
Abstract
In controlled environments, the influence of rice growth stage, temperature, relative humidity and wetness duration on infection by Villosiclava virens (cause of rice false smut) was assessed by inoculating the rice cultivar ‘Yueyou938’ with a conidial suspension (2 × 106 spores) until run-off. Rice plants grown in vitro, were inoculated at a late stage of panicle development, and averaged 90 % diseased panicles which was a higher incidence compared with those inoculated at a mid-stage in panicle development (66.5 %). There were significant differences in the effects of temperature and wetness duration on disease incidence, which increased rapidly from 48 to 120 h at both 20 and 25 °C. The highest level of disease (92.9 %) was obtained at 25 °C and 95 % RH with 120 h of wetness. In another set of experiments, inoculated rice plants were incubated in a plant growth chamber at 25 °C and 95 % RH for a wetness period; of 120 h, and then subjected to different combinations of temperature and relative humidity to determine the effects of both factors on infection by V. virens. A higher temperature (28 °C) during incubation was found to induce a significantly higher disease incidence compared with a lower temperature (20 °C). Given a wetness period of at least 120 h, temperature was the primary factor affecting disease development during the 2 week period after that. Disease incidence was modeled as a function of both temperature and wetness duration using a modified version of the Weibull model. There was good agreement between observed and predicted values (R 2 = 0.951 and P = 0.0001). The model was used to construct a risk chart for estimating the potential for infection based on observed or forecast temperatures and wetness durations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rice false smut (RFS), caused by the ascomycete Villosiclava virens (anamorph: Ustilaginoidea virens), is common on panicles of rice in most rice (Oryzae sativa) producing areas of the world (Zhou et al. 2008). Infection by V. virens (which can be detected by nested PCR before development of visual symptoms (Zhou 2004)) results in grains on mature head being transformed into globose, yellow-green and velvety smut balls (Wang et al. 2008). Since the 1980’s, with changes in rice cultivation patterns, increased nitrogen fertilization and use of compact panicle types of hybrid rice, RFS has gradually become one of the most serious diseases on rice (Huang and Yu 2002; Lv et al. 2008; Wang et al. 2004). In 2004, an outbreak of RFS occurred in central China, with 1.37 billion kilograms of rice being lost (Wang and Chen 2005). Since the pathogen can produce ustiltoxin a and b which can harm livestock and humans, V. virens not only decreased the yield but also reduced the quality of the rice (Li et al. 1995; Nakamura et al. 1994).
Growth stage, temperature, relative humidity (RH) and wetness duration are critical factors for successful primary infection of rice by V. virens, and the effects of these factors have been studied extensively. Rice seedling stage was once used as the basis for timing of susceptibility for infection by V. virens (Chen et al. 1995; Ikegami 1962). However, after inoculation experiments with conidial suspensions in vitro, infection was inferred to occur during the rice booting stage (Gao and Zhong 2011; Li et al. 1986; Lu et al. 1996). Artificial inoculation at different rice growth stages has shown the period of 6 to 9 days before rice heading is the optimal time for infection with this pathogen. The most suitable temperature range for growth of V. virens mycelium is 25 to 28 °C (Ma et al. 2001), and for germination of conidia it is 22 to 31 °C (Zhang et al. 2003). The germination rate reaches a maximum at 28 °C but shows a sharp decrease below 25 % RH (Zhang et al. 2003). A temperature of 16 °C was found to induce a higher disease incidence compared with other temperatures (Wang et al. 2008), and 5 days of wetness at 100 % RH was found to be the optimal wetness duration for infection (Fujita et al. 1989). However, the combined interactive effects of temperature and wetness duration on the infection by V. virens under controlled conditions have not been characterized.
A Weibull model may provide a good description of disease incidence response to temperature and wetness duration. In contrast to some other models, e.g. the exponential, monomolecular, or logistic, the Weibull model can incorporate curves of some other models by adjusting the value of the shape parameter, and this model has been widely used in many fields of plant disease epidemiology research (Carisse et al. 2000; Tan 1991; Wang et al. 1987). Wu et al. (1999) inoculated three peanuts cultivars under controlled environments with varying temperature and wetness duration, and modelled severity of Cercospora leaf spot using the Weibull model. With the same model, Carisse et al. (2000) determined the combined effects of temperature and wetness duration on infection by Mycosphaerella fragariae, and built a disease risk forecasting model to guide fungicide application in strawberry production.
