
Abstract
This study investigated sub-lethal effects and detoxification processes activated in free-ranging Red Knots (RKs) (Calidris canutus) from the Pertuis Charentais on the Atlantic coast of France, and compared the results with previous data obtained on another shorebird species, the Black-tailed Godwit (Limosa limosa). The concentrations of 13 trace elements (Ag, As, Cd, Co, Cr, Cu, Fe, Hg, Mn, Ni, Pb, Se, Zn) were assessed in the liver, kidneys, muscle and feathers. Stable isotope analyses of carbon and nitrogen were carried out to determine whether differences in diet explained variations in elemental uptake. The mRNA expression of relevant genes (cytochrome c oxidase 1, acetyl-CoA carboxylase, Cu/Zn and Mn superoxide dismutase, catalase, metallothionein, malic enzyme), antioxidant enzyme activities (catalase, glutathione peroxidase (GPx), superoxide dismutase), and metallothionein (MT) levels were investigated to shed light on trace element detoxification and toxic effects. Although Red Knots were characterized by elevated As and Se concentrations which were potentially toxic, most elements were usually below toxicity threshold levels. The results strongly suggested a dietary specialization of Red Knots, with individuals feeding on higher trophic status prey experiencing higher As, Hg and Se burdens. Red Knots and Godwits also showed discrepancies in elemental accumulation and detoxification processes. Higher As and Se concentrations in Red Knots enhanced catalase gene expression and enzyme activity, while Godwits had higher Ag, Cu, Fe and Zn levels and showed higher MT production and GPx activity. The results strongly suggest that detoxification pathways are essentially trace element- and species-specific.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Over the last three decades, the massive dispersal of toxic compounds into coastal areas as a result of human activities has greatly impacted and degraded those ecosystems, which rank today among the most endangered ecosystems in the world (Vitousek et al. 1997; Halpern et al. 2008). The loss and degradation of habitats at different points along shorebird flyways have partly caused their decline (Stroud et al. 2006). On a global scale, with respect to known population trends, 46 % of wader populations are in decline (Delany and Scott 2006; Stroud et al. 2006). The Red Knot (Calidris canutus) is a shorebird species highly specialized for long distance migrations. Two subspecies, C. c. islandica (the so-called Nearctic Knots) and C. c. canutus (the so-called Afro-Siberian Knots) reach the north of Europe in late July and August after breeding, where they possibly mingle on some sites like the Wadden Sea or the Pertuis Charentais (Piersma et al. 1993; Nebel et al. 2000; Koopman 2002; Bocher et al. 2012). Demographic studies in the Wadden Sea demonstrated the loss of 25 % of Red Knots C. c. islandica population between 1997 and 2003 (van Gils et al. 2006). Similarly, C. c. canutus is considered to possibly decreasing in numbers (Spaans et al. 2011).
The Pertuis Charentais on the French central Atlantic coast is the main wintering and/or stopover site during Spring and Fall migration for shorebirds in France (Mahéo 2011). This area is subject to trace element inputs through river discharges (Pigeot et al. 2006) which lead to subsequent bioaccumulation in the macrofaunal community (Bustamante and Miramand 2004, 2005) and, therefore, in the prey of shorebirds. Consequently, shorebirds could be exposed during the wintering period to non-essential (and thus potentially toxic beyond a threshold) elements such as silver (Ag), cadmium (Cd), lead (Pb) and, mercury (Hg) and also to essential elements that could be toxic at high levels such as arsenic (As), cobalt (Co), chromium (Cr), copper (Cu), iron (Fe), manganese (Mn), nickel (Ni), selenium (Se) and zinc (Zn).
Reporting population declines at the community scale provides a late-stage signal of potential contaminant effects. In this context, investigating sub-lethal effects of contaminants in wild individuals to provide early diagnosis of endangered species poses a major challenge (Adams et al. 2001). Nonetheless, in contrast to terrestrial birds and some waterbirds (e.g. Anatidae) for which oxidative stress related to trace elements has been demonstrated in the wild or experimentally (Hoffman et al. 2000; Hoffman 2002; Mateo et al. 2003; Berglund et al. 2007; Lucia et al. 2009), cellular damages due to trace elements are relatively unexplored in shorebirds. Filling in these gaps in the data is crucial for these endangered species. Yet, a previous study on the Black-tailed Godwit (Limosa limosa) from the Pertuis Charentais demonstrated possible production of reactive oxygen species, and suggested that in order to counter trace element toxicity, additional energy needs are required, especially to sustain detoxification mechanisms (Lucia et al. 2012). However, even though Red Knots and Godwits are both specialized to feed on mudflats, their morphology and feeding behaviours are different. A comparison of these shorebird species could, therefore, provide new clues on sub-lethal effects and detoxification mechanisms implemented by free-ranging birds and reveal possible species-specific responses to trace element accumulation.
The objective of the current study was to compare the detoxification processes and sub-lethal effects of trace elements in free-ranging Red Knots to previous results obtained on Black-tailed Godwits (Lucia et al. 2012) wintering or staging in the Pertuis Charentais. The concentrations of the 13 trace elements cited above were determined in the liver, kidneys, muscle and feathers of Red Knots. The first two tissues are specifically involved in detoxification of trace elements, while muscle could function as an important storage tissue for some elements and feathers as a pathway for their excretion (Lewis and Furness 1991). Since ingestion of food is considered to be the main exposure pathway for shorebirds, stable isotopes of carbon and nitrogen were also analysed in Red Knots to determine whether differences in trophic position or diet explained variations in elemental uptake (Jardine et al. 2006). Second, seven genes were selected in order to study trace element effects on key metabolic functions such as lipogenesis and mitochondrial metabolism as well as detoxification processes and oxidative stress responses. Mitochondrial metabolism was investigated using cytochrome c oxidase subunit 1 (cox1) and lipogenesis was investigated through expression of acetyl-CoA carboxylase (acc) and malic enzyme (me). Moreover, organisms display different mechanisms to counter trace element toxicity. Among them, metallothioneins (MT), low molecular weight cysteine-rich proteins, are known to be involved in homeostasis of essential metals such as Cu and Zn as well as in protection against toxic metals such as Ag, Cd and Hg. Another means employed to counter trace element toxicity is to increase antioxidant defenses such as Cu/Zn and Mn superoxide dismutase (sod1 and sod2, respectively) and catalase (cat). Expression of these genes and the activities of the corresponding enzymes were studied, as well as the activity of glutathione peroxidase (GPx). Finally, some elements such as Cd induce lipid peroxidation of hepatocyte membranes (Stohs and Bagchi 1995). Lipid peroxidation was therefore investigated via malondialdehyde (MDA) contents.
