
Abstract
Apple orchards are agro-ecosystems managed with high levels of inputs and especially pesticides. Epigeal spider communities were sampled in three seasons using pitfall traps in 19 apple orchards with four different management strategies (abandoned, under organic, Integrated Pest Management or conventional protection) and thus significantly different pesticide usage. The abundance and diversity of the spider communities was the highest in abandoned orchards. Higher diversity and evenness values were the only difference in spider communities from the organic orchards compared to the other commercial orchards. The analysis of five ecological traits (proportion of aeronauts, type of diet, overwintering stages, body size and maternal care), however, clearly showed differences in the spiders from the organic orchards. The spider species in the other commercial orchards were smaller and have higher dispersal abilities. Seven bioindicator species were identified in abandoned orchards, two species in organic ones (only Lycosidae) and one species in conventional orchards (Linyphiidae).
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Numerous studies have shown that spiders are a suitable invertebrate model group for studying ecological impacts of habitat quality and its modifications (Bonte et al. 2004; Lambeets et al. 2008). Several features explain this: they are generalist predators, many species live in the same habitat, they inhabit numerous spatial and temporal niches and are easy to collect (Wise 1995). Thus spiders are useful indicators of the ecological status of biotic communities (Marc et al. 1999).
Spiders may play an important role in the control of pest populations (Nyffeler and Sunderland 2003). However spider diversity may also be influenced by cultivation practices and especially the use of insecticides, acaricides, fungicides and herbicides. In many cases, the application of insecticides led to a decrease in both their abundance and diversity (Park et al. 2007; Santos et al. 2007; Marko et al. 2009). The abundance and species richness of spiders decreased in orchards under conventional management practices, due to the use of non-selective pesticides, compared to integrated pest management (IPM), and organic orchards, a general observation for many agro-ecosystems, and specifically in fruit orchards (Marko et al. 2009).
Commercial apple orchards are highly manipulated agro-ecosystems, in which inputs of a large amount of pesticides are used to increased productivity. However this results in a parallel increase in negative externalities (Croft 1982). In such a fruit production system, the potential for exposure of wildlife to frequent pesticide spraying is considerable (Sauphanor et al. 2012). French apple orchards approximate a global area of 54,000 ha, and are the most intensively sprayed crop (Codron et al. 2003). To date, French growers have three available strategies, for which diverging environmental impacts are expected. First, the conventional management strategy uses synthetic chemical pesticides only and remains the most frequently implemented. Second, the IPM strategy involves restricted use of chemical compounds against Cydia pomonella, the major pest in apple production, in addition to alternative practices with low environmental impact such as mating disruption (saturation of an area with synthetic female pheromone). Third, the organic management strategy excludes both synthetic fertilisers and pesticides and uses both mating disruption and a biological insecticide (the granulosis virus) against C. pomonella (Sauphanor et al. 2009).
In orchards, spiders are found both on the ground and in the crowns. Although the species in the crown are the most affected by pesticide applications (except for herbicides) (Pekar 2012), important communities of spider species can also be found on the ground or in the vegetation under the trees. In the present study, we chose to test the effects of different chemical management practices (from conventional to organic and abandoned ones) on the ground spider communities of apple orchards using a classical approach studying spider abundance and diversity, but also using a trait-based approach. The development of a trait-based ecology provides insight into assemblage-wide functional responses in environmentally variable environments (LeViol et al. 2008; Violle et al. 2007). Until now, in spiders, this approach by traits has been rarely used (Lambeets et al. 2009; Langlands et al. 2011). For environments that are strongly affected by natural or anthropogenic disturbance, species assemblages are expected to be structured by their ability (traits) to react to these disturbances (Ribera et al. 2001; Bonte et al. 2006). In apple orchards, the disturbances are a complex result of pesticides usages with direct (lethal) and indirect (depletion of preys) effects (Pekar 2012). Our aim was thus to study the epigeal spider communities in apple orchards in response to the seasonal dynamics and pest management strategies (organic, IPM and conventional). Abandoned orchards were added as a non-commercial reference. A functional trait-based approach was used to determine whether the observed changes in abundance and diversity of spiders could be explained by behavioural, ecological and phenological traits of the spider species. Finally the bioindicator status of some spider species was assessed for both factors (season and pest management practice).
