
Abstract
For the purpose of biomonitoring, species that combine ecological and commercial importance may provide a link between ecological and human health risk. The common cuttlefish, Sepia officinalis, holds both characteristics in south-western Europe, albeit remaining unsurveyed in ecotoxicological studies. Cuttlefish collected from an impacted estuary in SW Portugal and a reference location off the coast were analysed for a battery of biomarker responses in the digestive gland and gills. The contrast to reference animals revealed that biomarker responses, especially those related to oxidative stress, were consistent with sediment contamination by PAHs, even in a situation that combines complex toxicant mixtures, moderate levels of contamination and high ecotoxicological diversity. However, environmental parameters related to the differences between shore and estuarine habitats should not be overruled. Also, digestive gland metallothionein retained significant specificity to metals even though previous studies in the area with clams and fish failed to trigger a conclusive response. The highest net differences in biomarker responses were detected in the gills, likely indicating higher sensitivity to environmental stressors. Still, the digestive gland responses were overall the most consistent with sediment contamination and effectively differentiated between estuarine industrial- and rural-impacted sites. The results indicate that S. officinalis may be a candidate to meet the European Union’s requirements for efficient biomonitoring programmes, with the additional importance of being cosmopolitan, abundant, commercially valuable and combining the molluscan biology that has been granting bivalves their high value for biomonitoring with foraging behaviour, thus better able to reflect anthropogenic stressors impacting a wider area than sedentary organisms. Nevertheless, further investigations in unpolluted sites are needed to better evaluate the background levels of biomarker responses in the species.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Coastal environments, especially confined waterbodies such as estuaries, are impacted by various anthropogenic pressures. The release of xenobiotics, inherent to many human activities, is one of the greatest concerns. Ecological Risk Assessment (ERA) is one of the most acknowledged approaches to address the problem of coastal pollution. The process of ERA may comprise several steps, e.g. from environmental contaminant analyses to the determination of its potential effects to organisms, biomonitoring being one of the most important stages (see Chapman 2007 for a review). Recent European legislation regarding the assessment and safeguard of environmental quality in coastal environments, namely through the recent Marine Strategy Framework Directive (MSFD, Directive 2008/56/EC), implicitly or explicitly highlights the need to survey organisms that are both ecologically and economically important, therefore bridging environmental and human risk (refer to Lyons et al. 2010). However, the MSFD does not explicitly state which environmental or biological parameters are required, leaving to regional conventions (such as MEDPOL, HELCOM or OSPAR) the role to develop suitable indicators, including the choice of adequate target species, thus acknowledging the ecological diversity among Europe’s coastal areas.
It is nowadays recognized, in face of the many constraints in the interpretation of biomarker responses when contaminant interactions and other confounding factors are at stake, that integrative multi-biomarker approaches allow a more efficient evaluation of the effects of pollutants on individuals (see, for instance, Picado et al. 2007). However, the choice of adequate bioindicator species should not be astray from its relevance to the ecosystem (Martín-Díaz et al. 2008). Benthic organisms are of great importance mainly due to their interaction with the most important reservoir of xenobiotics, sediments, especially in transitional ecosystems like estuaries. Molluscs have been widely surveyed in biomonitoring programs, owing to their ecological and economical importance, albeit the vast majority of the studies are focused on clams, cockles and mussels, i.e. sedentary organisms (sessile or burrowing), thus potentially reflecting the conditions of their immediate surroundings whereas foraging animals are potential indicators for wider areas. Among the latter, benthic fish are often considered prime targets for ecotoxicological surveys, whereas cephalopod molluscs remain little studied, even though they combine foraging ability with the basic molluscan physiology. In addition, cephalopods possess high commercial value (see Guerra et al. 2010 for a recent review), hence the plausible link between ecological and human risk, if consumption rates and toxicant bioaccumulation are taken into account. Still, no ecotoxicological research has been found to date focusing on Sepia officinalis (L., 1758) as sentinel/biomonitoring organism, although Bustamante et al. (2006), in an entirely laboratorial study (with hatchery-brooded animals) disclosed the animals’ ability to significantly bioaccumulate metals. In fact, the very little toxicological research performed on cephalopods, namely on feral Octopus, revealed that cephalopods are highly sensitive to environmental toxicants but biomarker and general toxicological research in these animals is still incipient (refer to Raimundo et al. 2010 and references therein). However, when compared to the cuttlefish, octopuses are not as tolerant to environmental variation (especially salinity) and therefore are unlikely bioindicators for transition waterbodies.
Sepia officinalis is cosmopolitan, found throughout the Mediterranean basin and the eastern Atlantic Ocean, from Southern Norway and Northern England to the north-western coast of Africa and it is known to tolerate a high-range of salinities, inhabiting both marine and brackish water ecosystems (see Guerra 2006). Its wide distribution and resilience to environmental variables surpasses those of mussels, clams and flatfish (the most common subjects in marine biomonitoring programmes), potentially rendering the cuttlefish as a more efficient bioindicator for interregional comparisons. In Portugal, cuttlefish can be found throughout the coast, including transitional water bodies, which contributes to its high importance for traditional fisheries (Guerra 2006; Neves et al. 2009), particularly in the Sado estuary, where it stands as the most prized species for human consumption.
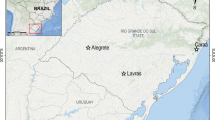
The Sado estuary (≈180 km2) is one of the largest coastal basins in Portugal (Fig. 1), with well mixed flows and high dilution potential. It is impacted by multiple anthropogenic stressors, yet most of the estuary is classified as Natural Reserve, which generates conflicts between environmental quality and the safeguard of human socio-economical activities (e.g. Costa et al. 2008a, 2011, 2012; Caeiro et al. 2009). The main pressures are located in the northern part of the estuary, due to the presence of a large heavy-industry park (including shipyards, paper mills, a thermoelectrical plant, chemical industries and mineral ore shipment facilities); the sizable metropolitan area of Setúbal, heavy-duty maritime transport, plus aquaculture and fisheries (e.g. Caeiro et al. 2005, 2009; Costa et al. 2012). Closer to the river mouth, in the southern region of the estuary, the fishing pressure is felt mostly during summer and extensive agriculture grounds (mostly rice fields) likely carry pesticides and fertilizers to the estuarine basin (Costa et al. 2009, 2011). The need to implement effective ERA strategies in the estuary led to recent biomarker-based studies in some of the most relevant commercial species to estuary’s fisheries, namely flatfish (Costa et al. 2008b, 2009, 2011) and clams (Carreira et al. 2013; Costa et al. 2013), however often producing results that are either inconclusive or contradictory to acknowledged biomarker theory due to constraints that may apply to many confined marine ecosystems: diffuse sources of pollution, moderate levels of contamination and the presence of complex mixtures of pollutants (Costa et al. 2012).