Materials and methods
Study site and sampling
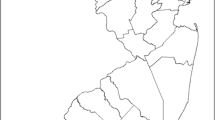
15 Red Knots that accidentally died during mist net capture for ringing purposes, were collected between October 2007 and February 2011 in the Pertuis Charentais, on the Atlantic coast of France (Fig. 1). Birds were sampled at three sites: Ré Island (n = 3), Yves Bay (n = 3) and Marennes-Oléron Bay (n = 9).

Pertuis Charentais study site on the French Atlantic coast with the sampling stations: Ré Island, Yves Bay, and Marennes-Oléron Bay
When it was possible during the dissection, sex and age class (juvenile/adult) were determined for each individual. The liver, kidneys, pectoral muscle and ventral feathers were sampled, weighed (wet weight, ww), placed in individual plastic bags and stored at −20 °C. Liver, kidneys and muscle samples were later freeze-dried and weighed again (dry weight, dw). Freeze-dried tissues were then ground and stored in individual plastic vials until further trace element, isotopic and MT analyses. Ventral feathers were washed to remove oil and dirt in a chloroform–methanol solution (2:1) in an ultrasonic bath for 2 min. Afterwards, they were rinsed in two consecutive pure methanol baths for a few seconds and dried at 40 °C for 48 h prior to being used for trace element and stable isotope analyses.
In order to specifically investigate mRNA expressions in the liver, kidneys and muscle, and enzymatic activities in the liver, four Red Knots accidentally dead in nets were collected at Marennes-Oléron in February 2011. For these analyses, the liver, kidneys and muscle were immediately dissected in the field after their deaths and divided into two parts. The first part was immediately frozen in liquid nitrogen and stored at −80 °C for enzymatic and genetic analyses. The second part was stored at −20 °C and subjected to the same treatment as previously described for trace element, stable isotope and MT determinations.
Trace element determinations
Total Hg analyses were carried out in Red Knots by atomic absorption spectrophotometry with an Advanced Mercury Analyser (ALTEC AMA 254) on dried tissue aliquots (liver, kidneys, pectoral muscle, ventral feathers) ranging from 4 to 50 mg (±0.01 mg) depending on the Hg concentration in the tissue following the protocol described in Bustamante et al. (2006). Mercury analyses were run under a thorough quality controlled program that includes analysis of Certified Reference Material (CRM; lobster hepatopancreas TORT-2; National Research Council, Canada). CRM aliquots were treated and analysed according to the same conditions as the samples. The results were in good agreement with the certified values, with a mean recovery rate of 92 %. The detection limit was 5 ng Hg g−1 dw.
Ag, As, Cd, Co, Cr, Cu, Fe, Mn, Ni, Pb, Se and Zn were analysed using a Varian Vista-Pro ICP-OES and a Thermo Fisher Scientific XSeries II ICP-MS in the liver, kidneys, pectoral muscle and ventral feathers. Aliquots of the biological samples (30–300 mg) were digested with 6 mL 67–70 % HNO3 and 2 mL 34–37 % HCl (Fisher Scientific, trace element grade quality), with the exception of feathers (3 mL HNO3 and 1 mL HCl). Acidic digestion of the samples was carried out overnight at room temperature, then using a Milestone microwave (30 min with constantly increasing temperature up to 120 °C, then 15 min at this maximal temperature). Each sample was brought up to a volume of 50 mL (or 25 mL for feathers) with milli-Q water. Three control samples (two CRMs and one blank) treated and analysed the same way as the samples were included in each analytical batch. The CRMs were dogfish liver DOLT-4 (NRCC) and lobster hepatopancreas TORT-2 (NRCC). Quantification limits and mean recovery rates were, respectively, equal to 0.1 μg L−1 and 82 % for Ag, 1 μg L−1 and 98 % for As, 0.1 μg L−1 and 94 % for Cd, 0.1 μg L−1 and 99 % for Co, 0.1 μg L−1 and 102 % for Cr, 5 μg L−1 and 93 % for Cu, 20 μg L−1 and 86 % for Fe, 5 μg L−1 and 90 % for Mn, 0.2 μg L−1 and 101 % for Ni, 0.1 μg L−1 and 86 % for Pb, 0.5 μg L−1 and 109 % for Se, and 20 μg L−1 and 98 % for Zn. Trace element concentrations are expressed in μg g−1 dw. Values below the quantification limit were taken into account in calculating of the means by assigning them values one-half of the detection limit for the given element (e.g. a value <0.02 was reported as 0.01 μg g−1 dw).
Nitrogen and carbon stable isotope analysis
Cleaned feathers of Red Knots were chopped using surgical scissors and accurately weighed (±0.001 mg) to a range between 0.200 and 0.500 mg. Liver and muscle samples were also weighed with the same accuracy and in the same range of masses. All samples were placed in tin capsules for carbon and nitrogen stable isotope analysis and analysed using an elemental analyser (Flash EA 1112 fitted with a “No Blank” option, Thermo Scientific, Milan, Italy) coupled to an isotope ratio mass spectrometer (Delta V Advantage, Conflo IV interface, Smart EA option, Thermo Scientific, Bremen, Germany). The results are reported in δ unit notation (expressed in per mil relative to standards: Vienna Pee Dee Belemnite for δ13C and N2 in air for δ15N). The analytical precision of the measurements was <0.06 and <0.1 ‰ for carbon and nitrogen, respectively.
Metallothionein determination
Approximately 100 mg of each freeze-dried liver aliquot was homogenized on ice in 6 mL 100 mM Tris buffer with β-mercaptoethanol at pH 8.1 and then centrifuged (30,000×g; 30 min; 4 °C). Before determining the MT level, the supernatant aliquot was submitted to heat denaturation (95 °C; 15 min), placed on ice for 10 min and centrifuged (10,000×g; 10 min, 4 °C) to separate heat-stable proteins from denatured proteins. The supernatant from the second centrifugation was then frozen (−20 °C) until MT quantification.
Red Knots were analysed for their hepatic MT levels. Differential pulse polarographic analysis (DPP) was used to determine the level of MT in the heat-denatured soluble fraction. DPP is a technique based on –SH compound determination according to the Brdička reaction (Brdička 1933) as described by Thompson and Cosson (1984). A Model 303A static Hg drop electrode was used. Certified rabbit liver MT (Sigma Chemical Co., St. Louis, MO) were used to carry out the calibration according to the method of standard additions. The system consisted of a bevelled capillary, an Hg working electrode, a platinum counter electrode and an Ag/AgCl reference electrode. Results are expressed in μg of MT per g of dry homogenized tissue.