Materials and methods
Sampling sites
Field sampling was carried out in three seasons (Autumn of 2009 (5th October), Spring (11th May) and Summer (26th July) of 2010) in 19 apple orchards located in a roughly 15 km diameter circular region, south of Avignon (N43.950° E4.816°, South-East of France). In the commercial orchards, pesticides were applied from mid-March to August. Thus, more cumulative effects are expected in the summer than in the spring samples. The autumn sample was taken after at least 2 months without pesticide application. Fifteen (five of each protection management type) of these orchards were commercial ones and four had been abandoned since at least 5 years. Abandoned orchards were chosen as a reference without any human perturbation (no pesticides, no traffic, no tree pruning, no irrigation, no fertilisers) resulting in a more or less closed vegetation style (brushes). The commercial orchards were chosen (i) to be representative of the three pest management strategies present in this region (organic, IPM and conventional) and (ii) with similar small-scale landscape conditions (kind of hedges and proportion of orchards in the neighbourhood). At the end of 2010, the treatment calendars of each grower were recorded. The calendars were analysed by computing global frequency treatment indices (FTI) as the mean number of treatments per hectare with commercial products, weighted by the ratio of the dose used to the recommended dose (Jørgensen 1999). This indice is thus a measure of pesticide use intensity. However, since organic farming often used different kinds of pesticides compared to IPM or conventional producers, TFI were determined for five classes of pesticides: mineral fungicides, synthetic fungicides, microbiological insecticides (granulosis virus and Bacillus thuriengiensis), synthetic insecticides and insecticides authorized under organic farming (mineral oils, Pyrethrum and spinosad). For each pesticide class, a non-parametric test (Mann–Whitney test) was used to determine differences between the three management practices.
Spider sampling and identification
Spiders were collected using pitfall traps (diameter = 11 cm, depth = 8 cm) partially filled with a solution of diluted (1:5) ethylene glycol mixed with a few ml of detergent. In each orchard at each date, six pitfall traps were placed in the centre of three different inter-rows of the orchard separated from each other by at least one row to avoid spatial autocorrelation. The rows, all covered with grass, were chosen to be the furthest possible from the borders. In the selected rows, the first and the last pitfall traps were located at least 20 m from the row extremities. Pitfall traps were always placed more than 20 m away from any hedges or orchard borders. Their contents were collected after 1 week and preserved in 70 % alcohol for ulterior identification. Adult spiders were identified to the species level (Roberts 1996; Nentwig 2003). Platnick (2012) was used as a standard for taxonomy.
Spider community parameters
Different kinds of parameters of the epigeal spider communities were computed for each combination of season and type of orchard. These parameters gave an indication of the spider abundance-activity and their structural, compositional and functional diversities. To avoid the effect of rare species in analyses of compositional and functional diversity, spider species for which the abundance was less than 0.5 % of the total spider abundance were discarded.
Abundance-activity, structural diversity and species bioindication status
The means of the abundance-activity (for adults or for adults and juveniles), the number of species (species richness SR), the Shannon–Wiener diversity index (H′) and the Pielou’s equitability index (J′) were calculated using the ‘vegan’ package in R for each orchard to describe the structure of the community. We used a linear mixed-effect (LME) regression model to determine whether management type, season or their interaction influenced these parameters. Using the nlme library for R (Pinheiro et al. 2011), we created LME regression models with an intercept-only fixed-effect term and a random effect term that included the orchard which was sampled. Post-hoc comparisons were made using the ‘multcomp’ library for R.
The IndVal value (Dufrêne and Legendre 1997) quantifies the fidelity and specificity of species to groups of sites. It was computed here for each season using the ‘labdsv’ package in R to assess the potential of each spider species as a bioindicator. Only spider species for which (i) IndVal values was above the arbitrary threshold of 25 %, (ii) the P value of the associated permutation test was below 0.05 were retained.
Dissimilarity in species composition
We computed Bray–Curtis dissimilarity indices between species pools and tested whether these values varied depending on the management system (β-diversity), or seasons (within-plot heterogeneity) and the interaction of both factors using a permutational multivariate analysis of variance (permanova) (McArdle and Anderson 2001). We made 1,000 permutations on pairwise dissimilarities and calculated pseudo-F ratio tests. For this, we used the ‘adonis’ procedure of the ‘vegan’ package included in R software (Oksanen et al. 2011). Further, NMDS ordination was used to ordinate Bray–Curtis dissimilarities among orchards. NMDS is considered one of the most robust ordination methods when dealing with zero–zero species abundance pairs (Field et al. 1982). Ordinations were plotted using the ‘metaMDS’ procedure from the ‘vegan’ package in R (Oksanen et al. 2011). Ellipses were overlaid on the graph representing the 95 %-confidence interval of one standard error from the centroid of each group.