Map of the study area evidencing the fishing areas for S. officinalis (Sado 1, Sado 2 and Reference) and sediment collection sites (S1–S5 from Sado, plus M, from the reference area)
The main objectives of the present work may be summarized as: (1) to determine the potential of a novel bioindicator species, the coleoid cephalopod S. officinalis; through a biomarker approach; (2) to compare biomarker responses between two target organs, gills and digestive gland, (3) to integrate sediment contamination data with biomarker responses and (4) to obtain an accountable measure of contamination for the Sado estuary from an effective application of the species as bioindicator.
Materials and methods
Sampling
Sediment and organism sampling locations are indicated in Fig. 1. Sediment sampling and characterization are described in detail in the preceding work by Carreira et al. (2013), which is framed in the same research project as the present work. The choice of sediment collection sites within the Sado estuary (S1–S5) related to the need to obtain a general contamination overview of the two main commercial fishing areas hereby designated Sado 1 (north) and Sado 2 (south), representing colder and warmer months fishing grounds, respectively. The sediment collection sites S1, S2 and S3 are located within the northern fishing area (Sado 1), whereas the sites S4 and S5 are representative of the southern fishing area (Sado 2), located near the mouth of the river. Due to the impossibility of collecting sufficient animals from the only acknowledged clean estuary within the same biogeographical area, the small (≈5 km2) Mira Estuary (see Vasconcelos et al. 2007), the reference area for cuttlefish sampling consisted of a commercial fishing site off the coast, facing this estuary. To provide a measure of contamination of this area, a sediment collection site (M) was selected for analysis (termed M2 in Carreira et al. 2013). All sediments were collected using a grab and during the same seasons in which organisms were captured, i.e. M, S1, S2 and S3 during the fall/winter and S4 and S5 in the spring.
Approximately 20 juvenile (determined to be sexually immature upon dissection) S. officinalis (130 ± 8 mm mantle length; 300 ± 50 g total wet weight) per site were collected from the three fishing areas, Sado 1, Sado 2 and Reference. Animals, alive upon collection, were immediately transported to the laboratory on ice, in cold containers, processed for standard measurements, namely mantle length (Lm) and total wet weight (wwt), and readily dissected for the collection of digestive gland and gill samples, which were frozen at −80 °C for subsequent analyses.
Sediment characterization
Sediment contamination and physico-chemical characterization was retrieved from Carreira et al. (2013), and apply to the same ERA research project of the present work. The details on analyses are described in detail therewith. In brief: sediment granulometric fractions were obtained by hydraulic sieving and total organic matter (TOM) was determined by carbon loss-at ignition at 500 ± 25 °C during 4 h, being both results expressed as percentage per total sediment dry weight (dw). Sediment redox potential (Eh) was measured using an Orion model 20A meter with a H3131 Ag/AgCl reference electrode. Sediment metals (Cr, Ni, Cu, Zn, Cd and Pb) and metalloids (As and Se) were determined as follows: dry sediment samples were mineralized in closed Teflon vessels, followed by elution with MilliQ-grade water (18.2 mΩ cm) and quantification by inductively coupled plasma mass spectrometry using a Thermo Elemental X-Series equipment. Polycyclic aromatic hydrocarbon (PAH) concentrations were determined by a Finnigan GCQ gas chromatography–mass spectrometry (GC–MS) system and concentrations were measured by the internal standard peak area method and with a calibration curve for each compound. Organochlorines (PCBs plus the pesticides DDT and HCB) were determined by Soxhlet-extraction of dried sediments with n-hexane and quantified by GC-ECD (gas chromatography with an electron capture detector). As in Carreira et al. (op. cit.), in absence of specific Sediment Quality Guidelines (SQGs) for Portugal, the sediments’ potential to cause adverse effects to organisms was estimated by comparison of the obtained sediment contaminant concentrations with the SQGs developed for coastal waters by Macdonald et al. (1996), namely the threshold effects level (TEL) and the probable effects level (PEL) guidelines.
Biomarker analysis
In order to compare the two target organs, all biomarkers were analysed separately in the gills and digestive gland of all individuals. Lipid peroxides were determined following the thiobarbituric acid reactive substances protocol first developed by Uchiyama and Mihara (1978) and adapted by Costa et al. (2011). In brief: approximately 100 mg of each organ was homogenized in 250 μL of phosphate-buffered saline [PBS (pH 7.4, with 0.7 % NaCl)], followed by centrifuging for 5 min at 7,000×g. A 50 μL aliquot of the supernatant was taken and incubated with 100 μL of 10 % m/v trichloroacetic acid for 15 min, at 4 °C, to precipitate protein. After a 2,200×g, 15 min, centrifuging, 100 μL of 0.1 % m/v thiobarbituric acid was added to 100 μL of the clear supernatant and heat-treated (at ≈100 °C) for 15 min. The resulting red pigment was extracted with a mixture of pyridine:butanol (1:15) and the absorbance measured at 530 nm. To each well of the 96-well plates were added 150 μL of the reaction containing samples, blanks and standards. Determination was done through an eight-point calibration curve using malondialdehyde bis(dimethylacetal), from Merck, as standard.
Total glutathione (GSHt) was determined from approximately 100 mg of digestive gland and gill tissue through the enzymatic recycling method, using a commercial kit (Sigma-Aldrich), following manufacturer instructions, by measuring the increase in the absorbance of the reactions at 412 nm during 5 min at 1 min intervals. The GSH/GSSG (reduced/oxidised glutathione) ratio was estimated following derivatization of GSHt subsamples with 2-vinylpyridine (Sigma-Aldrich), in order to obtain the GSSG concentration. The ratio was determined as GSH/(GSSG/2).
To estimate glutathione S-transferase (GST) activity, a homogenate was made from approximately 100 mg of digestive gland and gill tissue in 300 μL of PBS, followed by centrifugation (5 min, 7,000×g). Activity was determined in the diluted supernatant (1:10), by measuring the increase in absorbance at 340 nm during 5 min, using chloro-2,4-dinitrobenzene (CDNB) as substrate. The procedure was performed using a commercial kit (Sigma-Aldrich), according to manufacturer instructions. An aliquot of the same homogenate was used to quantify metallothionein-like protein (MT) using the protocol described by Costa et al. (2008a), following from Paleček and Pechan (1971), with modifications. In brief: after centrifuging the homogenate for 10 min at 12,000×g, the diluted (1:10) supernatant (extracted cytosol) was heated (≈80 °C, 10 min) and centrifuged (10 min, 12 000×g), to precipitate non heat-stable proteins. In absence of an available commercial cuttlefish MT, MT-1 from rabbit liver (Alexis Biochemicals) was used to obtain a five-point calibration curve. The concentration of metallothionein-equivalents were measured by differential pulse polarography with a static mercury drop electrode using a Metrohm 694 stand and a 693 processor. The electrode system consisted of a mercury capillary working electrode, an Ag/AgCl reference electrode and a platinum auxiliary electrode. The supporting electrolyte contained 1 M NH4Cl, 1 M NH4OH and 2 mM [Co(NH3)6]Cl3.