Sequencing of genes
Five DNA fragments of the Red Knot were sequenced for β-actin (act), acetyl-CoA carboxylase (acc), Cu/Zn superoxide dismutase (sod1), metallothionein (mt) and NADP-dependent malic enzyme (me).
A quantity of 40 mg of liver was homogenized for extraction of total RNA using the Absolutely Total RNA Miniprep kit (Agilent Technologies, USA) according to the manufacturer’s instructions. All RNA extracted was of good quality after evaluation by electrophoresis on a 1 % agarose-formaldehyde gel, and the concentration was determined by spectrophotometry. First-strand cDNA was synthesized from 5 μg of previously extracted total RNA with the AffinityScript Multiple Temperature cDNA Synthesis kit (Agilent Technologies, USA) according to manufacturer’s instructions.
The non-specific primers used to obtain amplified cDNA fragments of the different genes by PCR were determined after multiple sequence alignments of birds or mammalian species using Clustal W software (Infobiogen), or were qPCR primers based on duck Cairina moschata and Anas platyrhynchos (Supplementary material Table S1). Amplified products were cloned into pGEM-T vector (Promega) and sequenced (Millegen, France).
For Mn superoxide dismutase (sod2), catalase (cat) and cytochrome c oxidase subunit 1 (cox1) primer pairs from C. moschata and A. platyrhynchos were used to amplify products less than 200 bp. Cloning and sequencing demonstrated that the primer pairs chosen from conserved regions of the C. moschata and A. platyrhynchos genes could be used for C. canutus for qPCR analysis (Table Supplementary material Table S2).
Quantitative real-time PCR
The mRNA expression levels of eight genes were investigated in the liver, kidneys and muscle of Red Knots by quantitative real-time PCR: β-actin, cytochrome c oxidase subunit 1, acetyl-CoA carboxylase, Cu/Zn superoxide dismutase, Mn superoxide dismutase, catalase, metallothionein, and NADP-dependent malic enzyme. Quantitative real-time PCR reactions were performed in an Mx3005P QPCR Systems (Agilent Technologies, USA) following the manufacturer’s instructions (one cycle at 95 °C for 10 min followed by 40 amplification cycles at 95 °C for 30 s, 60 °C for 30 s and 72 °C for 30 s). Each 25 μL reaction contained 1 μL of reverse transcribed product template, 12.5 μL of Brilliant master mix including the SyberGreen I fluorescent dye (Agilent Technologies, USA), enabling monitoring of the PCR amplification, and the gene-specific primer pair at a final concentration of 200 nM for each primer. Gene-specific primer pairs were determined using the LightCycler probe design software (version 1.0, Roche) (Supplementary material Table S2).
Reaction specificity was determined for each reaction from the dissociation curve of the PCR product. This dissociation curve was obtained by monitoring the SyberGreen fluorescence level during gradual heating of the PCR products from 60 to 95 °C. Relative gene expression levels were normalized according to β-actin gene expression which was stable across tissues and samples. Induction and repression factors were calculated between the Red Knot and the Black-tailed Godwit (Lucia et al. 2012) following the formula:
2(−ΔCT BQN)/2(−ΔCT RK), where ΔCT is the Threshold Cycle of the Black-tailed Godwit (BTG) and the Red Knot (RK) normalized according to their respective β-actin CT.
Antioxidant enzymes and lipid peroxidation
Glutathione peroxidase (GPx) activity was determined in the liver of Red Knots according to the method of Paglia and Valentine (1967) using a GPx assay kit (RS504/RS505, RANDOX, France) as previously described (Lucia et al. 2012).
Hepatic superoxide dismutase (SOD) and catalase activities were determined according to the methods of Wooliams et al. (1983) and Deisseroth and Dounce (1970), respectively, using assay kits (SD125, RANDOX, France and CAT100, Sigma-Aldrich, USA) as previously described (Lucia et al. 2012). The results are presented in units of SOD or catalase per mg of protein.
All activities were expressed in relation to protein concentration, measured according to the Bradford method with slight modifications, using bicinchoninic acid and 4 % copper sulfate (Smith et al. 1985). Serum albumin was used as a standard (Sigma-Aldrich, France).
Lipid peroxidation levels were assessed in liver according to MDA content, determined using a commercial MDA assay kit (Oxis International, USA), following the manufacturer’s instructions. The results are presented in nmol MDA per g of tissue. Antioxidant enzyme activity and lipid peroxidation were measured in UV microplates (Greiner Bio One, Germany) using a spectrophotometer (SAFAS Flx-Xenius, Monaco).
Statistical analysis
As normality and homogeneity of variance were not achieved despite log10(x + 1) transformation (Cochran C test), non-parametric analysis of variance was applied to assess differences in trace element concentrations between tissues (liver, kidneys, muscle and feathers), sexes or age classes in the Red Knot (Kruskal–Wallis and Mann–Whitney U test, Statistica 7.1). Non-parametric analyses of variance (Mann–Withney U test) were also employed to study differences in MT concentration, mRNA expression, enzymatic activities and trace element concentrations between the Red Knot (current study) and the Black-tailed Godwit (Lucia et al. 2012). Moreover, the Spearman test was applied to all correlations achieved in this study.
Distance-based redundancy analysis (dbRDA) was used to analyse the relations between trace element concentrations measured in Red Knots and Godwits and mRNA expressions, enzyme activities and MT protein concentrations. Trace element concentrations as well as mRNA expressions, enzyme activities and MT protein concentrations were first standardised according to the maximum value measured for each of these variables. Standardised trace elements were then used to produce a distance matrix among individual birds using Euclidean distance. This distance matrix was used to produce an ordination of individual birds constrained by the standardised values for mRNA expressions, enzyme activities and MT protein concentrations. Two separate analyses were performed on (1) liver data and (2) kidneys data. These analyses were performed using the PERMANOVA+ package (Anderson et al. 2008) from PRIMER v6 (Clarke and Gorley 2006).
Results
Trace element bioaccumulation
The mean concentrations and ranges of the 13 trace elements in four tissues of 15 Red Knots sampled in the Pertuis Charentais are presented in Table 1. Although concentrations were usually low, especially for Ag, Co, Cu, Cr, Hg, Mn, Ni, Pb, and Zn, elevated concentrations were observed for As, Cd and Se. Moreover, important variations in individual concentrations were observed for the majority of the elements. For example, two birds exceeded Cd concentrations of 10.0 μg g−1 in their kidneys while the others were below 2.0 μg g−1 dw. Hg concentrations appeared to be highest in feathers, with one individual reaching 4.5 μg g−1 dw (Kruskal–Wallis test, p < 0.05). For the essential elements, As and Se, 73 % of the individuals exceeded Se concentrations of 10.0 μg g−1 and 47 % exceeded 30.0 μg g−1 in the liver. Similarly, 80 % of the Red Knots exceeded As concentrations of 3.0 μg g−1 in the liver.