The objectives of this work were: (i) to determine the effects of panicle development stage (PDS), temperature, wetness duration and relative humidity on disease incidence induced by V. virens in controlled environments; and (ii) to quantify the combined effects of temperature and wetness duration on disease incidence based on a nonlinear model, in order to improve the efficiency of the artificial inoculation protocol for other research purposes, including evaluation of rice resistance against RFS.
Materials and methods
Inoculum preparation and plant cultivation
A mono-conidial isolate of V. virens, HWD-2, was obtained using the tissue separation method (Fang 1998) from a naturally infected rice panicle in 2009, in Wulijie District, Wuhan City, Hubei Province, China. This virulent isolate had higher conidial production (approximately 1 × 107 spores per ml) compared with other isolates (averaging 1 × 106 spores per ml), and was used for the inoculation tests. Isolate HWD-2 is stored at the China Center for Type Culture Collections (CCTCC NO: 2011023) in Wuhan, China. To produce sufficient quantities of conidia for artificial inoculation, a 6-mm-diameter mycelial plug incubated on potato sucrose agar (PSA, 2 % sucrose plus extract from boiled fresh potatoes) for 7 days was placed in 150 ml sterilized potato sucrose broth (PSB) in a 250 ml flask which was placed in a shaking incubator (HZ300L, Wuhan Ruihua Appliance Limited Company, China) at 28 °C in the dark and 180 rpm for 7 days. The cultures were filtered through four layers of gauze, and conidia were obtained from the filtrate by centrifugation (3,000 rpm for 5 min, TGL-10B, Wuxi Ruijiang analytical instrument Limited Company, China). The conidial suspension was adjusted to 1 × 106 conidia per ml using a haemocytometer (XB.K.25, Shanghai Qiujing Biochemical reagent and Instrument Limited Company, China).
The RFS-susceptible rice cultivar used for inoculations‘Yueyou938’ (Zhang et al. 2006), is a type of three-line var. indica hybrid rice, derived from YuetaiA × R938, with a 158-day growth period, a stem length average of 114 cm and an average of 162 grains per panicle. Seeds were treated for 24 h with 0.1 to 0.15 % potassium permanganate solution and soaked in water for 5 days. The seeds were incubated at 30 °C for 24 h before sowing. Four rice seedlings were transplanted into individual plastic buckets (25 × 20 × 30 cm) containing 5 kg paddy soil autoclaved at 121 °C for 1 h and placed in a lathe-house. Before the inoculations at the booting stage (approximately 90 days after sowing), rice plants were fertilized (4 g carbamide in each bucket) once at tillering (approximately 45 days after sowing) and once at booting, and treated with insecticide only as necessary (no fungicide). Irrigation was applied every other day.
Inoculation and environmental conditions
We divided the booting stage into eight panicle development stages (PDS) (Ding, 1961). To place PDS in accordance with different distal internode lengths (DIL), a preliminary experiment was carried out before the inoculation tests. Anthers in florets on the middle section of panicles of cv. Yueyou938 were chosen for observation throughout the booting stage with a DIL range of 1 to 17 cm and examined at 1 cm intervals. For each DIL, five panicles were collected, and three florets were collected from each panicle. The samples were examined by microscopy (LX71, Olympus Corporation, Japan) at a magnification of 200 using the simple squash method (Yang and Zhou 1979; Yang and Dai 2009). Based on the descriptions of Ding (1961), the morphology of pollen cells was characterized, and a pairwise link between DIL and PDS was established.
-
Experiment 1
To examine the effect of PDS on disease incidence, and to confirm optimal PDS for artificial inoculation efficiency, panicles differing in DIL, and hence belonging to each of the different PDS groups, were inoculated by a syringe with 2 ml of conidial suspension (2 × 106 spores) in the middle section of the distal internodes. The rice plants were placed in a plant growth chamber (ZSX1500GS, Wuhan Ruihua Appliance Limited Company, China) equipped with a high pressure sodium lamp (12 h light/dark cycle) with conditions set at a RH of 95 ± 5 % and a temperature of 25 ± 1 °C. After a post-inoculation surface wetness period of 120 h, three buckets of rice plants of each PDS treatment were transferred to a greenhouse equipped with an automatic climate control system (AUTO-2000, Beijing, China) set at 28 ± 2 °C and 90 ± 7 % RH. This experiment was repeated three times.