The samples’ total protein was estimated through the method of Bradford (1976), in order to normalize biomarker responses to protein content. All colorimetric assays were performed using a Benchmark Microplate Reader (Bio-Rad).
Statistical analysis
After the invalidation of at least one of the assumptions for parametric analysis, namely homogeneity of variances (through the Levene’s test), non-parametric analyses were employed; specifically, the Mann–Whitney U test to search for inter-site differentiation, and the non-parametric Spearman’s Rank-order correlation R statistic. All variables (biomarker responses and morphometrics) were modelled through multivariate statistics, namely discriminant analysis, to determine the significance of each variable in site differentiation. A significance level α = 0.05 was set for all analyses. All statistics were performed using Statistica (StatSoft), following Zar (1998).
Integrated biomarker response (IBR)
The IBR index was computed to integrate all biomarker responses determined in both organs, according to the method described by Beliaeff and Burgeot (2002). In brief: the score (S) for each biomarker in each site and for each organ was calculated through the formula:
where S ≥ 0, since |Min| is the absolute minimum value obtained for the biomarker and:
where Z is either positive or negative, depending on the activation or inhibition of the biological effect, respectively. The standardized values Z were estimated through the mean value for each biomarker in each site (X), the mean value for each biomarker (m) and the standard deviation of X(s). The area (A) connecting two consecutive coordinates was calculated for each biomarker result in star plots, being S i and S i+1 two consecutive scores and n the number of biomarkers under analysis:
where:
The IBR was then calculated through the sum of all the areas (A) for a given site and organ.
Results
Sediment characterization
The Sado fishing grounds (Sado 1 and Sado 2) were overall more contaminated than the reference area, as inferred from the sediment samples’ contamination levels (Table 1). Still, within the Sado estuary’s areas, considerable variation was observed, especially concerning the Sado 1 area, where sediments S1 and S2 were found to be uncontaminated, i.e. presenting contamination levels similar to those from the sediments collected from the reference location (sample M). Sediment samples S3 (Sado 1, off the heavy-industry belt) and S4 and S5 (Sado 2) had the highest concentrations of contaminants. These sediments also reached values of TOM between seven and ten times higher than S1, S2 and M; as well as the lowest Eh and a high percentage of FF (more than 50 %). Sites S3, S4 and S5 yielded the highest metal and metalloid concentrations, having, in the most extreme case (as in sediment S5), Zn concentrations reaching about 300-fold the concentration found in M. These high values exceeded, in most cases, the TEL guideline, in some cases even exceeding PEL (Cu and Zn for S3 and Zn for S5). The elements of most concern were Zn and Cu, the latter attaining a 70-fold concentration in S3 relatively to M. The PAH concentrations exceeded slightly the TEL values only in S3, for the three-ring PAHs acenaphthylene and acenaphthene, the four-ring fluoranthene and pyrene and the five-ring dibenzo[a,h]anthracene. However, tPAH was not above TEL in any site. Sediment S3 also had the highest tDDT and tPCB values, although still below TEL, having, however, a pp’DDT concentration close to the TEL value of 1.19 ng g−1. All other sediments showed no relevant concentrations of organic contaminants, being all values below TEL.
Biomarker responses
There were significant differences between all sites for all studied biomarkers and distinct response patterns between digestive gland and gills, except for GST and LPO (Fig. 2). All biomarker results in the digestive glands revealed a clear differentiation between Sado 1 and Reference, whereas in gills this segregation was obtained only for the GSH/GSSG ratio and GST activity. Sites Sado 2 and Reference were differentiated by LPO and GSHt in both organs and by GSH/GSSG ratio in gills. Total GSH and GST in both organs plus the GSH/GSSG ratio in gills differentiated Sado 1 from Sado 2. The biomarker yielding highest inter-site differentiation in gills was the GSH/GSSG ratio, while for the digestive gland was GSHt. The biomarkers showing the highest differences relative to the reference site were LPO and GSHt in the digestive gland, reaching values three-fold higher in Sado 1.

Mean results of biomarker responses per site (Sado 1, Sado 2 and Reference) in digestive gland and gills (error bars indicate 95 % confidence intervals). a Lipid peroxidation (as thiobarbituric acid reactive species). b Total glutathione. c Reduced/oxidised glutathione ratio (GSH/GSSG). d Glutathione S-transferase activity. e Metallothionein-like protein concentration. Different letters mean significant differences (Mann–Whitney U, p < 0.05)
In general, the biomarker values in digestive gland were higher than in gills, except for GSHt, where values were, in average, tenfold lower in the digestive gland. In the digestive glands, LPO and the GSH/GSSG ratio, were twofold higher than in gills, whereas digestive gland MT reached tenfold gill MT in animals from Sado 1. The two organs yielded a GST response within the same order of magnitude.
Statistical integration of data
Discriminant analysis was performed by deriving four models (A–D) comprising different variables (Table 2). In model A (all variables included), size and digestive gland biomarkers yielded no significance (p > 0.05), however, gill LPO and GST provided a significant contribution to distinguish between sites. In model B (without size variables), gill LPO, GST and GSH/GSSG ratio were still significant, together with digestive gland LPO. When taking into account only digestive gland biomarkers (C), LPO and GST became significant. In the model with gill biomarkers only (D), besides LPO and GST, the GSH/GSSG ratio also became significant. There was a general tendency for LPO and GST (especially in gills) to be the most significant biomarkers for site differentiation.
All models yielded inter-site differentiation (Fig. 3). In the model including all biomarker responses in digestive gland and gills plus length and weight, a clear differentiation of the reference site was obtained, together with a good separation of both sites from Sado (Fig. 3a). In the model without length and weight (Fig. 3b), the site segregation is reduced but still achieved, and in models C and D (Fig. 3c, d, respectively) a less conspicuous site separation was observed, when comparing models A and B. Nevertheless, gill biomarkers (Fig. 3d) could better differentiate sites than digestive gland responses (Fig. 3c), especially regarding site Sado 1.