Many correlations were observed between elements in the four tissues studied. As and Se were positively correlated (liver: R = 0.71; kidneys: R = 0.89; muscle: R = 0.85; feathers: R = 0.66), as well as Cu and Zn (liver: R = 0.66; kidneys: R = 0.74; muscle: R = 0.90). As for the non-essential elements, Cd was significantly correlated with Zn in internal tissues (liver: R = 0.71; kidneys: R = 0.69; muscle: R = 0.62) and with Pb (muscle: R = 0.66; feathers: R = 0.66). Moreover, when Se:Hg molar ratios were calculated in the livers of Red Knots, the ratios always exceeded one for all the birds and ranged between 5 and 594.
Influence of individual characteristics (sex, age class, body weight)
No differences in trace element concentrations were observed according to the sex of the birds. In contrast, the results highlighted discrepancies in element concentrations between age classes, especially for Cd. Indeed, Cd concentrations in adult birds were significantly above concentrations in juveniles in muscle (0.057 ± 0.093 vs 0.010 ± 0.001 μg g−1 dw, respectively; Mann–Whitney U test, p = 0.025) and below those of juveniles in the feathers (0.010 ± 0.001 vs 0.025 ± 0.011 μg g−1 dw, respectively; p = 0.012).
Trace element concentrations demonstrated positive relationships with the body weight of the Red Knots. Indeed, in kidneys, Ag, As, Cd, Co, Cr, Cu, Hg, Mn, Ni, Pb, Se and Zn concentrations were all positively correlated with weight (correlation coefficients were between 0.48 for As and 0.83 for Pb). The same trend was observed in the feathers only for As (R = 0.53), Hg (R = 0.59), Pb (R = 0.50) and Se (R = 0.65).
Influence of trophic position and feeding habitat
The relationship between trophic position assessed through determination of δ15N and trace element concentrations in the Red Knots was investigated in three tissues: liver, muscle and feathers. Depending on the tissue, As, Hg and Se concentrations were positively correlated with δ15N (Spearman correlation test, all p < 0.05; Fig. 2).

δ15N versus As, Hg and Se concentrations in the liver  (As, Se) and feathers
(As, Se) and feathers  (Hg, Se) of Red Knots
(Hg, Se) of Red Knots
The relationship between the foraging habitats of birds, assessed through determination of δ13C, and their trace element concentrations was also studied in the three tissues. The only significant correlation was between Hg concentrations and δ13C in the liver and was negative (R = −0.82, Spearman correlation test, p < 0.05).
Trace element detoxification and effects: comparison with the Black-tailed Godwit
The trace element concentrations in three tissues (liver, kidneys, muscle) of the Red Knot and the Black-tailed Godwit are presented in Table 2. The Red Knot had significantly lower Ag, Cu, Fe and Zn concentrations in the liver, Co, Hg and Zn concentrations in the kidneys, and Zn concentrations in muscle compared with the Black-tailed Godwit. Conversely, Se concentrations were higher for the Red Knot in the three tissues, as well as As and Mn concentrations in kidneys and muscle, and Pb concentrations in the kidneys.
During the current study, a Red Knot partial β-actin cDNA of 887 bp was sequenced. The corresponding protein of 295 amino acids (aa) presented strong similarities with the Black-tailed Godwit β-actin (99 % identity). In the case of acetyl-CoA carboxylase (acc), the fragment sequenced (245 bp, 81 aa) corresponded to acc of the Greylag Goose (Anser anser; 100 % identity) and the Black-tailed Godwit (99 % identity). The partial Cu/Zn superoxide dismutase (sod1) cDNA sequenced (273 bp, 40 aa) presented strong similarities with Dunlin sod1 (Calidris alpina; 98 % identity) and the metallothionein (mt) cDNA sequenced (224 bp, 62 aa) closely matched those of the Black-tailed Godwit (98 % identity) and the Great Cormorant (Phalacrocorax carbo; 95 % identity) mt. The last gene sequenced, NADP-dependent malic enzyme (215 bp, 71 aa), presented high similarities with the malic enzyme of the Muscovy Duck (C. moschata; 100 % identity) and the Black-tailed Godwit (100 % identity).
The relative mRNA expression of seven genes either induced or repressed in the Red Knot compared to the Black-tailed Godwit and involved in detoxification processes, defence against oxidative stress, lipogenesis, or mitochondrial metabolism were investigated in the liver, kidneys and muscle (Table 3). The results demonstrated that cat gene expression in the Red Knot was 50-fold the level of Black-tailed Godwit cat expression in the liver and kidneys (induction factor: 50). In contrast, this gene appeared to be down-regulated in muscle of the Red Knot compared to the other species. The Red Knot also seemed to highly express the acc gene in the muscle. The only other gene that differed between the two species was the mt gene. Indeed, this gene appeared to be 3–7 times lower in the three tissues of the Red Knot compared to the Black-tailed Godwit (repression factor between 1/3 and 1/7; Table 3).
MT concentrations in the livers of Red Knots were on average 3,582 ± 1,656 μg g−1 dw (n = 15), showing a wide variability among individuals. Nonetheless, MT concentrations were positively correlated with concentrations of Cd (R = 0.69; Spearman test, p < 0.05), Co (R = 0.83), Cu (R = 0.86), Hg (R = 0.70), Mn (R = 0.79), Pb (R = 0.77) and Zn (R = 0.88). MT concentrations in the Red Knots were significantly below those in Godwits previously reported by Lucia et al. (2012) (7,615 ± 4,340 μg g−1 dw (n = 31); Mann–Withney U test, p = 0.002).
Results of enzymatic activity assays demonstrated that the Red Knot displayed higher hepatic catalase and SOD activities than the Black-tailed Godwit (Fig. 3). Conversely, this species showed higher GPx activity than the Red Knot. For lipid peroxidation, no discrepancies appeared between the two species. Enzyme activities were positively correlated with elemental concentrations. Indeed, catalase activity significantly increased with As and Se concentrations (R = 0.76 and 0.74, respectively; Spearman test, p < 0.05), whereas SOD activity was positively linked only with Se concentration only (R = 0.76). GPx activity appeared to be influenced by several elements. Indeed, this enzyme was correlated with Ag (R = 0.81), Cu (R = 0.83), Fe (R = 0.74) and Zn (R = 0.76) concentrations.