-
Experiment 2
To study the effects of temperature and wetness duration on disease incidence, rice plants were inoculated with 2 ml of conidial suspension (2 × 106 spores) using a syringe in the middle section of the distal internodes at PDS 7. The plants were placed in chambers, each set at a different temperature (15 ± 1, 20 ± 1, 25 ± 1, 30 ± 1, or 35 ± 1 °C) and all on a 12-h light-and-dark cycle, with RH of 95 ± 5 % to ensure panicle wetness. After post-inoculation wetness periods of 12, 24, 48, 72 or 120 h, the rice plants at each temperature treatment were transferred to a greenhouse as described for experiment 1. The experiment was conducted using a split plot design where the main plots were temperature and subplots were wetness duration. This experiment was repeated three times, and each experimental run was treated as a block. In order to minimize variation among treatments, rice plants were randomly selected for inoculation, and distributed in chambers for incubation at different temperatures and wetness durations.
-
Experiment 3
To study the effects of post-inoculation cultivation conditions on disease incidence, and using the same inoculation and humidity conditions as in experiment 1 (25 ± 1 °C and 95 ± 5 % RH for 120 h), rice plants were transferred into four different greenhouses and were exposed to different combinations of temperature (20 ± 2 or 28 ± 2 °C) and relative humidity (75 ± 7 or 90 ± 7 % RH). This experiment was repeated three times.
For each experiment (1 to 3), rice panicles, which were marked by paper tags on each flag leaf, were inoculated with 2 ml of conidial suspension (2 × 106 spores) by syringe injection until runoff. Based on previous studies (Tan 2008), there was a strong correlation between disease incidence and other indices of RFS. After a further 2 weeks of growth, disease incidence (=number of diseased panicles / number of inoculated panicles × 100 %) was calculated from the results of the three independent experiment repetitions. In each experimental repetition, disease incidence was an average of observations made on ~60 panicles (three buckets of plants per replication, ~20 panicles in each bucket). Since smut balls can appear over a period of a few days, the disease incidence was evaluated at 18 and 21 days after inoculation to ensure smut balls were counted accurately.
Model development
Development of the model was done with PROC NLIN, PROC G3D and PROC GCOUNTRER in SAS 9.2. Following the method described by Carisse et al. (2000), a surface response model was developed to describe the effects of temperature and wetness duration on disease incidence in experiment 2. In brief, Eq. 1, proposed by Duthie (1997), explained incidence response as a function of wetness duration
where Y is the disease incidence in response to duration of wetness, A is the upper limit of the disease incidence, B is the intrinsic rate of increase, C is the minimum wetness duration for the response, D is the point of wetness duration where the incidence value decelerated, and w is the wetness duration (Carisse et al. 2000).
First, Eq. 2, which represented the maximum disease incidence in response to temperature (T), was used to calculate the upper asymptote (A)
where A is the upper asymptote of disease incidence in response to temperature, E’ is equal to E[(H + 1)/H]H 1/(H + 1), E is the maximum incidence value, F is the temperature at which the incidence value attained a maximum, G is the intrinsic rate of incidence declining from the maximum, and H is the parameter describing asymmetry of the curve. In order to estimate parameters E, F, G, and H, partial derivative equations and the Marquardt iterative method were used during the NLIN procedure in SAS. On the assumption that infection responded immediately to increases in wetness duration after a minimum of 12 h, the parameter C in Eq. 2 was fixed at 0 to avoid over-parameterization. For the estimation of parameters B and D, the same method of the NLIN procedure described above was used. Finally, the asymptote (A), the parameters B and D and the value C (assumed to be 0) were included in Eq. 1, which was used to predict disease incidence at a given temperature and wetness duration (Carisse et al. 2000).
Data analysis and model evaluation
Homoscedasticity (homogeneity of variance) of the response variables in the different experiments was tested using PROC ANOVA in SAS 9.2 (SAS Institute Inc., Cary, NC) to ensure that the assumption of normality was maintained before analysis of the data in each experiment. The effects of PDS, temperature, wetness duration and relative humidity on disease incidence were evaluated based on the results of three independent experimental repetitions. Separation of means was tested using the Least Significant Difference (LSD) test at α = 0.05.