Functional profiles of spider communities
For the functional trait-based approach, five functional traits of spiders were chosen. Body size (six classes between 0 and 20 mm) was derived from Roberts (1996) and Nentwig (2003) since only adults were considered in this study. Maternal care (binary attributes) and prey types (three attributes: walking, flying or jumping) were derived from the review on hunting and reproductive strategies in Foelix (2011) and Nentwig (2013). Ballooning, which is the propensity to aerial dispersion of juveniles and adults was taken from the data of Greenstone et al. (1987) and the review of Bell et al. (2005). The status for survival during winter (overwintering) was detailed by LePeru (2006) for our species. All ecological and morphological traits were stored in a database called “Biological & Ecological functional Traits of Soil Invertebrates” (Hedde et al. 2012a). To take into account all the information gathered on each species, we did not consider traits as exclusive values instead we used the fuzzy coding method (Chevenet et al. 1994). The percentage of affinity of each species to a class of a trait was thus coded by a procedure adapted to terrestrial invertebrates (Hedde et al. 2012b, 2013). This means that a species is not “forced” to feed on only one type of traits (walking or flying or jumping) but that depending on the information found in the scientific literature, a species can have, for example, a diet of 50, 30 and 20 % of flying, walking and jumping prey respectively. Community-weighted means trait value (CWM; Lavorel et al. 2008) were computed using the ‘FD package in the R environment (Laliberté and Shipley 2011). CWM for one trait is simply the mean affinity of the spider community for this trait taking into account (i) the trait value of each species composing this community and (ii) the species abundance. CMW were then used to analyse functional profiles (i.e. distribution of affinities between the classes of a trait) in response to orchard management type or season. We used LME, as described previously, to identify the effect of the studied factors on CWM. Post-hoc comparisons were made using the ‘multcomp’ library for R.
Results
Frequency treatment index
No significant difference between the three management strategies in commercial apple orchards was observed regarding the global TFI. This suggests that the perturbation due to tractor (number of engine passages) is similar between the three commercial management types. However, the kind of pesticides applied was significantly different (Fig. 1). Orchards under organic management were characterised by a higher usage of microbial insecticides (with granulosis virus representing about 90 % of these applications) and a concurrent lower usage of other insecticides with broad-spectrum of toxicity. We observed a limited but significant difference (P = 0.036) between conventional and IPM management strategies: mating disruption replaced three synthetic insecticide applications.

Annual mean values (and standard deviations) for the treatment frequency indices (TFI) of each type of orchard management strategy, for different categories of pesticides used and the global value
Among commercial orchards, organic and non-organic ones (IPM and conventional) differed as well by other agricultural practices than protection management. Indeed organic orchards did not use herbicides under the trees or mineral fertilisers since only soil tillage and organic fertilisers are allowed. However, herbicides are only applied on the rows (i.e. on roughly 20 % of the orchard area) and the traps were located in the centre of the inter-rows. Thus even if these practices may have an effect on epigeal spiders (Minarro et al. 2009), we assumed than the main effects were indeed due to pesticide applications.
Response of community parameters
A total of 1,193 spiders were caught in pitfall traps at the three seasons. Among them, only 597 were included in the study as they were adults (and thus identified to the species level). This is the drawback of using a trait-based approach since most of the traits are clearly defined for species only. The most common spider families were Lycosidae (29.3 or 29.0 % when juveniles were included), Zodariidae (26.6 or 22.6 % when juveniles were included) but with only two species, Linyphiidae (18.9 or 20.3 % when juveniles were included), Hahniidae (11.6 or 6.4 % when juveniles were included) and Gnaphosidae (4.6 or 11.4 % when juveniles were included) (Table 1).