Results from discriminant analysis. Scatterplot of canonical scores for inter-site differentiation (Sado 1, Sado 2 and Reference) considering different variables in each case. a Length, weight plus all biomarkers studied in both organs (LPO, GSHt, GSH/GSSG, GST and MT). b Biomarkers only (both organs). c Digestive gland biomarkers only. d Gill biomarkers only
Correlations between all variables, independently of site, are presented in Table 3. The highest correlation (Spearman’s R = 0.94) was obtained between mantle length and total wet weight, as expected. Negative correlations were found between size variables and biomarkers, except for the GSH/GSSG ratio. The biomarkers most correlated with size (R > 0.4) were LPO in the digestive gland, GSH/GSSG ratio in gill and GST in both organs. The highest correlations (R > 0.5) between biomarkers were obtained for GST versus LPO, GSHt and MT, all in the digestive gland. All significant between-biomarker correlations (R > 0.4 and p < 0.05) were positive.
Integrated biomarker response (IBR)
The IBR values were calculated based on the following biomarker order: LPO > GSHt > GSH/GSSG > GST > MT, considering that LPO may occur before active anti-oxidant defences are triggered. Sado 1 attained the highest IBR scores, followed by Sado 2, and virtually null for reference animals, (Fig. 4). Lipid peroxidation was one of the most significant biomarkers, especially in the digestive gland, in animals from Sado 2. Overall, the integrated responses obtained for site Sado 1 were higher in gills, while in digestive gland they were highest in Sado 2.

Integrated biomarker response (IBR) star plots per site (Sado 1, Sado 2 and Reference), considering all biomarkers and both organs. a Scores (S) of digestive gland biomarkers for each site. b Scores of gill biomarkers for each site. c Biomarker scores combining both organs, for each site. d IBR values for each site considering each organ separately and combined
Discussion
The present study revealed different biomarker patterns between S. officinalis collected from the three fishing areas, including those allocated within the same estuary, Sado 1 and Sado 2. The Sado fishing areas hold point differences regarding sediment contamination, especially by PAHs, which attain higher levels in the sediments close to the heavy-industry belt adjacent to Sado 1. Given the overall moderate levels of contamination of the Sado estuary, the species proved sensitive to environmental contamination even when other molluscs, namely the bivalve Ruditapes decussatus, collected from the same areas during the same sampling effort, yielded unclear biomarker responses (Carreira et al. 2013), which contradicts findings by other authors. In fact, the clam R. decussatus has been suggested to be very sensitive to environmental contaminants and proposed for biomonitoring as an effective surrogate for mussels in areas where these are not abundant (Bebianno et al. 2004). This may indicate that, as in the Sado estuary, clams may be less efficient bioindicator organisms when complex mixtures of toxicants are involved. Additionally, there were differences in biomarker responses between S. officinalis digestive glands and gills. Digestive gland biomarkers were overall in better agreement with environmental contamination, although gills evidenced increased sensitivity while disclosing a distinct pattern of contamination between the industrial and agricultural areas of the estuary. The findings are further sustained by the IBR results, a complementary leverage to the results obtained by the statistical analyses, and an expedient tool for decision makers which examines differences in response between populations and test groups by integrating biomarker responses into a single index (Broeg and Lehtonen 2006). The IBR estimates (Fig. 4) evidenced Sado 1 as the globally most contaminated site, regardless of high heterogeneity in sediment contamination profiles. However, caution should be taken when interpreting IBR results, since calculations reflect the choice of biomarker hierarchy, the number of surveyed biomarkers and their relative weight, to which is added the fact that it is a dynamic index, and thus cannot be used in direct comparisons between distinct studies (Damiens et al. 2007; Tsangaris et al. 2011; Serafim et al. 2012). Additionally, such indexes reflect only a qualitative assessment and thus should not serve as a single-stand Line-Of-Evidence in biomonitoring, management and decision-making processes (Serafim et al. op. cit.). Still, in the present study the IBR values were consistent with the differences between sites and revealed patterns of response between the two surveyed organs that are in good agreement with multivariate statistics, which calls for its applicability in studies when the same batch of biomarkers are determined in organisms from distinct, albeit proximal, locations.
Contamination profiles
The contrast between sediment contamination profiles and SGQ’s allowed detecting the compounds potentially posing higher risk to the biota. The results retrieved from Carreira et al. (2013), are consistent with previous sediment analyses performed in the Sado estuary and compiled by Costa et al. (2012), revealing that the Sado estuary is moderately contaminated, impacted by an intricate mixture of contaminants and that metals, especially Zn and Cu, are the contaminants of highest risk. The most significant organic contaminants in the sediments from the Sado estuary were PAHs, similarly to those described by previous works in the area (Costa et al. 2011, 2012). Still, the overall levels were relatively low, with few individual contaminants approaching the TEL guideline, such as the hazardous (genotoxic and carcinogenic) benzo[a]anthracene and benzo[a]pyrene, especially in sediment S3, from the Sado 1 fishing grounds, located near the city of Setúbal and its adjacent heavy-industry belt. It should also be noted that the physico-chemical characteristics of the sediments play a major role in contaminant bioavailability. The most contaminated sites had also higher FF and TOM and lower Eh, similarly to previous studies (e.g. Caeiro et al. 2005, 2009; Costa et al. 2008b). Even if high proportions of FF and TOM act as a trap for contaminants, therefore rendering them less immediately available to aquatic organisms, disturbance, combined with low Eh, may favour toxicant speciation and release (see for instance Caccia et al. 2003; Eggleton and Thomas 2004; Du Laing et al. 2009).
The contamination levels of sediments from the Sado estuary revealed considerable spatial variation, as previously evidenced by Caeiro et al. (2005, 2009). Sediments S1 and S2, from Sado 1 fishing grounds, were essentially clean, even though sediments S3, from a nearby sampling site, were found the overall most contaminated. Sediments S1 and S2 are in fact sandy shellfish beds subjected to high hydrodynamics and high oceanic influence, which most likely contributed to the low contamination levels. However, cuttlefish from this area demonstrated biomarker responses consistent, comparatively, to higher contamination patterns. In comparison, clams (filter-feeders) collected from these sandbanks yielded reduced responses, likely reflecting the low contamination pattern of their immediate surroundings (Carreira et al. 2013; Costa et al. 2013). On the contrary, cuttlefish are territorial predators, thus acting as indicators of a broader biogeographical area that includes, in the present situation, both clean and contaminated sediments. Also, sediments S4 and S5, from Sado 2, presented more similar contamination patterns, although distinct from S2, and eliciting in the animals biomarker responses lower than those from Sado 1 but still higher than the reference cuttlefish. Previous data from this area is scarce; however, analyses performed on sediments from the river mouth, close to these sites, yielded the highest values for Zn and Cd in the estuary (Cortesão and Vale 1995), which is consistent with the present results. In fact, some of the analysed metals attained higher concentrations in sites S4 and S5 (especially S5), than in site S3. Possible explanations may derive from pyrite mining along the river basin (Vale and Cortesão 1989), the fertilizers applied in agriculture grounds upstream (especially extensive rice farming), some of which may have metals (e.g. Cr, Cu, Cd, Zn and Ni) as constituents (Nziguheba and Smolders 2008), and pesticide use (Cerejeira et al. 2003; Villaverde et al. 2008).