To assess the appropriateness of the models, certain criteria had to be met: (i) the residuals should be randomly and normally distributed; and (ii) a good fit was required between predicted disease incidence and observed values. To evaluate the nonlinear regression, a linear regression analysis of the predicted disease incidence and the observed values was performed. The evaluation criteria for the regressions were a t-test to determine whether the intercept of the regression curve was significantly different from 0, and to determine whether the calculated slope was different from 1, the coefficient of determination (R 2) for the regression solution, and the F-value of the model (which had to be significant at P = 0.05) (Dent and Blackie 1979).
Results
Effect of panicle development stage (PDS) on disease incidence
In the preliminary experiment, rice plants with different DIL were classified into four groups; PDS 5 with DIL 1–5 cm, PDS 6 with DIL 6–10 cm, PDS 7 with DIL 11–15 cm, and PDS 8 with DIL 16–17 cm (Table 1). Pollen mother cells had formed by PDS 5 (Fig.1a). At PDS 6, pollen mother cells were undergoing meiosis, and producing dyads and tetrads (Fig. 1b). At PDS 7, tetrad cells developed from monokaryons to dikaryons which were full of inclusions (Fig. 1c). At PDS 8, pollen cells developed into trikaryons which are associated with mature pollen (Fig. 1d). Thus rice plants for experiment 1 were classified into 4 groups, and used to evaluate the optimal timing for inoculation. The plants inoculated at PDS 5 showed no smut balls while the plants inoculated at PDS 6 to 8 showed the same type of symptoms as those observed in the field, i.e. yellow smut balls replacing the kernels in the heads. In experiment 1, the results of three repetitions showed a consistent trend in disease incidence across PDS (Table 1), where PDS values of 7 to 8 had higher disease incidence (more than 90 %) compared with the earlier stages (PDS 5 and 6), and were thus considered the optimal stage for inoculation.

The morphology of pollen cells at panicle development stages (PDS) 5 to 8. a Pollen cells at PDS 5, b Pollen cells at PDS 6, c Pollen cells at PDS 7, d Pollen cells at PDS 8. a–c are images of pollen cell tissue stained by carbol fuchsin (Sigma), d is an image of pollen cell tissue stained by hemataxylin (Sigma). p pollen mother cell, d dyad, t tetrad, mk monokaryon, dk dikaryon, tk trikaryon
Effect of temperature and panicle wetness duration on disease incidence
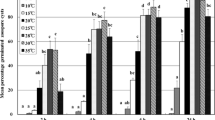
The mean RFS incidence for each treatment was similar (neither significant block effects nor significant experiment × treatment interactions) in the three repeats of experiment 2. However, temperature, wetness duration and the interaction (temperature × wetness duration) significantly affected mean RFS incidence (Table 2). At 20 and 25 °C, there was significant variation among mean RFS incidence induced by five different wetness durations (12, 24, 48, 72, 120 h). At the other temperatures tested (15, 30, or 35 °C), the mean RFS incidence induced by five different wetness durations (12, 24, 48, 72, 120 h) did not always show significant separation from each other, e.g. disease incidence at 35 °C for 12, 72 and 120 h of wetness were similar to each other (Fig. 2). The highest incidence of RFS-infected panicles was obtained at 25 °C and 95 % RH for 120 h of wetness (92.9 %). The lowest disease incidence (5 %) was observed on the plants exposed to 35 °C for 120 h of wetness (Fig. 2).

Rice false smut incidence on plants inoculated with 2 ml of conidial suspension (2 × 106 spores) of Villosiclava virens and incubated under various combinations of temperature (15, 20, 25, 30, 35 °C) and wetness duration (12, 24, 48, 72, 120 h). HPI is the hours of wetness post inoculation. Each bar height is an average of observations made on ~180 panicles (three experimental repetitions with three buckets of plants per repetition, ~20 panicles in each bucket). At each temperature, bars sharing the same letter are not significantly different according to a Least Significant Difference test (α = 0.05)
The upper asymptote (maximum RFS incidence of 98.9 %) was at 23.8 °C and the lowest (2.5 %) at 35 °C. The relationship between the upper asymptote and temperature was described as follows
where E′ = 0.9891 × [(2.2866 + 1)/2.2866] × 2.28661/(2.2866 + 1) Parameter values and corresponding errors are listed in Table 3. Regression of the observed upper asymptote against the predicted value showed the intercept was not different from 0 (P > 0.05), but the slope was significantly different from 1 (P < 0.01). The model F-test was highly significant (P < 0.01), and the associated coefficient of determination (R 2) was 0.996 (Fig. 3).