Impacts on the abundance-activity and structural diversity
All parameters used to characterize the spider communities varied significantly depending on orchard type and season (Table 2). Indices recorded in abandoned orchards were always significantly higher than those recorded in IPM or conventional orchards. For example, the abundance (of adults or of adults and juveniles), SR, H′ and J′ were 4-, 2-, 3- and 2-fold higher, respectively, in abandoned compared to conventional orchards. The abundance-activities and SR of spider communities in organic orchards were the same as those for conventional or IPM orchards. In contrast, their H′ and J′ indices were similar to those calculated for abandoned orchards. Regarding season, all community parameters were significantly higher in summer (Table 2). A significant interaction between season and orchard type was only significant for H′ and abundance.
Impacts on the composition of spider communities
The results of the permanova on pairwise Bray–Curtis dissimilarity matrices showed highly significant effects (P < 0.001) for both factors and their interaction. When ordinated by NMDS, management types were clearly separated (Fig. 2). Three groups could be identified from the overlap of CI-ellipses: (i) conventional and IPM, (ii) organic and (iii) abandoned orchards. In this kind of representation, the differences between seasons appeared to be weaker and probably in interaction with management type as highlighted by the permanova results.

NMDS plot of spider assemblage composition showing the 95 % confidence interval ellipse for both factors: orchard management type (plain lines ‘Ab’ for abandoned, ‘PFI’ for IPM, ‘conv’ for conventional and ‘Bio’ for organic orchards) and season (dashed lines)
Impacts on the functional profiles of spider communities
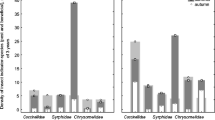
We found that management type had significant effects on all functional traits. Season had an effect on diet and the overwintering stage. No significant interaction was found between the two factors on any traits (Table 3). The percentage of spiders with the ability to disperse through aeronauts (at both the juvenile and adult stages) was much lower in organic and abandoned orchards (5–7 %) than in IPM and conventional orchards (43–63 %). The percentage of individuals feeding on walking preys decreased from abandoned (59 %) to organic (39 %) and to IPM and conventional (17–27 %) orchards. Subsequently, the percentage of individuals feeding on either jumping or flying prey displayed an inverse trend. Considering seasonal effects, the percentage of individuals feeding on jumping prey was rather stable (30–40 %). In contrast, we observed an increase in the percentage of predators of walking preys from fall to spring and to summer (19, 33 and 46 %, respectively), to the detriment of the percentage of flying prey predators (41, 34 and 23 %, respectively). The percentage of spiders which showed maternal care significantly decreased from organic orchards (55 %), to IPM orchards (26 %) and to abandoned or conventional orchards (2–7 %). The communities mostly consisted of spiders which overwinter as adults (39–82 %) or as juveniles (13–46 %). The highest percentage of spiders overwintering in egg sacs, as juveniles and adults was found in abandoned (22 %), organic (46 %) and conventional orchards (82 %), respectively (Table 3). The cumulated distribution of body size was not significantly different between management types in autumn. In spring (Fig. 3) and in summer (data not shown), conventional and IPM orchards hosted communities with a significantly higher proportion of smaller individuals than organic or abandoned orchards.

Cumulative distribution (mean curve and 95 % confidence intervals) of the percentage of spider individuals with increasing body size classes in spring (for the sake of readability, only commercial orchards are represented; curves for abandoned orchards are not significantly different from those for organic orchards)
Bioindicator species
The IndVal method classified 0 (IPM), 1 (conventional), between 1 and 2 (organic) and between 3 and 4 (abandoned) species as potential bioindicators for the different seasons (Table 4). In the abandoned orchards, these species belonged to five families: two species of Zodariidae and Hahniidae and one species of Gnaphosidae, Lycosidae and Theriididae and Thomisidae. Z. italicum (Zodaridae) appeared to be indicator species regardless of season. In organic orchards, two species were identified as possible bioindicators (Lycosidae: P. proxima and P. hortensis) in spring and/or summer. No species was found to be a relevant bioindicator in IPM orchards in any season. Conventional orchards had one bioindicator species (Linyphiidae: O. apicatus) in autumn and summer. It is interesting to note that most of the species have at least a Paleartic distribution (especially those the bioindicator species in commercial orchards). Three species are limited to Europe (H. petrobia, O. sanctuary and Z. italicum) and only two to the Mediterranean zone (H. signifer and A. albomaculata).