Biomarker approach
All biomarkers were responsive, especially in the digestive gland, indicating sensitivity to low-moderate levels of mixtures of both organic and inorganic xenobiotics. Also, the combination of biomarker responses yielded a clear distinction between the three fishing areas (Fig. 3). Biomarker responses were overall consistent with sediment contamination, especially in the digestive gland, with higher levels of response and damage (the last given by LPO) being measured in cuttlefish from site Sado 1 (near the heavy-industry area). Nonetheless, the differences between sites were generally more pronounced in the gills (with the exception of MT induction). Some of the differences between the two organs may be explained, by the fact that digestive glands likely tend to reflect a more chronic exposure than gills. Metallothionein induction, for instance, was higher in the digestive gland, probably due to the organ’s ability to store higher levels of metals. Conversely, gills reflect the pathway for metal uptake and short-time storage, as previously observed in fish and even in the cephalopod Octopus vulgaris (Hamza-Chaffai et al. 1995; Raimundo et al. 2010).
Overall, oxidative stress-related biomarkers, especially GST and LPO were the most relevant biomarkers for site differentiation, evidenced by statistical analyses and IBR. This finding is consistent with the higher levels of PAHs found in Sado 1, since cuttlefish sampled at this location yielded the highest responses. In fact, LPO and GST have been suggested as trustworthy biomarkers of contaminated sediments (Moreira et al. 2006). Correlations were found between digestive gland biomarker responses, which contribute to sustain the premise of active and integrative responses to contamination occurring in this organ. For instance, the positive correlation between MT and LPO might indicate that the protective antioxidant role of MT (yet another function of these proteins) was not sufficient to prevent oxidative stress, contrary to what Correia et al. (2002) found in amphipod Gammarus locusta exposed to Cu and sediments from the Sado estuary. In one of the few studies on biomarkers (including LPO) performed on S. officinalis, Zielinski and Pörtner (2000) also found the gills to be sensitive and responsive to oxidative stress, as in the present study, however, the test variable was age and not toxicological challenge.
Biomarkers such as LPO are known to be positively influenced by endogenous variables like age and size (Zielinski and Pörtner 2000), plus exogenous variables like seasonality (Company et al. 2006; Pytharopoulou et al. 2008). However, regarding exogenous variables, even if seasonal differences can justify some differences between Sado 1 and Sado 2, they cannot single-handedly explain the differences between Sado 1 and Reference; since the specimens were collected during the same season. Regarding environmental contamination, LPO may be induced by metals in invertebrates (Correia et al. 2002; Pytharopoulou et al. 2008), as supported by the present findings. Oddly, the inverse was reported by Viarengo et al. (1990). Some authors stated that the link between LPO and exposure to metals is only significant when contamination levels are high (Pedrajas et al. 1995; Martín-Díaz et al. 2009). In general, regardless of the contaminant and concentration, long-term exposures are thought to be necessary to induce lipid peroxidation (Gravato et al. 2010), which is suspected to occur in feral animals, as in the present study.
Nevertheless, as previously noted, it must be pointed out that reference animals were collected from an off-coast fishing area, which implies a distinct set of unaccounted environmental variables, from water salinity, pH and temperature to prey type and availability. These variables likely influenced the animals’ physiological status and, therefore, are probable confounding factors of biomarker responses. As such, caution is mandatory when interpreting potential background levels of biomarker responses from these animals and it is clear that further research, meaning surveys in unpolluted estuarine habitats, is still needed to disclose the true sensitivity of the species toward toxicological challenge.
To cope with exposure to xenobiotics, the organisms may trigger defences towards oxidative stress often resulting from the catalysis/excretion of organic xenobiotics, usually hydrophobic, like PAHs. After phase I enzymes having biotransformed compounds (such as some PAHs and dioxins), the phase II enzyme GST enters in action to catalyse GSH conjugation with electrophilic toxicants or hazardous contaminant metabolites such as PAH epoxides. On the other hand, GSH may scavenge ROS, becoming oxidised itself (see van der Oost et al. 2003; Martín-Díaz et al. 2008, Oliveira et al. 2009, for reviews). Thus, the reduction in the GSH/GSSG ratio in cuttlefish from Sado (more pronounced in the gills), is in good accordance with previous research relating reduced GSH/GSSG ratio in aquatic organisms exposed to organic and inorganic contaminants (see van der Oost et al. 1996; Moreira et al. 2006; Martins et al. 2012; Taylor and Maher 2012). Even though GSHt was not very significant for inter-site differentiation, it was strongly correlated with the GST activity levels in both organs, as expected.
The activity of the phase II enzyme GST was similar in both organs. The adequacy of GST activity as a biomarker of exposure has already been demonstrated in a wide range of organisms, including bivalve molluscs (e.g. Hoarau et al. 2004; Moreira et al. 2006; Fonseca et al. 2011a). Its modulation by other environmental factors besides pollution, such as seasonality, has been dismissed by some authors in experiments performed in fish (Kopecka and Pempkowiak 2008; Fonseca et al. 2011a); in opposition to Serafim et al. (2012). It is known that induction of GST activity may counteract oxidative stress triggered by exposure to metals such as Cd, Cu, As, Pb and Zn (Damiens et al. 2007; Martín-Díaz et al. 2009; García-Alonso et al. 2011; Ramos-Gómez et al. 2011), and organochlorine compounds, like PCBs and DDTs, in clams (Hoarau et al. 2001, 2004). However, PAHs have been reported to cause either GST inhibition (Fonseca et al. 2011b) or induction (Hoarau et al. 2001; Damiens et al. 2007; Gravato et al. 2010). Interestingly, even a case of no correlation between these toxicants and this enzyme’s activity has been reported (García-Alonso et al. 2011). In the present study, PAH-induced GST activity is suggested, since the higher sediment PAH levels observed in Sado 1 sediments were linked to higher GST activity, in both organs.
Metallothioneins (MTs) are important proteins in the regulation and detoxification of both essential and non-essential metals. Still, MTs may also protect the cells against oxidative stress and function as radical scavengers (Buico et al. 2008). The present study revealed a modest, however significant, MT response in the digestive gland of cuttlefish collected off the Sado estuary’s industrial belt, where the highest levels of metals, especially Cu and Zn (known MT inducers) were recorded. Importantly, the significant MT response contradicts previous studies with clams collected from this area (Carreira et al. 2013) and different species of fish exposed to sediments from the Sado Estuary (Costa et al. 2008a, 2009). This may indicate that the MT response in cuttlefish retains specificity when complex mixtures of organic and metallic toxicants are involved, which is usually a critical confounding factor when surveying this biomarker (e.g. Costa et al. 2009, 2012).