Regression of the predicted values (Y) calculated from Eq. 3 (see text) against observed values (X) of the maximum rice false smut incidence (=number of diseased panicles / number of inoculated panicles × 100 %) induced by Villosiclava virens based on observations made at different temperatures (15, 20, 25, 30, 35 °C) levels. Coefficient of determination R 2 = 0.996 (P = 0.0001)
The surface response (Fig. 4) describing the relationship between the combined effects of temperature and wetness duration on RFS incidence was obtained by merging Eqs. 3 and 1 with C = 0.

Three-dimensional response surface of the predicted incidence of rice false smut caused by Villosiclava virens at temperatures and wetness durations, where the predicted disease incidences (=number of diseased panicles / number of inoculated panicles × 100 %) were calculated from Eq. 4 (see text). As predicted by Eq. 4, the maximum disease incidence of the rice plants inoculated at PDS 7 was 90.9 % under optimal incubation conditions of 23.8 °C and 95 % RH for 120 h
where parameter A was calculated from the results of Eq. 3. For all temperatures and wetness durations, a random distribution was found in the residual plots analysis, and the model gave a precise prediction of RFS incidence for all wetness durations. The regression of the observed RFS incidence against the predicted value was not significantly different from 0 (P > 0.05), but the slope was highly significantly different from 1 (P < 0.01) (Table 4). The model F-test was highly significant (P < 0.01) and the coefficient of determination (R 2) was 0.951 (Fig. 5).

Regression of the predicted values (Y) calculated from Eq. 4 (see text) against observed values (X) for the incidence of rice false smut (=number of diseased panicles / number of inoculated panicles × 100 %) caused by Villosiclava virens based on observations made at different combinations of temperature (15, 20, 25, 30, 35 °C) and wetness duration (12, 24, 48, 72, 120 h). Coefficient of determination R 2 = 0.951 (P = 0.0001)
On the basis of the model (Eq. 4), the optimum temperature from the artificial inoculation experiments was predicted to be 23.8 °C. The minimum wetness duration for appearance of disease symptom was 12 h. If wetness duration was fixed, the disease incidence gradually increased with the rise of temperature and declined rapidly after RFS incidence reached its maximum value at 23.8 °C. If a constant temperature value was used in the model, the RFS incidence gradually increased with more prolonged wetness, and attained a maximum value after 120 h of wetness. In general, the model overestimated RFS incidence at temperatures of 15, 20 and 25 °C and wetness durations of 24 or 48 h. It also overestimated disease incidence at a temperature of 30 °C and wetness durations of 48 or 72 h by up to 39.6 %. But for the remaining treatments at different combinations of temperature and wetness duration, the model tended to underestimate RFS incidence by 36.2 % on average.
Since a long period of wetness can induce severe RFS, a contour map (Fig. 6) was developed for analysis of variable infection efficiency in a controlled environment. If the temperatures were too low (≤7 °C) or too high (≥33 °C), the RFS incidence will be low (not more than 5 %) regardless of the wetness duration (Eq. 4). However, RFS incidence could attain a high value (greater than 50 %) if the wetness duration exceeded 48 h with a moderate temperature (22 to 25 °C).

Risk prediction of rice panicle smut caused by Villosiclava virens. Categories are based on the predicted rice false smut (RFS) incidence (=number of diseased panicles / number of inoculated panicles × 100 %) calculated by Eq. 4 (see text). The separate categories correspond to Light RFS incidence (0 to 5 %) representing conditions related to low efficiency inoculation; Moderate RFS incidence (5 to 80 %) representing conditions for medium efficiency; and Severe RFS incidence (80 to 100 %) representing conditions for high efficiency
Effects of post-inoculation cultivation conditions on disease incidence
After 120 h of wetness, rice plants, cultivated in four greenhouses set for temperature and relative humidity combinations of 28 °C and 75 % RH, 28 °C and 90 % RH, 20 °C and 75 % RH, or 20 °C and 90 % RH for more than 2 weeks, showed mean RFS incidence of 25.1 to 92.7 % (Table 5). There was a significant (P < 0.05) difference in mean RFS incidence at different temperatures when relative humidity was the same, while there was no significant (P > 0.05) difference in mean RFS incidence at different relative humidity levels when temperature was the same. Thus, higher incidence of RFS may result from rice cultivation under conditions at 28 °C compared with at 20 °C.