Discussion
Effects of management type and season on the structure of spider communities
Not surprisingly, the spider epigeal communities sampled in the abandoned orchards were characterised by the highest abundance and species richness values and by higher H′ and J indices (but similar to those from organic orchards). Indeed, abandoned orchards were chosen as an undisturbed reference where no pesticides had been applied but also lacking any human intervention (fertilisation, pruning, grass mowing). These findings are related to the increased number and diversity of micro-habitats in these orchards (Langellotto and Denno 2004).
Orchard management types had a significant effect on the taxonomical composition but not on total abundance and species richness. Generally speaking, organic farming was shown to enhance the abundance of natural enemies abundance and species richness in various agrosystems (Bengtsson et al. 2005; Hole et al. 2005). For epigeal or ground spiders, several studies found that spider communities were more abundant (Pfiffner and Luka 2003; Cardenas et al. 2006; Prieto-Benitez and Mendez 2011) or more species-rich in organic systems (Pekar and Koucourek 2004). This was not the case in our study where epigeal spiders abundance and species richness were similar in organic, IPM and conventional orchards. This result was not completely unexpected because the total TFI values were similar between these three management strategies. In addition some insecticides used in organic orchards are broad-spectrum insecticides such as spinosad or pyretre (Bahlai et al. 2010). The two microbial insecticides, granulosis virus and Bacillus thuringiensis, mainly used under organic management strategies, are expected to have a low direct toxicity towards spider (Bajwa and Aliniazee 2001). Nevertheless the organic orchards did have significantly higher H′ and J indices. This is in agreement with the results of the meta-analysis carried out by Crowder et al. (2010) which showed that in general organic farming increases evenness. None of the characteristics of the spider communities separated IPM from conventional orchards. We presume that the difference in the number of insecticide applications was not great enough to have visible effects.
Ordination of the species assemblage based on Bray–Curtis dissimilarities showed that spider communities were different between (i) abandoned, (ii) organic and (iii) IPM and conventional orchards. Differences between seasons were less marked. Communities sampled in summer, where the cumulative effects of the pesticide applications are the highest, appeared to be different from autumn and spring communities but despite this were still characterised by the higher abundance, species richness, H′ and J′ indices. As previously reported by Marko et al. (2009), differences in spider communities between IPM and conventional orchards were much smaller in autumn in apple orchards due to recolonisation processes by spiders from the hedges or plots in the neighbourhood.
The differences in spider communities according to orchard management strategy resulted in the presence of spider species with a significant bioindicator value. These species were more numerous in abandoned orchards (eight species belonging to six different families) and showed seasonal variations with the noticeable exception of Z. italicum. This illustrates (i) the diversity of niches that can be encountered in non-mowed orchards and (ii) the effect of the absence of human perturbations on spider communities. Bioindicator species for organic orchards were found only in spring and summer with two Lycosidae species of the Pardosa genus. No indicator species was found for the IPM strategy and one Linyphiidae species was found for the conventional orchards (O. apicatus). These results are in agreement with previous studies by Pfiffner and Luka (2003) or Glück and Ingrisch (1990) which found that some Pardosa species had higher abundances in organic compared to conventional plots. The latter authors also found that conventional arable plots were dominated by two Linyphiids (Erigone atra and O. apicatus). Additionally, Feber et al. (1998) previously observed that O. apicatus densities were higher in conventional wheat compared to organic plots. These results are likely to be linked with the high recolonisation ability of O. apicatus from the borders of the plots after a treatment (Lemke and Poehling 2002).
Relevance of trait-based approaches
Our results highlight the usefulness of trait-based approaches for depicting the effects of orchard management and season on epigeal spiders. The orchard management strategy was found to significantly influence the five selected ecological traits. Nevertheless, season also had an influence on some of the ecological traits examined.
Trait-based approaches were previously applied to spider communities to study landscape fragmentation in apple orchards (Herrmann et al. 2010), and heathland restoration (Cristofoli et al. 2010). To our knowledge, this approach has not yet been used to study the effects of management strategies (including abandoned orchards) or seasons (i.e. different intensity of pesticide application) on spider communities. In our study, five traits were chosen to encompass important and different ecological functions: resource conservation (body size), recolonisation ability (aeronauts), resource acquisition (feeding strategy), survival mode in negative climatic conditions (overwintering type) and fitness (maternal care).