The gills and liver (in fish) or digestive gland (molluscs) are the most usually surveyed organs for MT induction, due to their role in metal uptake and bioaccumulation/detoxification, respectively (Hamza-Chaffai et al. 1995). The MT response was not significant in gills, contrary to other studies performed with clams and fish (Hamza-Chaffai et al. 1997; Bebianno and Serafim 2003; Oliveira et al. 2009). This occurrence might be explained by overall low levels of metals in the estuary, combined with the plausible adaptation of cuttlefish to their environment. However, due to the strong correlation in the digestive gland between MT and two of the most significant biomarkers studied (GST and LPO) a link between MT and environmental contamination may be disclosed. It is likely that the higher MT levels in the cuttlefish digestive gland relate with a higher predisposition of this organ to bioaccumulate important levels of metals like Cd, Cu and Zn, as known to occur in cephalopods (Miramand and Bentley 1992; Raimundo et al. 2010). Interactions between contaminants (including between metals and between metals and organic compounds) can also modulate MT expression, often inhibiting the response (e.g. Roméo et al. 1997; Risso-de Faverney et al. 2000; Majumder et al. 2003). In fact, many authors reported inconsistencies in the MT response when acting as a potential biomarker of exposure to metals (Mouneyrac et al. 2002; Pytharopoulou et al. 2008; Serafim et al. 2012). Metallothionein induction is also known to occur as a function of animal size (Hamza-Chaffai et al. 1995), hypoxic stress (Sampaio et al. 2008) and may even be elicited by organic contaminants (Costa et al. 2009). Regardless of all these contradictions and confounding factors, it is important to emphasize that most of the studies were performed with fish, not cephalopods.
Concluding remarks
Sepia officinalis revealed to be a sensitive bioindicator of the effects elicited by mixtures of toxicants, even if present at moderate concentrations. The cuttlefish, a marine species known to occupy brackish water ecosystems at least during part of its life cycle, revealed to be a potential candidate for the monitoring of transition ecosystems, considering the link between biomarkers and environmental contamination in animals collected from impacted areas. However, further research is still needed to understand the adaptation mechanisms of the species to its habitat and their effect on biomarker responses, since it was not possible, in the present study, to consider a reference location within an estuarine environment. Additionally, gill biomarkers evidenced higher contrast between sites, whereas digestive gland responses were overall more consistent with contamination, which may result from two main aspects: (1) the differential sensitivity of both organs; (2) the effect of unknown exogenous variables, such as unsurveyed toxicants, hydrology and season-related parameters. In any case, the current findings indicate a clear response to environmental stressors, with emphasis on oxidative stress, which is consistent with higher levels of PAHs in the sediments near the Sado’s industrial belt. Still, unlike recent research on fish and bivalves in the area, even MT induction retained some degree of specificity to metals. The species was proven sensitive enough to distinguish adjacent areas within the same eco-geographical unit. This, combined with its wide ecological representation and high commercial value, makes the species a good candidate for practical applications within the EU’s Marine Strategy Framework Directive in SW Europe coastal ecosystems, likely combining the characteristics of the two groups of organisms that constitute the basis of most biomonitoring strategies in Europe: bivalve molluscs and fish.
References
Bebianno MJ, Serafim MA (2003) Variation of metallothionein and metal concentrations in a natural clam population of Ruditapes decussatus. Arch Environ Contam Toxicol 44:53–56
Bebianno MJ, Géret F, Hoarau P, Serafim MA, Coelho MR, Gnassia-Barelli M, Roméo M (2004) Biomarkers in Ruditapes decussatus: a potential bioindicator species. Biomarkers 9:305–330
Beliaeff B, Burgeot T (2002) Integrated biomarker response: a useful tool for ecological risk assessment. Environ Toxicol Chem 21:1316–1322
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Broeg K, Lehtonen KK (2006) Indices for the assessment of environmental pollution of the Baltic Sea coasts: integrated assessment of a multi-biomarker approach. Mar Pollut Bull 53:508–522
Buico A, Cassino C, Dondero F, Vergani L, Osella D (2008) Radical scavenging abilities of fish MT-A and mussel MT-10 metallothionein isoforms: an ESR study. J Inorg Biochem 102:921–927
Bustamante P, Bertrand M, Boucaud-Camou E, Miramand P (2006) Subcellular distribution of Ag, Cd Co., Cu, Fe, Mn, Pb and Zn in the digestive gland of the common cuttlefish Sepia officinalis. J Shellfish Res 25:987–993
Caccia VG, Sillero FJ, Palanques A (2003) The distribution of trace metals in Florida Bay sediments. Mar Pollut Bull 46:1420–1433
Caeiro S, Costa MH, Ramos TB, Fernandes F, Silveira N, Coimbra AP, Medeiros G, Painho M (2005) Assessing sediment heavy metals contamination in Sado Estuary: an index analysis approach. Ecol Indic 5:151–169
Caeiro S, Costa MH, DelValls A, Repolho T, Gonçalves M, Mosca A, Coimbra AP, Ramos TB, Painho M (2009) Ecological risk assessment of sediment management areas: application to Sado Estuary, Portugal. Ecotoxicology 18:1165–1175
Carreira S, Costa PM, Martins M, Lobo J, Costa MH, Caeiro S (2013) Ecotoxicological heterogeneity in transitional coastal habitats assessed through the integration of biomarkers and sediment contamination profiles: a case study with a commercial clam. Arch Environ Contam Toxicol 64:97–109
Cerejeira MJ, Viana P, Batista S, Pereira T, Silva E, Valério MJ, Silva A, Ferreira M, Silva-Fernandes AM (2003) Pesticides in Portuguese surface and ground waters. Water Res 37:1055–1063
Chapman PM (2007) Determining when contamination is pollution: weight of evidence determinations for sediments and effluents. Environ Int 33:492–501
Company R, Serafim A, Cosson R, Fiala-Médioni A, Dixon D, Bebianno JM (2006) Temporal variation in the antioxidant defence system and lipid peroxidation in the gills and mantle of hydrothermal vent mussel Bathymodiolus azoricus. Deep-Sea Res Pt I 53:1101–1116
