
Abstract
The woodlouse Porcellio dilatatus (Crustacea) is a suitable model organism as a biological indicator for the toxic effects evaluation induced by pollutants exposure in the environment. In this study, the influence of cadmium (Cd) species on survival and reproduction of isopods was investigated. Survival, growth and reproductive parameters (time to pregnancy, pregnancy duration, pregnancy and abortion occurrence, number of juveniles per female and juvenile weight) were recorded after isopods were exposed to two species of Cd deployed in food: Cd(Cys)2 and Cd(NO3)2. There was a difference between survival rates of exposed males and females to both Cd species but in the case of Cd(NO3)2 these differences were more accentuated, with females having higher survival rates. In the presence of both metal species a reduction in the number of pregnancies and pregnancy duration was observed, while in the case of Cd(Cys)2 all pregnancies were inconclusive. The number of juveniles delivered per female fed with Cd(NO3)2 contaminated food was lower than in the control, whereas the juvenile weights were higher. To the best of our knowledge, the present study is the first toxicity test demonstrating that metal speciation affects reproduction in isopods. Cd(Cys)2 showed to be the most toxic of the contaminants tested in this long term exposure and capable of jeopardizing the reproduction efforts of isopods.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Terrestrial isopods are saprophytic detritivores that inhabit the upper layer of the soil and surface leaf litter, while playing an important role in maintaining the structure and fertility of soils (Loureiro et al. 2006). Those invertebrate-mediated processes such as drainage, aeration, incorporation and degradation of organic matter are important in improving soil quality and energy flow through ecosystems (Hornung et al. 1998b; Odendaal and Reinecke 1999).
Isopods have been frequently used as model organisms in soil ecotoxicity tests as important representatives of the invertebrate soil fauna (Calhoa et al. 2006; Caseiro et al. 2000; Drobne et al. 2008; Engenheiro et al. 2005; Jemec et al. 2010; Ribeiro et al. 2001; Sousa et al. 1998; Stanek et al. 2006). They are also easily cultured under laboratory conditions, where they can complete their entire life-cycle (Caseiro et al. 2000), and protocols for toxicity testing are available (Hornung et al. 1998a, b) although no standard methods have been developed for these organisms (i.e. ISO, ASTM, OECD). Moreover they have been widely used for the examination of metal accumulation and toxicity testing, due to their extraordinary capacity to accumulate high metal concentrations from the environment, predominantly in their hepatopancreas (Donker et al. 1990; Drobne 1997; Farkas et al. 1996; Godet et al. 2011; Hames and Hopkin 1991; Hopkin 1990; Hornung et al. 1998b; Paoletti and Hassall 1999; Udovic et al. 2009).
The most widely used toxicological endpoints in isopod testing are survival, growth, food consumption and assimilation, moult frequency, reproduction, avoidance behaviour, enzymatic biomarkers and locomotory behaviour (Drobne 1997; Drobne et al. 2008; Drobne and Strus 1996; Engenheiro et al. 2005; Hornung et al. 1997; Jansch et al. 2005; Jemec et al. 2010; Loureiro 2005; Loureiro et al. 2006, 2009; Santos et al. 2010; Stanek et al. 2006; Vink and Kurniawati 1996). The use of reproduction patterns as response to toxicity is sometimes not convenient, as such tests are considered time-consuming and laborious. Reproduction is also difficult to assess, as females may retain the sperm for long periods of time following mating and before egg fertilization (Drobne 1997; Vink and Kurniawati 1996). On the other hand, reproduction as a response endpoint to test sublethal effects of chemicals in long-term exposures is crucial to understanding and transposing those effects to population and community levels. Similarly, evaluating changes in reproduction and survival patterns induced by stressors exposure is crucial to understanding population stability/growth and will reflect isopods’ role in decomposition processes and cycling of nutrients.
Cadmium has shown to be a critical pollutant in the environment as it is considered a priority substance for water and related matrices (soil, sediment, biota) in Directive 2008/105/EC. Soil contamination by Cd can have several sources like agriculture practices, by the use of contaminated water for irrigation, or mining and industry, including in urban soils aerial deposition of particulates from smelting activities, burning of fossil fuels or tyre wear (UK-EA 2002). In addition, previous dietary studies on the assimilation of Cd in the terrestrial isopod Porcellio dilatatus (Calhoa et al. 2006) have indicated that Cd speciation dictate the assimilation efficiency (AE) of Cd in plant-isopod food chain.
In this study, the effects of different Cd species on the survival and reproduction of P. dilatatus upon a long term exposure was investigated. The main goal of this study was to assess differences in toxicity of two Cd species [Cd(Cys)2 and Cd(NO3)2] on the survival and reproductive effort of isopods, using as parameters the time to reach pregnancy (as an indication of fertilization ability), pregnancy duration, pregnancy and/or inconclusive pregnancy, number of juveniles per female and juvenile weight. The use of Cd-cysteinate in these studies provides an experimental tool for exploring the bioavailability of Cd (Udovic et al. 2009) that is complexed within biological tissue. Therefore Cd-cysteinate represents the most elementary form (species) of thiol-bound Cd in biological systems. In previous studies (Calhoa et al. 2011), the AE for Cd was lower in isopods fed with food containing Cd(Cys)2, than for that containing Cd(NO3)2, demonstrating that the cysteine conjugate was less available.