Overall, the four types of orchard management represented an intensification gradient characterised by two different kinds of perturbations. The abandoned orchards are at one end of the gradient, creating a stable environment without any human interference. Next the organic orchards differed from abandoned orchards mainly as a result of mechanical perturbations (grass cutting, pruning) and to a lesser extent, by the use of a few pesticides. At the other end of this gradient, the IPM and then conventional orchards differed mainly from organic ones due to the intensive use of synthetic pesticides. These pesticides have two complementary effects through direct toxicity and indirectly through resource depletion (Marko et al. 2009; Pekar 2012). For example, insecticides may reduce overall insect abundance and thus limit prey availability. This can in turn influence the body size and fecundity of spiders or induce their emigration and postpone recovery (Wisniewska and Prokopy 1997; Pekar 2012).
Abandoned orchards with higher, more stable and predictable resources, hosted spiders species with larger body sizes, lower dispersal abilities but no maternal care such as Gnaphosidae (as exemplified by Haplodrassus signifer being an indicator species). This relationship between large body size and the greater availability of niches was illustrated by Wilson (1975). Organic orchards also hosted species with large body sizes, low dispersal abilities and overwintering mainly as juveniles but were also characterised by a very proportion of individuals exhibiting maternal care. As a consequence, some Lycosidae species (Pardosa hortensis and P. proxima) were found to have significant bioindication values for this orchard management strategy. Conventional orchards, subjected to the higher insecticide pressure, hosted species with smaller body sizes and thus high dispersal ability that are able to recolonize orchards from its borders (Bonte et al. 2004; Schmidt et al. 2008). Moreover the spider community in conventional orchards is as well characterised by spider overwintering mainly as adults. The linyphiid spider O. apicatus is significantly associated with this kind of management strategy. These results are in agreement with previous studies indicating that Linyphiidae are often associated with disturbed environments (Topping and Sunderland 1994; Halley et al. 1996).
References
Bahlai CA, Xue Y, Mc Creary CM, Schaafsma AW, Hallett RH (2010) Choosing organic pesticides over synthetic pesticides may not mitigate environmental risks in soybeans. PLoS One. doi:10.1371/journal.pone.0011250
Bajwa WI, Aliniazee MT (2001) Spider fauna in apple ecosystem of western Oregon and its field susceptibility to chemical and microbial insecticides. J Econ Entomol 94:68–75
Bell JR, Bohan DA, Shaw EM, Weyman GS (2005) Ballooning dispersal using silk: world fauna, phylogenies, genetics and models. Bull Entomol Res 95:69–114
Bengtsson J, Ahnstrom J, Weibull AC (2005) The effects of organic agriculture on biodiversity and abundance: a meta-analysis. J Appl Ecol 42:261–269
Bonte D, Baert L, Lens L, Maelfait JP (2004) Effects of aerial dispersal, habitat specialisation and landscape structure on spider distribution across fragmented grey dunes. Ecography 27:343–349
Bonte D, Lens L, Maelfait JP (2006) Sand dynamics in coastal dune landscapes constrain diversity and life-history characteristics of spiders. J Appl Ecol 43:735–747
Cardenas M, Ruano F, Garcia P, Pascual F, Campos M (2006) Impact of agricultural management on spider populations in the canopy of olive tress. Biol Control 38:188–195
Chevenet F, Doledec S, Chessel D (1994) A fuzzy coding approach for the analysis of long-term ecological data. Freshw Biol 31:295–309
Codron JM, Habib R, Jacquet F, Sauphanor B (2003) Bilan et perspectives environnementales de la filière arboriculture fruitière. In: Dron D (ed) Agriculture, territoire, environnement dans les politiques européennes. Dossiers de l’Environnement de l’INRA 23, pp 31–67
Cristofoli S, Mahy G, Kekenbosh R, Lambeets K (2010) Spider communities as evaluation tools for wet heatland restoration. Ecol Indic 10:773–780
Croft BA (1982) Arthropod resistance to insecticides: a key to pest control failures and successes in North American apple orchards. Entomol Exp Appl 31:88–110