Discussion
Currently, there are two main methods for the timing of inoculation of rice with V. virens. One method relies on inoculation 6 to 9 days before heading of rice plants (Zhang et al. 2004), and the other when the DIL of the rice panicles is 2 to 12 cm (Wang et al. 2008). Because different rice cultivars and plants grown under different conditions may not share the same DIL or heading times, the methods based on heading time or DIL may not be reliable or easily distinguishable under different conditions. Thus we developed an alternative method to optimize inoculation time for improvement of inoculation protocols, including the growth stage of the rice plant.
The optimum inoculation time was found to be PDS 7 to 8 which is equivalent to 3 to 5 days before heading for rice cultivar Yueyou938, when the DIL was 11 to 17 cm. A possible reason for the discrepancies in choice of the timing of inoculation between the present study and previous studies is perhaps that different rice cultivars may not have the same heading stage or a consistent DIL during the same physiological period. This new method, which involves inoculation times based on PDS, allows a more precise estimate of the optimal time for inoculation with V. virens considering the development stage of the rice panicle, but further study with other isolates and other cultivars is needed to confirm this result.
In the present study, a DIL of 1 to 17 cm corresponded to PDS 5 to 8 for Yueyou938 (Ding 1961). The PDS of rice can be represented by DIL, but the relationship between the DIL and PDS was not previously stated explicitly in the literature. By more precisely timing the optimal inoculation period, some of the variation in inoculation efficiency can be removed. Rice plants at PDS 1 to 4 were not used for inoculation in this experiment because DIL of the panicle at PDS 1 to 4 were negative values and could not be discriminated by eye. The assessment of the optimal period for inoculation should be tested during the relevant developmental stage of the host plant i.e. PDS 7 to 8. Another interesting observation in this study was that the conidial suspension caused inhibition of young panicles from heading at early stages, confirming previous observations by Zhang et al. (2004).
Temperature and wetness duration are two factors which affect infection by V. virens, so these factors should be considered first in studies requiring inoculation. As Fujita et al. (1989) found, a maximum disease incidence of 83.3 % could be obtained at the optimal conditions of 26 °C and 100 % RH for 5 days subsequent to a preliminary inoculation treatment at 15 °C and 90 % RH for 2 days. However, a precise temperature and relative humidity cannot easily obtained in many greenhouses, so the primary task should be to provide wider yet optimal ranges for artificial inoculation, not only specific values for optimal environmental conditions.
The Weibull model was constructed to quantitatively depict RFS incidence as affected by temperature and wetness duration. The derived Weibull model gave a good fit between the observed RFS incidence and the predicted values calculated using the final equation (the coefficient of determination = 0.951). Moreover, the contour map (Fig. 6), developed based on the Weibull model, showed us ranges of environmental conditions that were favorable to a high efficiency of inoculation with V. virens. Temperatures (22 to 25 °C) with no less than 48 h of wetness duration were considered to be environmental conditions necessary for effective inoculation (with the resulting RFS incidence no less than 50 %). These results may provide an important guideline for selection of inoculation conditions in a greenhouse.
Tang et al. (2013) observed that the pathogen grew out of the spikelet 10 days after inoculation and the smut balls formed 15 days after inoculation. During the latent period, incubation conditions after inoculation may have an effect on development of V. virens. By artificial inoculation with a conidial suspension, growth conditions at relative high temperatures (25 to 35 °C) were found to be more suitable for development of RFS symptoms caused by V. virens compared with at 15 °C (Fujita et al. 1989). Observations from the present study are consistent with the previous results. More severe RFS was induced by a favorable temperature range (26 to 30 °C) in the later period of growth along with sufficient wetness.
Beyond the factors mentioned above, other environmental conditions may affect infection of rice by V. virens and the subsequent development of RFS. Although light treatments have no known influence on either promoting or inhibiting the germination of chlamydospores of V. virens, light can inhibit the formation of secondary spores from chlamydospores (Wang 1988). Nitrogen fertility is also thought to affect incidence of RFS (Brooks et al. 2009). Savary et al. (1995) reported that high levels of nitrogen fertilization increased rice foliar growth which allowed for higher humidity below the canopy and created an environment favourable for the development of RFS. Additionally, irrigation has been found to be a major factor which affects the development of RFS (Brooks et al. 2010). More work is needed on the effects of nitrogen fertilization rate and planting density affecting light level on the severity of RFS.