Correia AD, Lima G, Costa MH, Livingstone DR (2002) Studies on biomarkers of copper exposure and toxicity in the marine amphipod Gammarus locusta (Crustacea): I. Induction of metallothionein and lipid peroxidation. Biomarkers 7:422–437
Cortesão C, Vale C (1995) Metals in sediments of the Sado estuary, Portugal. Mar Pollut Bull 30:34–37
Costa PM, Repolho T, Caeiro S, Diniz ME, Moura I, Costa MH (2008a) Modelling metallothionein induction in the liver of Sparus aurata exposed to metal-contaminated sediments. Ecotoxicol Environ Saf 71:117–124
Costa PM, Lobo J, Caeiro S, Martins M, Ferreira AM, Caetano M, Vale C, DelValls TA, Costa MH (2008b) Genotoxic damage in Solea senegalensis exposed to sediments from the Sado Estuary (Portugal): effects of metallic and organic contaminants. Mutat Res 654:29–37
Costa PM, Caeiro S, Diniz M, Lobo J, Martins M, Ferreira AM, Caetano M, Vale C, DelValls TÁ, Costa MH (2009) Biochemical endpoints on juvenile Solea senegalensis exposed to estuarine sediments: the effects of contaminant mixtures on metallothionein and CYP1A induction. Ecotoxicology 18:988–1000
Costa PM, Neuparth T, Caeiro S, Lobo J, Martins M, Ferreira AM, Caetano M, Vale C, DelValls TÁ, Costa MH (2011) Assessment of the genotoxic potential of contaminated estuarine sediments in fish peripheral blood: laboratory versus in situ studies. Environ Res 111:25–36
Costa PM, Caeiro S, Vale C, DelValls TÁ, Costa MH (2012) Can the integration of multiple biomarkers and sediment geochemistry aid solving the complexity of sediment risk assessment? A case study with a benthic fish. Environ Pollut 161:107–120
Costa PM, Carreira S, Costa MH, Caeiro S (2013) Development of histopathological indices in a commercial marine bivalve (Ruditapes decussatus) to determine environmental quality. Aquat Toxicol 126:442–454
Damiens G, Gnassia-Barelli M, Loquès F, Roméo M, Salbert V (2007) Integrated biomarker response index as a useful tool for environmental assessment evaluated using transplanted mussels. Chemosphere 66:574–583
Du Laing G, Rinklebe J, Vandecasteele B, Meers E, Tack FMG (2009) Trace metal behaviour in estuarine and riverine floodplain soils and sediments: a review. Sci Total Environ 407:3972–3985
Eggleton J, Thomas KV (2004) A review of factors affecting the release and bioavailability of contaminants during sediment disturbance events. Environ Int 30:973–980
Fonseca VF, França S, Vasconcelos RP, Serafim A, Company R, Lopes B, Bebianno MJ, Cabral HN (2011a) Short-term variability of multiple biomarker response in fish from estuaries: Influence of environmental dynamics. Mar Environ Res 72:172–178
Fonseca VF, França S, Serafim A, Company R, Lopes B, Bebianno MJ, Cabral HN (2011b) Multi-biomarker responses to estuarine habitat contamination in three fish species: Dicentrarchus labrax, Solea senegalensis and Pomatoschistus microps. Aquat Toxicol 102:216–227
García-Alonso J, Greenway GM, Munshi A, Gómez JC, Mazik K, Knight AW, Hardege JD, Elliott M (2011) Biological responses to contaminants in the Humber Estuary: disentangling complex relationships. Mar Environ Res 71:295–303
Gravato C, Guimarães L, Santos J, Faria M, Alves A, Guilhermino L (2010) Comparative study about the effects of pollution on glass and yellow eels (Anguilla anguilla) from the estuaries of Minho, Lima and Douro Rivers (NW Portugal). Ecotoxicol Environ Saf 73:524–533
Guerra A (2006) Ecology of Sepia officinalis. Life Environ 56:97–107
Guerra Á, Allcock L, Pereira J (2010) Cephalopod life history, ecology and fisheries: an introduction. Fish Res 106:117–124
Hamza-Chaffai A, Cosson RP, Amiard-Triquet C, El-Abed A (1995) Physico-chemical forms of storage of metals (Cd, Cu and Zn) and metallothionein-like proteins in gills and liver of marine fish from the Tunisian coast: ecotoxicological consequences. Comp Biochem Physiol C 111:329–341
Hamza-Chaffai A, Amiard-Triquet C, Abed AE (1997) Metallothionein-like protein: Is it an efficient biomarker of metal contamination? A case study based on fish from the Tunisian coast. Arch Environ Contam Toxicol 33:53–62
Hoarau P, Gnassia-Barelli M, Roméo M, Girard JP (2001) Differential induction of GST in the clam Ruditapes decussatus exposed to organic compounds. Environ Toxicol Chem 20:523–529
Hoarau P, Garello G, Gnassia-Barelli M, Roméo M, Girard JP (2004) Effect of three xenobiotic compounds on glutathione S-transferase in the clam Ruditapes decussatus. Aquat Toxicol 68:87–94
Kopecka J, Pempkowiak J (2008) Temporal and spatial variations of selected biomarker activities in flounder (Platichthys flesus) collected in Baltic proper. Ecotoxicol Environ Saf 70:379–391
Lyons BP, Thain JE, Stentiford GD, Hylland K, Davies IM, Vethaak AD (2010) Using biological effects tools to define Good Environmental Status under the European Union Marine Strategy Framework Directive. Mar Pollut Bull 60:1647–1651
Macdonald DD, Carr S, Calder F, Long E, Ingersoll C (1996) Development and evaluation of sediment quality guidelines for Florida coastal waters. Ecotoxicology 5:253–278
Majumder S, Ghoshal K, Summers D, Bai S, Datta J, Jacob ST (2003) Chromium (IV) down-regulates heavy-metal induced metallothionein gene transcription by modifying transactivation potential of the key transcription factor, metal-responsive transcription factor 1. J Biol Chem 278:26216–26226
Martín-Díaz ML, DelValls TÁ, Riba I, Blasco J (2008) Integrative sediment quality assessment using a biomarker approach: review of 3 years of field research. Cell Biol Toxicol 24:513–526
Martín-Díaz ML, Blasco J, Sales D, DelValls TÁ (2009) The use of a kinetic biomarker approach for in situ monitoring of littoral sediments using the crab Carcinus maenas. Mar Environ Res 68:82–88
Martins M, Costa PM, Ferreira AM, Costa MH (2012) Impact of remobilized contaminants in Mytilus edulis during dredging operations in a harbour area: bioaccumulation and biomarker responses. Ecotoxicol Environ Saf 85:96–103
Miramand P, Bentley D (1992) Concentration and distribution of heavy metals in tissues of two cephalopods, Eledone cirrhosa and Sepia officinalis, from the French coast of the English Channel. Mar Biol 114:407–414
Moreira SM, Lima I, Ribeiro R, Guilhermino L (2006) Effects of estuarine sediment contamination on feeding and on key physiological functions of the polychaete Hediste diversicolor: laboratory and in situ assays. Aquat Toxicol 78:186–201