Crowder DW, Northfield TD, Strand MR, Snyder WE (2010) Organic agriculture promotes evenness and natural pest control. Nature 466:109–123
Dufrêne M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol Monogr 67:345–366
Feber RE, Bell J, Johnston PJ, Firbank LG, Macdonald DW (1998) The effects of organic farming on surface-active spider (Aranea) assemblages in wheat in southern England, UK. J Arachnol 26:190–202
Field JG, Clarke KR, Warwick RM (1982) A practical strategy for analysing multispecies distribution patterns. Mar Ecol Prog Ser 8:37–52
Foelix RF (2011) Biology of spiders. Oxford University Press, New York
Glück E, Ingrisch S (1990) The effect of biodynamics and conventional agriculture management on Erigoninae and Lycosidae spiders. J Appl Entomol 110:136–148
Greenstone MH, Morgan CE, Hultsh A-L (1987) Ballooning spiders in Missouri, USA, and New South Wales, Australia: family and mass distributions. J Arachnol 15:163–170
Halley JM, Thomas CFG, Jepson PC (1996) A model for the spatial dynamics of linyphiid spiders in farmland. J Appl Ecol 33:471–492
Hedde M, Pey B, Auclerc A, Capowiez Y, Cluzeau D, Cortet J et al (2012a) BETSI, a complete framework for studying soil invertebrate functional traits. XVI ICSZ—International Colloquium on Soil Zoology, Coimbra
Hedde M, van Oort F, Lamy I (2012b) Functional traits of soil invertebrates as indicators for exposure to soil disturbance. Environ Pollut 164:59–65
Hedde M, van Oort F, Renouf E, Thénard J, Lamy I (2013) Dynamics of soil fauna after plantation of perennial energy crops on polluted soils. Appl Soil Ecol 66:29–39
Herrmann JD, Bailey D, Hofer G, Herzog F, Schmidt-Entling MH (2010) Spiders associated with meadow and tree canopies of orchards respond differently to habitat fragmentation. Landsc Ecol 25:1375–1384
Hole DG, Perkins AJ, Wilson JD, Evans AD (2005) Does organic farming benefit biodiversity? Biol Conserv 122:113–130
Jørgensen LN (1999) Denmark’s action plans for pesticides: status and role of research. Nord Jordbrugsforsk 81:201–202
Laliberté E, Shipley B (2011) FD: measuring functional diversity from multiple traits, and other tools for functional ecology. R package version 1.0-11
Lambeets K, Hendrickx K, Vanacker S, Van Looy K, Maelfait JP, Bonte D (2008) Assemblage structure and conservation value of spiders and carabid beetles from restored lowland river banks. Biodivers Conserv 17:3133–3148
Lambeets K, Vandegehuchte ML, Maelfait JP, Bonte D (2009) Integrating environmental conditions and functional life-history traits for riparian arthropods conservation planning. Biol Conserv 142:625–637
Langellotto GA, Denno RF (2004) Responses of invertebrate natural enemies to complex-structured habitats: a meta-analytical synthesis. Oecologia 139:1–10
Langlands PR, Brennan KEC, Frameneau VW, Main BY (2011) Predicting the post-fire responses of animal assemblages: testing a trait-based approach using spider. J Anim Ecol 80:558–568
Lavorel S, Grigulis K, McIntyre S, Williams NSG, Garden D, Dorrough J, Berman S, Quetier F, Thebault A, Bonis A (2008) Assessing functional diversity in the field—methodology matters! Funct Ecol 22:134–147
Lemke A, Poehling H-M (2002) Sown weed strips in cereal fields: overwintering site and “source” habitat for Oedothorax apicatus (Blackwall) and Erigone atra (Blackwall) (Araneae: Erigonidae). Agric Ecosyst Environ 90:67–80
LePeru B (2006) Catalogue et repartition des Araignées de France. Rev Arachnol 16:1–468
LeViol I, Julliard R, Kerbiriou C, Redon L, Carnino N, Machon N, Porcher E (2008) Plant and spider communities benefit differently from the presence of planted hedgerows in highway verges. Biol Conserv 141:1581–1590
Marc P, Canard A, Ysnel F (1999) Spiders (Araneae) useful for pest limitation and bioindication. Agric Ecosyst Environ 74:229–273
Marko V, Keresztes B, Fountain MT, Cross JV (2009) Prey availability, pesticides and the abundance of orchard spider communities. Biol Control 48:115–124
McArdle BH, Anderson MJ (2001) Fitting multivariate models to community data: a comment on distance-based redundancy analysis. Ecol 82:290–297
Minarro M, Espadaler X, Melero VX, Suarez-Alvarez V (2009) Organic versus conventional management in an apple orchard: effects of fertilization and tree-row management on ground-dwelling predaceous arthropods. Agric For Entomol 11:133–142