Malondialdehyde concentrations (nmol MDA per g of tissue), superoxide dismutase (units of SOD per mg of proteins), catalase (units of catalase per mg of proteins) and glutathione peroxidase activities (units of GPx per mg of proteins) in liver of Red Knots (RK, mean ± SD; n = 4) and Black-tailed Godwits (BTG, mean ± SD; n = 3). Significant differences between species are indicated by letters at the level α = 0.05 (Mann–Whitney U test)
A distance-based redundancy analysis (dbRDA) was performed on the liver and kidneys data (Fig. 4). The first-two axes extracted 80.1 and 80.7 % of the variance of the data clouds in the liver and kidneys, respectively. The first axis discriminated the two species. In the liver, the dbRDA demonstrated that the higher As and Se concentrations in Red Knots were linked with cat gene expression, as well as SOD and catalase enzyme activities (Fig. 4). In contrast, the higher Ag, Cu, Fe and Zn concentrations in Godwits were positively linked with mt gene expression, the concentration of MT proteins and GPx activity (Fig. 4). In the kidneys, higher As, Mn, Pb and Se concentrations were correlated with elevated expression of acc, cat, cox1, me, sod1 and sod2 mRNAs. In muscle, Spearman correlations demonstrated that acc gene expression was positively correlated with As (R = 0.82), Hg (R = 0.90), Mn (R = 0.92), and Se (R = 0.82) concentrations.

Distance-based redundancy analysis based on the trace elements concentrations in the liver (a, b) and kidneys (c, d) of the Red Knots (RK, n = 4) and the Black-tailed Godwit (BTG, n = 3). a–c Relation of the trace elements with the first two dimensions; b–d relation of the mRNA expressions and the enzyme activities with the first two dimensions. cox1 cytochrome c oxidase subunit 1, acc acetyl-CoA carboxylase, sod1 superoxide dismutase (Cu/Zn), sod2 mitochondrial superoxide dismutase (Mn), cat catalase, mt metallothionein, me NADP-dependent malic enzyme, CAT catalase enzyme activity, SOD superoxide dismutase enzyme activity, GPx glutathione peroxidase, MDA malondialdehyde
Discussion
Trace element distribution and toxicity significance
The current study, one of the first investigating trace elements in the Red Knot C. canutus, demonstrated generally low elemental concentrations in the investigated tissues. Nevertheless, several elements reached levels above potentially toxic thresholds, especially As and Se in the liver and kidneys as well as Cd in the kidneys. These results are in accordance with a previous study in the Southwest Atlantic coast of France where three individuals of this species were reported to have high As and Se concentrations (Lucia et al. 2010).
In birds, inorganic As is considered highly toxic in comparison with organic compounds of this element and may disrupt reproduction, act as an endocrine disruptor, trigger sub-lethal effects or even induce individuals’ death (Eisler 1994; Kunito et al. 2008). As concentrations in living organisms are generally low (<1 μg g−1 ww, approximately 5 μg g−1 dw; Braune and Noble 2009). 60 % of the birds were above this threshold. However, all the Red Knots were below 50 μg g−1 dw, a threshold associated with direct toxic effects in seabirds (Neff 1997). Even if As is mostly accumulated in its organic form (Neff 1997), many organic arsenicals undergo biotransformation and organisms could be exposed to toxic intermediates (Albert et al. 2008). Moreover, Red Knots prey on benthic macroinvertebrate (Quaintenne et al. 2010) which can accumulate inorganic forms of As including As(V) or As(III) (Baumann and Fisher 2011). Consequently, birds may be able to accumulate both inorganic and organic As that could trigger sub-lethal effects in the Red Knot. In the future, further investigations should be performed to assess the different forms of As accumulated by Red Knots in order to empirically determine whether birds can accumulate the more toxic forms.
The results highlighted discrepancies in Cd concentrations between age classes. Indeed, Cd concentrations of adult birds were significantly above concentrations of juveniles in muscle but below juveniles in feathers. Age is one of the main factors explaining Cd accumulation, as observed in waterfowl from southern Spain (Gómez et al. 2004). With continued exposure, even at low levels, Cd is accumulated throughout the life span of birds (Dailey et al. 2008). The lower Cd concentration in the feathers of adults compared to young birds could be a consequence of the balance between elemental uptake during the feather’s formation and elemental removal during the moult (Lewis and Furness 1991). Adults may therefore be able to deposit Cd in their plumage and subsequently eliminated it through moulting. Nonetheless in this study Cd mainly accumulated in the kidneys, which represent the major internal organ for accumulation of this non-essential element (Barjaktarovic et al. 2002; Gómez et al. 2004; Kojadinovic et al. 2007). The toxicity of this metal was previously demonstrated in vertebrates and birds in particular (Hughes et al. 2000). Although the current results were below concentrations observed in previous studies where adult Barau’s Petrel from the Reunion island were reported to have accumulated 66.8 ± 31.8 μg g−1 (Kojadinovic et al. 2007), low environmental levels such as 4.7 ± 0.7 μg g−1 in the liver of birds experimentally contaminated could trigger the appearance of sub-lethal effects (Lucia et al. 2009). Lucia et al. (2009) observed the appearance of oxidative stress and up-regulation of genes involved in mitochondrial metabolism. Cd is indeed able to inhibit the electron transfer chain and induce the production of reactive oxygen species (Wang et al. 2004; Bertin and Averbeck 2006). Similar endpoints could be expected in Red Knots given their exposure to Cd in the environment.
Although Se is essential to living organisms’ metabolism, this element is considered highly toxic depending upon the chemical form ingested (Stewart et al. 1999). Effects of Se include direct mortality, reproductive impairment with teratogenesis, growth reduction, histopathological lesions and alterations in hepatic glutathione metabolism (Hoffman 2002). Some authors have suggested an effect threshold of 30 μg g−1 dw in the liver of birds (Skorupa et al. 1996). Moreover, hepatic Se concentrations exceeding 10 μg g−1 dw are associated with lower reproductive success in breeding females as well as reduced adult weight gain (Outridge et al. 1999). In the current study, 73 % of Red Knots exceeded 10 μg g−1 dw and 47 % reached or were above 30 μg g−1 dw. Nevertheless, the co-accumulation of Se and Hg could reduce the toxic effects of both trace elements (Peterson et al. 2009). A Se:Hg molar ratio approaching one suggests the existence of mercuric selenide (HgSe) which is a well-known non-toxic form in marine mammals and birds (Koeman et al. 1973; Martoja and Berry 1980; Nigro and Leonzio 1996; Ikemoto et al. 2004). In the current study, Hg and Se concentrations were not correlated in internal tissues of Red Knots and molar ratios were always above one. Consequently, the excess hepatic Se and the high concentrations reached in each tissue strongly support a possible toxicity of this element for the Red Knot.