Mouneyrac C, Amiard JC, Amiard-Triquet C, Cottier A, Rainbow PS, Smith BD (2002) Partitioning of accumulated trace metals in the talitrid amphipod crustacean Orchestia gammarellus: a cautionary tale on the use of metallothionein-like proteins as biomarkers. Aquat Toxicol 57:225–242
Neves A, Cabral H, Sequeira V, Figueiredo I, Moura T, Gordo LS (2009) Distribution patterns and reproduction of the cuttlefish, Sepia officinalis in the Sado estuary (Portugal). J Mar Biol Assoc UK 89:579–584
Nziguheba G, Smolders E (2008) Inputs of trace elements in agricultural soils via phosphate fertilizers in European countries. Sci Total Environ 390:53–57
Oliveira M, Maria VL, Ahmad I, Serafim A, Bebianno MJ, Pacheco M, Santos MA (2009) Contamination assessment of a coastal lagoon (Ria de Aveiro, Portugal) using defence and damage biochemical indicators in gill of Liza aurata—an integrated biomarker approach. Environ Pollut 157:959–967
Paleček E, Pechan Z (1971) Estimation of nanogram quantities of proteins by pulse polarographic techniques. Anal Biochem 42:59–71
Pedrajas JR, Peinado J, Lopez-Barea J (1995) Oxidative stress in fish exposed to model xenobiotics. Oxidatively modified forms of Cu, Zn-superoxide dismutase as potential biomarkers. Chem-Biol Interact 98:267–282
Picado A, Bebianno MJ, Costa MH, Ferreira A, Vale C (2007) Biomarkers: a strategic tool in the assessment of environmental quality of coastal waters. Hydrobiologia 587:79–87
Pytharopoulou S, Sazakli E, Grintzalis K, Georgiou CD, Leotsinidis M, Kalpaxis DL (2008) Translational responses of Mytilus galloprovincialis to environmental pollution: integrating the responses to oxidative stress and other biomarker responses into a general stress index. Aquat Toxicol 89:18–27
Raimundo J, Costa PM, Vale C, Costa MH, Moura I (2010) Metallothioneins and trace elements in digestive gland, gills, kidney and gonads of Octopus vulgaris. Comp Biochem Physiol C 152:139–146
Ramos-Gómez J, Coz A, Viguri JR, Luque Á, Martín-Díaz ML, DelValls TÁ (2011) Biomarker responsiveness in different tissues of caged Ruditapes philippinarum and its use within an integrated sediment quality assessment. Environ Pollut 159:1914–1922
Risso-de Faverney C, Lafaurie M, Girard J-P, Rahmani R (2000) Effects of heavy metals and 3-methylcholanthrene on expression and induction of CYP1A1 and metallothionein levels in trout (Oncorhynchus mykiss) hepatocyte cultures. Environ Toxicol Chem 19:2239–2248
Roméo M, Cosson RP, Gnassia-Barelli M, Risso C, Stien X, Lafaurie M (1997) Metallothionein determination in the liver of the sea bass Dicentrarchus labrax treated with copper and B(a)P. Mar Environ Res 44:275–284
Sampaio FG, Boijink CL, Oba ET, dos Santos LRB, Kalinin AL, Rantin FT (2008) Antioxidant defenses and biochemical changes in pacu (Piaractus mesopotamicus) in response to single and combined copper and hypoxia exposure. Comp Biochem Physiol C 147:43–51
Serafim A, Company R, Lopes B, Fonseca VF, Franc S, Vasconcelos RP, Bebianno MJ, Cabral HN (2012) Application of an integrated biomarker response index (IBR) to assess temporal variation of environmental quality in two Portuguese aquatic systems. Ecol Indic 19:215–225
Taylor AM, Maher WA (2012) Exposure-dose-response of Anadara trapezia to metal contaminated estuarine sediments. 1. Cadmium spiked sediments. Aquat Toxicol 109:234–242
Tsangaris C, Hatzianestis I, Catsiki V-A, Kormas KA, Strogyloudi E, Neofitou C, Andral B, Galgani F (2011) Active biomonitoring in Greek coastal waters: application of the integrated biomarker response index in relation to contaminant levels in caged mussels. Sci Total Environ 412(413):359–365
Uchiyama M, Mihara M (1978) Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal Biochem 86:271–278
Vale C, Cortesão C (1989) Partitioning of Cd, Cu, Zn, Mn, and Fe to suspended solids of an upper estuary receiving a mine input (Sado River estuary). In: Vernet J-P (ed) Heavy metals in the environment. CED Consultants Ltd., Edinburgh, pp 379–382
van der Oost R, Goksøyr A, Celander M, Heida H, Vermeulen NPE (1996) Biomonitoring aquatic pollution with feral eel (Anguilla anguilla). II. Biomarkers: pollution-induced biochemical responses. Aquat Toxicol 36:189–222
van der Oost R, Beyer J, Vermeulen NPE (2003) Fish bioaccumulation and biomarkers in environmental risk assessment: a review. Environ Toxicol Pharmacol 13:57–149
Vasconcelos RP, Reis-Santos P, Fonseca V, Maia A, Ruano M, França S, Vinagre C, Costa MJ, Cabral H (2007) Assessing anthropogenic pressures on estuarine fish nurseries along the Portuguese coast: a multi-metric index and conceptual approach. Sci Total Environ 374:199–215
Viarengo A, Canesi L, Pertica M, Poli G, Moore MN, Orunesu M (1990) Heavy metal effects on lipid peroxidation in the tissues of Mytilus galloprovincialis Lam. Comp Biochem Physiol C 97:37–42
Villaverde J, Hildebrandt A, Martínez E, Lacorte S, Morillo E, Maqueda C, Viana P, Barceló D (2008) Priority pesticides and their degradation products in river sediments from Portugal. Sci Total Environ 390:507–513
Zar JH (1998) Biostatistical analysis, 4th edn. Prentice Hall, Upper Saddle River
Zielinski S, Pörtner H-O (2000) Oxidative stress and antioxidative defense in cephalopods: A function of metabolic rate or age? Comp Biochem Physiol B 125:147–160
Acknowledgments
P.M. Costa was supported by the Portuguese Science and Technology Foundation (FCT) through the grant SFRH/BPD/72564/2010. The present research was financed by FCT and co-financed by the European Community FEDER through the program COMPETE (project HERA, Reference no. PTDC/SAU-ESA/100107/2008). The authors thank C. Gonçalves and J. Lobo (IMAR); M. Martins (IMAR/IPMA); G. Silva and L. Jacinto (Docapesca) and Mr. and Mrs. J. Bacalhau for their important assistance.
Conflict of interest
The authors declare that there are no conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rodrigo, A.P., Costa, P.M., Costa, M.H. et al. Integration of sediment contamination with multi-biomarker responses in a novel potential bioindicator (Sepia officinalis) for risk assessment in impacted estuaries. Ecotoxicology 22, 1538–1554 (2013). https://doi.org/10.1007/s10646-013-1140-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-013-1140-3