Bartos, T. J. Stohlgren, & L. G. Eskew (Eds.), Sustaining aspen in western landscapes: symposium proceedings; 13–15 June 2000; Grand Junction, CO (USDA Forest Service Proceedings RMRS-P-18) (pp. 5–14). Fort Collins: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Bartos, D. L., & Campbell, R. B. (1998). Decline of quaking aspen in the interior West—examples from Utah. Rangelands, 20(1), 17–24.
Beck, J. L., Peek, J. M., & Strand, E. K. (2006). Estimates of elk summer range nutritional carrying capacity constrained by probabilities of habitat selection. Journal of Wildlife Management, 70(1), 283–294.
Beck, J. L., Dauwalter, D. C., Gerow, K. G., & Hayward, G. D. (2010). Design to monitor trend in abundance and presence of American beaver (Castor canadensis) at the national forest scale. Environmental Monitoring and Assessment, 164, 463–479.
Bonham, C. D. (1989). Measurements for terrestrial vegetation. New York City: Wiley.
Brandt, J. P., Cerezke, H. F., Mallett, K. I., Volney, W. J. A., & Weber, J. D. (2003). Factors affecting trembling aspen (Populus tremuloides Michx.) health in the boreal forest of Alberta, Saskatchewan, and Manitoba, Canada. Forest Ecology and Management, 178(3), 287–300.
Campbell, R. B., & Bartos, D. L. (2001). Aspen ecosystems: objectives for sustaining biodiversity. In W. D. Shepperd, D. Binkley, D. L. Bartos, T. J. Stohlgren, & L. G. Eskew (Eds.), Sustaining aspen in western landscapes: symposium proceedings; 13–15 June 2000; grand junction, CO (USDA forest service proceedings RMRS-P-18) (pp. 299–307). Fort Collins: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Carlsson, A. L. M., Bergfur, J., & Milberg, P. (2005). Comparison of data from two vegetation monitoring methods in semi-natural grasslands. Environmental Monitoring and Assessment, 100(1–3), 235–248.
Cohen, J. (1988). Statistical power analysis for the behavioral sciences (2nd ed.). Mahway: Lawrence Erlbaum Associates.
Cressie, N. A. C. (1991). Statistics for spatial data. New York City: Wiley.
Einspahr, D. W., & Winton, L. L. (1976). Genetics of quaking aspen. USDA forest service research paper WO-25. Washington: U.S. Department of Agriculture, U.S. Forest Service.
Elzinga, C. L., Salzer, D. W., Willoughby, J. W., & Gibbs, J. P. (1998). Measuring and monitoring plant populations. Denver: U.S. Department of the Interior, Bureau of Land Management.
Environmental Data Standards Council. (2006). Quality assurance and quality control data standard. Standard No. EX000012.1. http://www.exchangenetwork.net/standards/qa_qc_01_06_2006_final.pdf. Accessed 13 October 2014.
Frey, B. R., Lieffers, V. J., Hogg, E. H., & Landhäusser, S. M. (2004). Predicting landscape patterns of aspen dieback: mechanisms and knowledge gaps. Canadian Journal of Forest Research, 34(7), 1379–1390.
Garrett, L. K., Rodhouse, T. J., Dicus, G. H., Caudill, C. C., & Shardlow, M. R. (2007). Upper Columbia basin network vital signs monitoring plan, (natural resource report NPS/PWR/UCBN/NRR—2006/001). Fort Collins: National Park Service.
Godínez-Alvarez, H., Herrick, J. E., Mattocks, M., Toledo, D., & Van Zee, J. (2009). Comparison of three vegetation monitoring methods: their relative utility for ecological assessment and monitoring. Ecological Indicators, 9(5), 1001–1008.
Gotfryd, A., & Hansell, R. I. (1985). The impact of observer bias on multivariate analyses of vegetation structure. Oikos, 45, 223–234.
Hart, J. H., & Hart, D. L. (2001). Interaction among cervids, fungi, and aspen in northwest Wyoming. In W. D. Shepperd, D. Binkley, D. L. Bartos, T. J. Stohlgren, & L. G. Eskew (Eds.), Sustaining aspen in western landscapes: symposium proceedings; 13–15 June 2000; grand junction, CO (USDA forest service proceedings RMRS-P-18) (pp. 197–205). Fort Collins: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Holm, A. M., Burnside, D. G., & Mitchell, A. A. (1987). The development of a system for monitoring trend in range condition in the arid shrublands of Western Australia. The Australian Rangeland Journal, 9(1), 14–20.