Relation between trace element concentrations, trophic level and feeding habitat
Stable isotopes of carbon and nitrogen are useful tools to study trophic relationships and prey origins to elucidate inter‐ and intra-specific dietary patterns, and determine if variations in elemental uptake could be explained by differences in foraging strategy (Jardine et al. 2006; Michener and Kaufman 2007). Depending on the tissue, As, Hg and Se were positively linked to the trophic level of birds as reflected by δ15N values. This result highlights the dietary specialization occurring in one particular species with individuals feeding on high trophic status prey and thus experiencing high As, Hg and Se exposure. Indeed, despite their trophic specialization (Piersma et al. 1998), Red Knots are able to feed on a variety of benthic macroinvertebrates that differ in profitability as well as digestive quality (van Gils et al. 2005). In their European wintering grounds, most of the diet consists of the Baltic clam Macoma balthica, the edible cockle Cerastoderma edule and the mudsnail Hydrobia ulvae (Quaintenne et al. 2010). These authors have demonstrated variations in the diet composition of Red Knots between sites but also within sites in Western Europe, corroborating the observed dietary specialization of individuals. The variation in δ15N signatures between individuals from the current study could partly come from the different life status and origin of the birds. However, when taken into account, these parameters triggered no statistical discrepancies in δ15N signatures. Nonetheless, this dietary specialization could explain the different trace element accumulations observed in Red Knots of the Pertuis Charentais.
Hepatic Hg concentrations were also negatively linked to δ13C, which is mainly used to indicate the foraging habitats of predators (Rubenstein and Hobson 2004). This result highlighted the fact that the more birds fed on continental prey impoverished in 13C, the more Hg concentrations increased. Inter-individual feeding behaviour discrepancies could partly be the origin of the important variations in trace element accumulation observed in free-ranging Red Knots.
Trace element detoxification and effects: comparison with the Black-tailed Godwit
This study was designed to investigate the detoxification of trace elements in shorebird species, which are vulnerable to high exposure when they feed on coastal prey during their migration. A previous study of the Black-tailed Godwit gave some insight about the possible effects triggered by a combination of contaminants in free-ranging species difficult to obtain from a fairly large number of individuals (Lucia et al. 2012). The objective was thus to compare the genetic and enzymatic responses of both Red Knots and Godwits.
The Red Knot and the Black-tailed Godwit were characterized by differential elemental bioaccumulation. The Red Knot had notably higher As and Se concentrations while the Black-tailed Godwit had higher hepatic Ag, Cu, Fe and Zn levels. This discrepancy resulted in differential gene expression, hepatic enzyme activities and MT concentrations between the two species. First, the cat gene was up-regulated in the liver and kidneys of Red Knots compared to Godwits. This pattern was also reflected in the catalase enzyme, whose activity was higher in the Red Knot. Correlation tests and redundancy analyses in both tissues demonstrated that catalase gene expression and enzyme activity were enhanced by higher As and Se concentrations. Bioaccumulation of both trace elements appeared to trigger oxidative stress in the Red Knot. Se especially has the property to react with thiols which generate reactive oxygen species (Spallholz and Hoffman 2002). Experimental studies with selenomethionine, an organic form of Se, have shown oxidative stress in wild aquatic birds (Hoffman 2002). Symptoms included alterations in hepatic glutathione metabolism towards a more oxidized state, accompanied by lipid peroxidation. Glutathione is an important antioxidant and a cofactor for antioxidant enzymes such as GPx. GSH depletion possibly triggered by Se could partly explain the reduced GPx activity observed in Red Knots in comparison to Black-tailed Godwits. Consequently, Red Knots may rely on other antioxidant defenses such as catalase or SOD whose activities were higher in this species.
Secondly, synthesis of MT is well known to be induced by several metals such as Ag, Cd, Cu, Hg and Zn (Scheuhammer 1987; Kägi 1991; Elliott et al. 1992). The synthesis of MT diverged between the two shorebird species. The gene encoding for MT was indeed down-regulated in the Red Knot in comparison to the Black-tailed Godwit. This down-regulation was also reflected on the MT protein concentration in Red Knots, which was two-fold lower. The higher hepatic Ag, Cu, Fe and Zn concentrations in Black-tailed Godwits might explain this difference. Indeed, mRNA expression of mt was positively influenced by these elements in the liver. Godwits therefore appeared to mainly use this detoxification system to counteract the toxicity of trace elements.
The use of sensitive biomarkers, such as mRNA expression, demonstrated that trace elements may trigger species-dependent effects and induce different detoxification pathways in two free-ranging shorebird species feeding in the same habitats during winter migration. In the case of the Red Knot, antioxidant enzymes such as catalase and SOD seemed to be the main protective mechanisms against oxidative stress possibly triggered by As and Se levels. Conversely, the Black-tailed Godwit strongly stimulated the production of MT and had higher GPx activity. Although both species used different detoxification systems, their MDA concentrations were similar. These different response pathways could therefore give the birds similar levels of protection against lipid peroxidation and likely trace element toxicity.
This study highlighted for the first time that two relatively closely related shorebird species (Scolopacidae) exploiting the same feeding habitats in the Pertuis Charentais during migration stopover and/or wintering, and subject to several contaminants in the natural environment, may develop divergent adaptive responses. Red Knots seemed to suffer from stronger oxidative stress than Godwits, probably triggered by higher As and Se concentrations. Nevertheless, our results tend to demonstrate that energy expenditure and the general health of shorebirds seem not to suffer strongly from trace element bioaccumulation. These results suggest that trace element effects and detoxification pathways are both species-dependent and trace element specific.