Nentwig W (2003) Management of biodiversity in agroecosystems. Basic Appl Ecol 4:105–106
Nentwig W (2013) Ecophysiology of spiders. Springer-Verlag, Berlin
Nyffeler M, Sunderland KD (2003) Composition, abundance and pest control potential of spider communities in agroecosystems: a comparison of european and US studies. Agric Ecosyst Environ 95:579–612
Oksanen J, Blanchet FG, Kindt R, Legendre P, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Wagner H (2011) Vegan: community ecology package. R package version 1.17-7
Park YC, Joo JS, Kim JP (2007) Diversity of spider communities in a pesticide-treated pine (Pinus densiflora) forest. J Ecol Field Biol 30:179–186
Pekar S (2012) Spiders in the pesticide world: an ecotoxicological review. Pest Manag Sci 68:1438–1446
Pekar S, Koucourek F (2004) Spider (Araneae) in the biological and integrated pest management of apple in Czech Republic. J Appl Entomol 128:561–566
Pfiffner L, Luka H (2003) Effects of low-input farming systems on carabids and epigeal spiders—a paired farm approach. Basic Appl Ecol 4:117–127
Pinheiro J, Bates D, DebRoy S, Sarkar D and the R Development Core Team (2011) Nlme: linear and nonlinear mixed effects models. R package version 3.1-113
Platnick NI (2012) The world spider catalog, version 13.0. American Museum of Natural History. doi:10.5531/db.iz.0001
Prieto-Benitez S, Mendez M (2011) Effect of land management on the abundance and richness of spiders (Aranea): a meta-analysis. Biol Conserv 144:683–691
Ribera I, Doledec S, Downie IS, Foster GN (2001) Effect of land disturbance and stress on species traits of ground beetle assemblages. Ecology 82:1112–1129
Roberts MJ (1996) Spiders of Britain and Northern Europe. Harper Collins, London
Santos SAP, Pereira JA, Torres LM, Nogueira AJA (2007) Evaluation of the effects on canopy arthropods of two agricultural management systems to control pests in olive groves from the north-east of Portugal. Chemosphere 67:131–139
Sauphanor B, Dirwimmer C, Boutin S, Chaussabel AL, Dupont N, Fauriel J, Gallia V, Lambert N, Navarro E, Parisi L, Plenet D, Ricaud V, Sagnes JL, Sauvaitre D, Simon S, Speich P, Zavagli F (2009) Analyse comparative de différents systèmes en arboriculture fruitière. In: INRA (Ed), Ecophyto R&D: vers des systèmes de culture économes en produits phytosanitaires. Rapport d’expertise, Tome IV, INRA, Paris
Sauphanor B, Severac G, Maugin S, Toubon J-F, Capowiez Y (2012) Exclusion netting may alter reproduction of the codling moth (Cydia pomonella) and prevent associated fruit damage to apple orchards. Entomol Exp Appl 145:134–142
Schmidt MH, Thies C, Nentwig W, Tscharntke T (2008) Contrasting responses of arable spiders to the landscape matrix at different spatial scales. J Biogeogr 35:155–166
Topping CJ, Sunderland KD (1994) A spatial population-dynamics model for Leptyphantes-tenuis (Araneae, Linyphiidae) with some simulations of the spatial and temporal effects of farming opérations and land-use. Agric Ecosyst Environ 48:203–217
Violle C, Navas ML, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E (2007) Let the concept of trait be functional. Oikos 116:882–892
Wilson EO (1975) The adequacy of body size as a niche différence. Am Nat 109:769–784
Wise D (1995) Spiders in ecological webs. Cambridge University Press, Cambridge
Wisniewska J, Prokopy RJ (1997) Pesticide effect on faunal composition, abundance, and body length of spiders (Araneae) in apple orchards. Environ Entomol 26:763–776
Acknowledgments
The authors warmly thanked the producers and owners for allowing them to sample spiders in their orchards. This study is dedicated to the memory of P. R., one of the producers, who committed suicide in 2012. The authors also wish to thank our colleagues participating in the BETSI project (funded by the French Foundation for Biodiversity) for valuable discussions on trait-based approaches.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Mazzia, C., Pasquet, A., Caro, G. et al. The impact of management strategies in apple orchards on the structural and functional diversity of epigeal spiders. Ecotoxicology 24, 616–625 (2015). https://doi.org/10.1007/s10646-014-1409-1
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-014-1409-1