Huang, C. Y., & Anderegg, W. R. (2012). Large drought‐induced aboveground live biomass losses in southern rocky mountain aspen forests. Global Change Biology, 18(3), 1016–1027.
Hurlbert, S. H. (1984). Pseudoreplication and the design of ecological field experiments. Ecological Monographs, 54(2), 187–211.
Irwin, R. J. (2008). Draft Part B lite QA/QC Review Checklist for Aquatic Vital Sign Monitoring Protocols and SOPs. Fort Collins, CO: National Park Service, Water Resources Division. http://www.nature.nps.gov/water/vitalsigns/assets/docs/PartBLite.pdf. Accessed 13 October 2014.
Jones, B. E., Burton, D., & Tate, K. W. (2005). Effectiveness monitoring of aspen regeneration on managed rangelands. U.S. Department of Agriculture, Forest Service, Pacific Southwest Region.
Jones, B. E., Lile, D. F., & Tate, K. W. (2009). Effects of simulated browsing on aspen regeneration: implications for restoration. Rangeland Ecology and Management, 62(6), 557–563.
Kay, C. E., & Bartos, D. L. (2000). Ungulate herbivory on Utah aspen: assessment of long-term exclosures. Journal of Range Management, 53(2), 145–153.
Kaye, M. W., Binkley, D., & Stohlgren, T. J. (2005). Effects of conifers and elk browsing on quaking aspen forests in the central Rocky Mountains, USA. Ecological Applications, 15(4), 1284–1295.
Kemperman, J. A., & Barnes, B. V. (1976). Clone size in American aspens. Canadian Journal of Botany, 54(22), 2603–2607.
Kilpatrick, S., Clause, D., & Scott. D. (2003). Aspen response to prescribed fire, mechanical treatments, and ungulate herbivory. In P. N. Omi and. L. A. Joyce, (Eds.), Fire, fuel treatments, and ecological restoration: Conference Proceedings; April 16–18, 2002; Fort Collins, CO (Proceedings RMRS-P-29; pp. 93–102). Fort Collins, CO: U.S. Department of Agriculture, U.S. Forest Service, Rocky Mountain Research Station.
Legg, C. J., & Nagy, L. (2006). Why most conservation monitoring is, but need not be, a waste of time. Journal of Environmental Management, 78(2), 194–199.
McCune, B., & Grace, J. B. (2002). Analysis of Ecological Communities. MjM Software, Gleneden Beach, Oregon, USA. p. 304. www.pcord.com. Accessed 02 July 2015.
McDonough, W. T. (1985). Sexual reproduction, seeds and seedlings. In N. V. DeByle & R. P. Winokur (Eds.), Aspen: Ecology and management in the western United States (General Technical Report RM-119) (pp. 25–28). Fort Collins: U.S. Department of Agriculture, U.S. Forest Service, Rocky Mountain Forest and Range Experiment Station.
Milberg, P., Bergstedt, J., Fridman, J., Odell, G., & Westerberg, L. (2008). Observer bias and random variation in vegetation monitoring data. Journal of Vegetation Science, 19(5), 633–644.
Miller, R. F., Bates, J. D., Svejcar, T. J., Pierson, F. B., & Eddleman, L. E. (2005). Biology, ecology, and management of western juniper (Juniperus occidentalis) (Technical Bulletin 152). Corvallis: Agricultural Experiment Station, Oregon State University.
Mitton, J. B., & Grant, M. C. (1996). Genetic variation and the natural history of quaking aspen. Bioscience, 46(1), 25–31.
Mock, K. E., Rowe, C. A., Hooten, M. B., Dewoody, J., & Hipkins, V. D. (2008). Clonal dynamics in western North American aspen (Populus tremuloides). Molecular Ecology, 17(22), 4827–4844.
Mueggler, W. F. (1989). Age distribution and reproduction of intermountain aspen stands. Western Journal of Applied Forestry, 4(2), 41–45.
Noss, R. F. (1990). Indicators for monitoring biodiversity: a hierarchical approach. Conservation Biology, 4(4), 355–364.
Oakley, K. L., Thomas, L. P., & Fancy, S. G. (2003). Guidelines for long-term monitoring protocols. Wildlife Society Bulletin, 31(4), 1000–1003.
Perala, D. A. (1990). Populus tremuloides. In R. M. Burns & B. H. Honkala (Eds.), Silvics of North America: 2. Hardwoods; agriculture handbook 654 (pp. 555–569). Washington: U.S. Department of Agriculture, U.S. Forest Service.