References
Adams SM, Giesy JP, Tremblay LA, Eason CT (2001) The use of biomarkers in ecological risk assessment: recommendations from the Christchurch conference on biomarkers in ecotoxicology. Biomarkers 6:1–6
Albert CA, Williams TD, Morrissey CA, Lai VWM, Cullen WR, Elliott JE (2008) Dose-dependent uptake, elimination, and toxicity of monosodium methanearsonate in adult zebra finches (Taeniopygia guttata). Environ Toxicol Chem 27:605–611
Anderson MJ, Gorley RN, Clarke KR (2008) PERMANOVA+ for PRIMER: guide to software and statistical methods. PRIMER-E Ltd, Plymouth
Barjaktarovic L, Elliott JE, Scheuhammer AM (2002) Metal and metallothionein concentrations in Scoter (Melanitta spp.) from the Pacific Northwest of Canada, 1989–1994. Arch Environ Contam Toxicol 43:486–491
Baumann Z, Fisher NS (2011) Modeling metal bioaccumulation in a deposit-feeding polychaete from labile sediment fractions and from pore water. Sci Total Environ 409:2607–2615
Berglund ÅMM, Sturve J, Förlin L, Nyholm NEI (2007) Oxidative stress in pied flycatcher (Ficedula hypoleuca) nestlings from metal contaminated environments in northern Sweden. Environ Res 105:330–339
Bertin G, Averbeck D (2006) Cadmium: cellular effects, modifications of biomolecules, modulation of DNA repair and genotoxic consequences (a review). Biochimie 88:1549–1559
Bocher P, Quaintenne G, Robin F, Doumeret A, Delaporte P (2012) Origins and age structure of Red Knot Calidris canutus staging and wintering on the Atlantic coast of France. J Ornithol 153:103–114
Braune BM, Noble DG (2009) Environmental contaminants in Canadian shorebirds. Environ Monit Assess 148:185–204
Brdička AR (1933) Polarographic studies with the dropping mercury method. A new test for proteins in the presence of cobalt salts in ammoniacal solution of ammonium chloride. Collect Czech Chem Commun 7:112–128
Bustamante P, Miramand P (2004) Interspecific and geographical variations of trace element concentrations in Pectinidae from European waters. Chemosphere 57:1355–1362
Bustamante P, Miramand P (2005) Subcellular and body distributions of 17 trace elements in the variegated scallop Chlamys varia from the Charente-Maritime coast (Bay of Biscay, France). Sci Total Environ 337:59–79
Bustamante P, Lahaye V, Durnez C, Churlaud C, Caurant F (2006) Total and organic Hg concentrations in cephalopods from the North Eastern Atlantic waters: influence of geographical origin and feeding ecology. Sci Total Environ 368:585–596
Clarke KR, Gorley RN (2006) PRIMER v6: user manual/tutorial. PRIMER-E Ltd, Plymouth
Dailey RN, Raisbeck MF, Siemion RS, Cornish TE (2008) Liver metal concentrations in Greater Sage-grouse (Centrocercus urophasianus). J Wildl Dis 44:494–498
Deisseroth A, Dounce AL (1970) Catalase: physical and chemical properties, mechanisms of catalysis, and physiological role. Physiol Rev 50:319–375
Delany S, Scott D (2006) Waterbird population estimates, 4th edn. Wetlands International, Wageningen
Eisler RA (1994) A review of arsenic hazards to plants and animals with emphasis on fishery and wildlife resources. In: Nriagu JO (ed) Arsenic in the environment, part II: human health and ecosystem effects. Wiley, New York, pp 185–259
Elliott JE, Scheuhammer AM, Leighton FA, Pearce PA (1992) Heavy metal and metallothionein concentrations in Atlantic Canadian seabirds. Arch Environ Contam Toxicol 22:63–73
Gómez G, Baos R, Gómara B, Jiménez B, Benito V, Montoro R, Hiraldo F, González MJ (2004) Influence of a mine tailing accident near Doñana National Park (Spain) on heavy metals and arsenic accumulation in 14 species of waterfowl (1998 to 2000). Arch Environ Contam Toxicol 47:521–529
Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, D’Agrosa C, Bruno JF, Casey KS, Ebert C, Fox HE, Fujita R, Heinemann D, Lenihan HS, Madin EMP, Perry MT, Selig ER, Spalding M, Steneck R, Watson R (2008) A global map of human impact on marine ecosystems. Science 319:948–952
Hoffman DJ (2002) Role of selenium toxicity and oxidative stress in aquatic birds. Aquat Toxicol 57:11–26
Hoffman DJ, Heinz GH, Sileo L, Audet DJ, Campbell JK, LeCaptain LJ, Obrecht HH III (2000) Developmental toxicity of lead-contaminated sediment in Canada geese (Branta Canadensis). J Toxicol Environ Health A 59:235–252
Hughes MR, Smits JE, Elliott JE, Bennett DC (2000) Morphological and pathological effects of cadmium ingestion on Pekin ducks exposed to saline. J Toxicol Environ Health A 61:591–608
Ikemoto T, Kunito T, Tanaka H, Baba N, Miyazaki N, Tanabe S (2004) Detoxification mechanism of heavy metals in marine mammals and seabirds: interaction of selenium with mercury, silver, copper, zinc, and cadmium in liver. Arch Environ Contam Toxicol 47:402–413
Jardine TD, Kidd KA, Fisk AT (2006) Applications, considerations, and sources of uncertainty when using stable isotope analysis in ecotoxicology. Environ Sci Technol 40:7501–7511
Kägi JH (1991) Overview of metallothionein. Methods Enzymol 205:613–626
Koeman JH, Peeters VHM, Koudstaal-Hol CHM, Tijoe PS, De Goeu JJM (1973) Mercury selenium correlation in marine mammals. Nature 246:385–386
Kojadinovic J, Cosson RP, Le Corre M, Bustamante P (2007) Trace elements in three marine birds breeding on Reunion Island (Western Indian Ocean). Part 1: factors influencing their bioaccumulation. Arch Environ Contam Toxicol 52:418–430
Koopman K (2002) Mass, moult, migration and subspecific status of red knots Calidris canutus on the Frisian Wadden Sea coast, the Netherlands. Wader Study Gr Bull 97:30–35
Kunito T, Kubota R, Fujihara J, Agusa T, Tanabe S (2008) Arsenic in marine mammals, seabirds, and sea turtles. Rev Environ Contam Toxicol 195:31–69
Lewis SA, Furness RW (1991) Mercury accumulation and excretion by laboratory reared black-headed gull (Larus ridibundus) chicks. Arch Environ Contam Toxicol 21:316–320
Lucia M, André JM, Gonzalez P, Baudrimont M, Gontier K, Maury-Brachet R, Davail S (2009) Impact of cadmium on aquatic bird Cairina moschata. Biometals 22:843–853
Lucia M, André JM, Gontier K, Diot N, Veiga J, Davail S (2010) Trace elements concentrations (mercury, cadmium, copper, zinc, lead, aluminium, nickel, arsenic, and selenium) in some aquatic birds of the Southwest Atlantic coast of France. Arch Environ Contam Toxicol 58:844–853
Lucia M, Bocher P, Cosson RP, Churlaud C, Robin F, Bustamante P (2012) Insight on trace element detoxification in the Black-tailed Godwit (Limosa limosa) through genetic, enzymatic and metallothionein analyses. Sci Total Environ 423:73–83