In conclusion, the optimal ranges of temperature and wetness duration were quantitatively predicted based on a Weibull model developed with data from several experiments. After further refinement of this model or other similar models for particular host cultivar by pathogen isolate combinations, the consistency of the incidence of RFS achieved by artificial inoculation with V. virens may be more reliable which in turn will facilitate evaluation of disease resistance and functional examination of virulence genes in this pathogen in the future.
References
Brooks, S. A., Anders, M. M., & Yeater, K. M. (2009). Effect of cultural management practices on the severity of false smut and kernel smut of rice. Plant Disease, 93, 1202–1208.
Brooks, S. A., Anders, M. M., & Yeater, K. M. (2010). Effect of furrow irrigation on the severity of false smut in susceptible rice varieties. Plant Disease, 94, 570–574.
Carisse, O., Bourgeois, G., & Duthie, J. A. (2000). Influence of temperature and leaf wetness duration on infection of strawberry leaves by Mycosphaerella fragariae. Phytopathology, 10, 1120–1125.
Chen, Y., Xiao, Y., & Zhao, Y. (1995). The capacity of germination and infection of overwintered chlamydospores of Ustilaginoidea virens (Cooke) Tak. Acta Phytophylacica Sinica, 22, 102–106. In Chinese, abstract in English.
Dent, J. B., & Blackie, M. J. (1979). Systems simulation in agriculture. Model Evaluation (p. 95). London: Applied Science Publishers Ltd.
Ding, Y. (1961). Growth and development of rice. Chinese Rice Cultivation (pp. 81–83). Beijing: Chinese Agricultural Press.
Duthie, J. A. (1997). Models of the response of foliar parasites to the combined effects of temperature and duration of wetness. Phytopathology, 87, 1088–1095.
Fang, Z. (1998). Isolation and culture of fungi. Research Method of Plant Pathology (pp. 122–125). Beijing: China Agriculture Press. In Chinese.
Fujita, Y., Sonoda, R., & Yaegashi, H. (1989). Inoculation with conidiospores of false smut fungus to rice panicles at the booting stage. Annual Phytopathology Society Japan, 55, 629–634. In Japanese, abstract in English.
Gao, B., & Zhong, J. (2011). Research advances in infection process of Ustilaginoiden virens. Hunan Agricultural Sciences, 3, 93–97. In Chinese, abstract in English.
Huang, X. (1978). Development of rice panicles. Physiology of Rice Cultivation (pp. 120–128). Shanghai: Shanghai Scientific & Technical Publishers. In Chinese.
Huang, S., & Yu, L. (2002). Present situation of studies on rice false smut (Ustilaginoidea virens) in China. Acta Agriculturae Jiangxi, 2, 45–51. In Chinese, abstract in English.
Ikegami, H. (1962). Study on false smut of rice v. seeding inoculation with the chlamydospores of the rice false fungus. The phytopathological Society of Japan, 27, 16–23. In Japanese, summary in English.
Li, Y., Kang, B., Zhang, B., Lan, Y., Zeng, H., Ma, H., Xie, K., & Li, T. (1986). A preliminary study on the rice false smut. Guangdong Agricultural Science, 4, 45–47 (In Chinese).
Li, Y., Koiso, Y., Kobayashi, H., Hashimoto, Y., & Iwasaki, S. (1995). Ustiloxins, new antimitotic cyclic peptides: interaction with porcine brain tubulin. Biochemical Pharmacology, 49, 1367–1372.
Lu, F., Chen, Z., Chen, Y., Shi, A., Wang, F., & Zhang, H. (1996). Study on biological characters and some uncertain key point relation to infection cycle of rice false smut. Jiangsu Journal of Agricultural Science, 12, 35–40. In Chinese, abstract in English.
Lv, B., Luo, H., Zhang, Q., Wang, S., & Huang, J. (2008). Research advances of rice false smut. Hubei Plant Protection, S1, 47–48. In Chinese.
Ma, H., Liu, P., Liu, S., Chen, L., Jiang, M., & Zhong, W. (2001). The preliminary research on raw mycotoxin produced by rice false smut. Liaoning Agricultural Sciences, 2, 40–41.
Nakamura, K., Izumiyama, N., Ohtsubo, K., Koiso, Y., Iwasaki, S., Sonoda, R., Fujita, Y., Yaegashi, H., & Sato, Z. (1994). “Lupinosis”-like lesions in mice caused by ustiloxin, produced by Ustilaginoidea virens: a morphological study. Natural Toxins, 2, 22–28.