Rehfeldt, G. E., Ferguson, D. E., & Crookston, N. L. (2009). Aspen, climate, and sudden decline in western USA. Forest Ecology and Management, 258(11), 2353–2364.
Rogers, P. C., Leffler, A. J., & Ryel, R. J. (2010). Landscape assessment of a stable aspen community in southern Utah, USA. Forest Ecology and Management, 259(3), 487–495.
Romme, W. H. (1982). Fire and landscape diversity in subalpine forests of Yellowstone National Park. Ecological Monographs, 52(2), 199–221.
Romme, W. H., Turner, M. G., Tuskan, G. A., & Reed, R. A. (2005). Establishment, persistence, and growth of aspen (Populus tremuloides) seedlings in Yellowstone National Park. Ecology, 86(2), 404–418.
Rumble, M. A., Flake, L. D., Mills, T. R., & Dykstra, B. L. (2001). Do pine trees in aspen stands increase bird diversity? In W. D. Shepperd, D. Binkley, D. L. Bartos, T. J. Stohlgren, & L. G. Eskew (Eds.), Sustaining aspen in western landscapes: symposium proceedings; 13–15 June 2000; grand junction, CO (USDA forest service proceedings RMRS-P-18) (pp. 185–191). Fort Collins: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station.
Shepperd, W. D., Bartos, D. L., & Mata, S. A. (2001). Above-and below-ground effects of aspen clonal regeneration and succession to conifers. Canadian Journal of Forest Research, 31(5), 739–745.
Software, E. S. R. I. (2012). ArcGIS Desktop v. 9 and 10. Redlands: Environmental Systems Research Inc.
Strand, E. K., Bunting, S. C., Steinhorst, R. K., Garrett, L. K., & Dicus, G. H. (2009). Upper Columbia basin network aspen monitoring protocol: narrative version 1.0 (natural resource report NPS/UCBN/NRR—2009/147). Fort Collins: National Park Service.
van Mantgem, P. J., & Stephenson, N. L. (2007). Apparent climatically induced increase of tree mortality rates in a temperate forest. Ecology Letters, 10(10), 909–916.
van Mantgem, P. J., Stephenson, N. L., Byrne, J. C., Daniels, L. D., Franklin, J. F., Fulé, P. Z., et al. (2009). Widespread increase of tree mortality rates in the western United States. Science, 323(5913), 521–524.
Vittoz, P., & Guisan, A. (2007). How reliable is the monitoring of permanent vegetation plots? A test with multiple observers. Journal of Vegetation Science, 18(3), 413–422.
West, N. E. (2003). Theoretical underpinnings of rangeland monitoring. Arid Land Research and Management, 17(4), 333–346.
Wildi, O., Feldmeyer-Christe, E., Ghosh, S., & Zimmerman, N. (2004). A note on vegetation monitoring approaches. Community Ecology, 5(1), 1–5.
Winternitz, B. L. (1980). Birds in aspen. In R. M. DeGraff & N. G. Tilghman (Eds.), Management of western forests and grasslands for nongame birds (general technical report INT-86) (pp. 247–257). Ogden: U.S. Department of Agriculture, U.S. Forest Service, Intermountain Forest and Range Experiment Station.
Woodley, S. (1993). Monitoring and measuring ecosystem integrity in Canadian national parks. In S. Woodley, J. Kay, & G. Francis (Eds.), Ecological integrity and the management of ecosystems (pp. 155–176). Del Ray Beach: St. Lucie Press.
Worrall, J. J., Egeland, L., Eager, T., Mask, R. A., Johnson, E. W., Kemp, P. A., & Shepperd, W. D. (2008). Rapid mortality of Populus tremuloides in southwestern Colorado, USA. Forest Ecology and Management, 255(3), 686–696.
Acknowledgments
Funding for this project was provided through the National Park Service Natural Resource Challenge and the Service-wide Inventory and Monitoring Program. We thank Dr. Kirk Steinhorst for statistical expertise and coordination and Gina Wilson for image processing of City of Rocks’ aerial photos. We thank the staff at the City of Rocks National Monument and Craters of the Moon National Monument and Preserve for contributions and critique of the aspen monitoring protocol and annual reports and for assistance during field reconnaissance. Thank you to the field assistants and summer interns that worked with us during the summers 2006–2012, you contributed immensely to field data collection, data recording, and fun.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Strand, E.K., Bunting, S.C., Starcevich, L.A. et al. Long-term monitoring of western aspen—lessons learned. Environ Monit Assess 187, 528 (2015). https://doi.org/10.1007/s10661-015-4746-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10661-015-4746-5