Mahéo R (2011) Limicoles séjournant en France (Littoral), 34ème contribution française aux dénombrements internationaux des oiseaux d’eau organisés par Wetlands International. Office national de la chasse et de la faune Sauvage, France
Martoja R, Berry JP (1980) Identification of tiemannite as a probable product of demethylation of mercury by selenium in cetaceans. A complement to the scheme of the biological cycle of mercury. Vie Milieu 30:7–10
Mateo R, Beyer WN, Spann JW, Hoffman DJ, Ramis A (2003) Relationships between oxidative stress, pathology and behavioral signs of lead poisoning in mallards. J Toxicol Environ Health A 66:1371–1389
Michener RH, Kaufman L (2007) Stable isotope ratios as tracers in marine food webs: an update. In: Michener RH, Lajtha K (eds) Stable isotopes in ecology and environmental science. Blackwell, Oxford, pp 238–282
Nebel S, Piersma T, van Gils JA, Dekinga A, Spaans B (2000) Length of stopover, fuel storage and sex-bias in the occurrence of red knots Calidris c. canutus and C. c. islandica in the Wadden Sea during southward migration. Ardea 88:165–176
Neff JM (1997) Ecotoxicology of arsenic in the marine environment. Environ Toxicol Chem 16:917–927
Nigro M, Leonzio C (1996) Intracellular storage of mercury and selenium in different marine vertebrates. Mar Ecol Prog Ser 135:137–143
Outridge PM, Scheuhammer AM, Fox GA, Braune BM, White LM, Gregorich LJ (1999) An assessment of the potential hazards of environmental selenium for Canadian wildlife. Environ Rev 7:81–96
Paglia DE, Valentine WN (1967) Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J Lab Clin Med 70:158–169
Peterson SA, Ralston NVC, Peck DV, Van Sickle J, Robertson JD, Spate VL, Morris JS (2009) How might selenium moderate the toxic effects of mercury in stream fish of the Western U.S.? Environ Sci Technol 43:3919–3925
Piersma T, Hoekstra R, Dekinga A, Koolhaas A, Wolf P, Battley P, Wiersma P (1993) Scale and intensity of intertidal habitat use by knots Calidris canutus in the Western Wadden Sea in relation to food, friends and foes. Neth J Sea Res 31:331–357
Piersma T, Van Aelst R, Kurk K, Berkhoudt H, Maas LRM (1998) A new pressure sensory mechanism for prey detection in birds: The use of principles of seabed dynamics? Proc R Soc B 265:1377–1383
Pigeot J, Miramand P, Guyot T, Sauriau PG, Fichet D, Le Moine O, Huet V (2006) Cadmium pathways in an exploited intertidal ecosystem with chronic cadmium inputs (Marennes-Oléron, Atlantic coast, France). Mar Ecol Prog Ser 307:101–114
Quaintenne G, van Gils JA, Bocher P, Dekinga A, Piersma T (2010) Diet selection in a molluscivore shorebird across Western Europe: Does it show short- or long-term intake rate-maximization? J Anim Ecol 79:53–62
Rubenstein DR, Hobson KA (2004) From birds to butterflies: animal movement patterns and stable isotopes. Trends Ecol Evol 19:256–263
Scheuhammer AM (1987) The chronic toxicity of aluminium, cadmium, mercury, and lead in birds: a review. Environ Pollut 46:263–295
Skorupa JP, Morman SP, Sefchick-Edwards JS (1996) Guidelines for interpreting selenium exposure of biota associated with nonmarine aquatic habitats. U.S. Fish and Wildlife Service, National Irrigation Water Quality Program, Sacramento
Smith P, Khron R, Hermanson G, Mallia A, Gartner F, Provanzano M, Fujimoto E, Goeke N, Olson B, Klenk D (1985) Measurement of protein using bicinchoninic acid. Anal Biochem 150:76–85
Spaans B, Van Kooten L, Cremer J, Leyrer J, Piersma T (2011) Densities of individually marked migrants away from the marking site to estimate population sizes: a test with three wader populations. Bird Study 58:130–140
Spallholz JE, Hoffman DJ (2002) Selenium toxicity: cause and effects in aquatic birds. Aquat Toxicol 57:27–37
Stewart MS, Spallholz JE, Neldner KH, Pence BC (1999) Selenium compounds have disparate abilities to impose oxidative stress and induce apoptosis. Free Radic Biol Med 26:42–48
Stohs SJ, Bagchi D (1995) Oxidative mechanisms in the toxicity of metal ions. Free Radic Biol Med 18:321–336
Stroud DA, Baker A, Blanco DE, Davidson NC, Delany S, Ganter B, Gill R, González P, Haanstra L, Morrison RIG, Piersma T, Scott DA, Thorup O, West R, Wilson J, Zöckler C (2006) The conservation and population status of the world’s waders at the turn of the millennium. In: Boere GC, Galbraith CA, Stroud DA (eds) Waterbirds around the world. The Stationery Office, Edinburgh, pp 643–648
Thompson JAJ, Cosson RP (1984) An improved electrochemical method for the quantification of metallothionein in marine organism. Mar Environ Res 11:137–152
van Gils JA, de Rooij SR, van Belle J, van der Meer J, Dekinga A, Piersma T, Drent R (2005) Digestive bottleneck affects foraging decisions in red knots Calidris canutus. I. Prey choice. J Anim Ecol 74:105–119
van Gils JA, Piersma T, Dekinga A, Spaans B, Kraan C (2006) Shellfish dredging pushes a flexible avian top predator out of a marine protected area. PLoS Biol 4:2399–2404
Vitousek PM, Mooney HA, Lubchenco J, Melillo JM (1997) Human domination of earth’s ecosystems. Science 277:494–499
Wang Y, Fang J, Leonard SS, Krishna Rao KM (2004) Cadmium inhibits the electron transfer chain and induces reactive oxygen species. Free Radic Biol Med 36:1434–1443
Wooliams JA, Wiener G, Anderson PH, McMurray CH (1983) Variation in the activities of glutathione peroxidase and superoxide dismutase and in the concentration of copper in the blood in various breed crosses of sheep. Res Vet Sci 34:253–256
Acknowledgments
The authors wish to thank the University of La Rochelle for its financial support through a post-doctoral grant to ML, as well as the Natural Reserves of the Pertuis Charentais (Moëze-Oléron, Marais d’Yves, Lilleau des Niges) for their technical assistance during bird catches. We especially thank P. Delaporte, P. Rousseau, J. Gautier and J. Gonin from the Moëze-Oléron Natural Reserve. We also thank P. Richard and G. Guillou (UMR LIENSs) for technical support during stable isotope analysis. This work was supported financially by the CPER (Contrat de Projet Etat-Région) and the CNRS.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Lucia, M., Bocher, P., Cosson, R.P. et al. Evidence of species-specific detoxification processes for trace elements in shorebirds. Ecotoxicology 21, 2349–2362 (2012). https://doi.org/10.1007/s10646-012-0991-3
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-012-0991-3