Savary, S., Castilla, P. N., Elazegui, F. A., McLaren, C. G., Ynalvez, M. A., & Teng, P. S. (1995). Direct and indirect effects of nitrogen supply and disease source structure on rice sheath blight spread. Phytopathology, 85, 959–965.
Tan, W. (1991). Richards function: a general model and its application in studying plant disease epidemics in time. Acta Phytopathologica Sinica, 21, 235–240. In Chinese, abstract in English.
Tan, X. (2008). Evaluation of rice varieties against the false smut and genetic diversity of Ustilaginiodea virens. [D]. Changsha: Hunan Agriculture University.
Tang, Y., Jin, J., Hu, D., Yong, M., Xu, Y., & He, L. (2013). Elucidation of the infection process of Ustilaginoidea virens (teleomorph, Villosiclava virens) in rice spikelets. Plant Pathology, 62, 1–8.
Wang, G. (1988). Studies on the factors influencing the germination of the chlamydospores of Ustilaginoidea virens. Acta Phytophylacica Sinica, 15, 241–245. In Chinese, abstract in English.
Wang, J., & Chen, Y. (2005). A survey of epidemic situation in Hunan province of year 2004. Plant Protection Guide in China, 25, 14–15. In Chinese.
Wang, Z., Lin, K., & Fan, H. (1987). The application of Weibull model to groundnut rust forecasting. Journal of South China Agricultural University, 8, 35–39. In Chinese, abstract in English.
Wang, D., Wang, S., & Fu, J. (2004). Research progress of rice false smut. Liaoning Agricultural Sciences, 1, 21–24. In Chinese, abstract in English.
Wang, S., Li, M., Dong, H., Liu, X., Bai, Y., & Yang, H. (2008). Sporulation, inoculation methods and pathogenicity of Ustilaginoidea albicans, the cause of white rice false smut in China. Journal of Phytopathology, 156, 755–757.
Wu, L., Damicone, J. P., Duthie, J. A., & Melouk, H. A. (1999). Effects of temperature and wetness duration on infection of peanut cultivars by Cercospora arachidicola. Phytopathology, 89, 653–659.
Yang, X., & Dai, X. (2009). Observation of mitosis and meiosis in rice cells by simple squash method. Agricultural Science and Technology, 10, 96–99.
Yang, H., & Zhou, C. (1979). The whole staining technology of rice anther. Hereditas (Beijing), 1, 35–36.
Zhang, J., Zhang, B., Chen, Z., Liu, Y., & Lu, F. (2003). Study on biology of conidia of Ustilaginoidea virens. Acta Phytopathologica Sinica, 33, 44–47. In Chinese, abstract in English.
Zhang, J., Zhang, B., Chen, Z., Liu, Y., & Lu, F. (2004). Research on inoculation technology of rice false smut. Acta Phytopathologica Sinica, 34, 463–467. In Chinese, abstract in English.
Zhang, S., Chen, Q., Lv, L., Yang, X., & Yu, D. (2006). Resistance evaluation of main rice cultivars in Hubei province to Ustilaginoidea virens. Anhui Agricultural Science Bulletin, 12, 76–78.
Zhou, Y. (2004). Specific and sensitive detection of the fungal pathogen Ustilaginoidea virens by nested PCR. Mycosystema, 23, 102–108.
Zhou, Y., Pan, Y., & Li, Z. (2008). Genetic diversity of rice false smut fungus, Ustilaginoidea virens and its pronounced differentiation of populations in North China. Journal of Phytopathology, 156, 559–564.
Acknowledgments
We thank Drs. Daohong Jiang and Guoqing Li for their instructive advice and comments during the course of this work. This work was supported by the Special Fund for Agro-scientific Research in the Public Interest (200903039-5), the Program for Changjiang Scholars and Innovative Research Team in University (IRT1247), the Fundamental Research Funds for the Central Universities of China (No. 2013JC006) and the Fundamental Research Funds for the Central Universities of China (No. 2013PY110).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Jia, Q., Lv, B., Guo, M. et al. Effect of rice growth stage, temperature, relative humidity and wetness duration on infection of rice panicles by Villosiclava virens . Eur J Plant Pathol 141, 15–25 (2015). https://doi.org/10.1007/s10658-014-0516-4
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10658-014-0516-4