Some vascular plants have mechanisms of tolerance that include the induction of metal-binding proteins such as phytochelatins and metallothioneins (Prasad 1995) which are small proteins with a significant concentration of cysteine (30%) (Ndayibagira et al. 2007). Cysteine contains a sulphydryl group that accounts for the Cd-metallothionein induction due to Cd high affinity for sulphur ligands (Roosens et al. 2005; Zalups and Ahmad 2003).
Materials and methods
Test organisms and culture conditions
Isopods were collected from cultures of P. dilatatus that have been maintained for more than 3 years under laboratory conditions and that have been derived from individuals collected in a secondary coastal dune system in Aveiro, Portugal. They were maintained on a substrate of sand in plastic containers at 20°C with a 16:8 h (light:dark) photoperiod. Oven-dried alder (Alnus glutinosa) leaves were provided ad libitum as a food source (Caseiro et al. 2000; Kautz et al. 2000) and distilled water was added for maintaining adequate moisture.
Lettuce and gelatine substrate
A mixture of lettuce (Lactuca sativa) leaves and gelatine was selected as a suitable food substrate to be used as the exposure route (Mann et al. 2005; Monteiro et al. 2008). Non-contaminated L. sativa leaves were reduced to powder using a mortar and pestle, and were mixed with a gelatine solution prepared from 2.5 g gelatine powder (VWR Prolabo, Fontenay Sous Bois, France) to which 12.5 ml deionised water (Milli-Q®) was added, and then mixed by vortexing (Wallace and Lopez 1996). Small portions of the mixture (gelatine discs) weighing approximately 9 mg (dry wt) were made by pipetting liquid gelatine onto Parafilm® (Pechiney Plastic Packaging, Menasha, WI, USA). These discs were stored frozen at −20 C while not in use (Wallace and Lopez 1997).
Three treatments (diets) were established based on previous studies (Calhoa et al. 2006, 2011) for this long term exposure test to evaluate the toxicity of metal speciation to the terrestrial isopod P. dilatatus:
-
Cd(Cys)2 contaminated food—gelatine contaminated with Cd-cysteinate mixed with non-contaminated leaves of L. sativa;
-
Cd(NO3)2 contaminated food—gelatine contaminated with Cd nitrate mixed with non-contaminated leaves of L. sativa.
-
Control food—gelatine mixed with non-contaminated leaves of L. sativa.
Gelatine discs were previously assayed for Cd radiospectrometry before being fed to isopods, and contained 335 ± 29 μg Cd/g dry wt for gelatine with Cd(NO3)2 and 744 ± 108 μg Cd/g dry wt (mean ± standard error) for gelatine with Cd(Cys)2.
Experimental setup
A total of 50 non-gravid females with similar weight (35–48 mg) were selected with both antennae and no phenotypic abnormalities and separated into a test box for 1 month to guarantee that they were not pregnant at the beginning of the test. Test boxes (polyethylene terephthalate (PET); ∅ 85 mm × 43 mm) were bottom covered with a thin layer of sand in order to provide similar conditions to those in the culture boxes. After this 1 month period, a total of 30 males were also selected. At this stage, a total of 30 females and 30 males (10 females and 10 males for each treatment previously described to guarantee Cd accumulation) were exposed individually in test boxes for a period of 28 days, and fed exclusively on gelatine discs. Hereafter, this period will be named individual exposure test. Survival was checked every 2 days.
Fungi growth may alter Cd bioavailability and thus gelatine disks were replaced every week, in order to prevent consumption of those disks that had become colonized with fungi.
After this 28 day period of individual exposure, one male and one female were paired randomly per box for mating, using 10 replicates per treatment. Hereafter, this test period will be referred as reproduction test. Female reproductive cycle and survival was monitored for 54 days, 3 times a week; when pregnancy was observed females were moved into a new box alone until they gave birth. These females were still exposed to the respective food treatments. The percentage of females that successfully reached pregnancy (i.e. successful egg fertilization), the time until pregnancy, duration of pregnancy and the percentage of inconclusive pregnancies (females that successfully reached pregnancy but were unable to carry it out until the end) were recorded. The number of juveniles born per female and their individual weight were also registered. Isopods were assayed for Cd body burden by radiospectrometry to obtain Cd accumulation.
Cadmium analysis
Individual isopods and gelatine discs (dried in an oven at 60°C, for 24 h) were placed in 3.5-ml Röhren tubes (Sarstedt, Newtown, NC, USA) and were analyzed for Cd109 by radiospectrometry in a Genesis Gamma1 bench-top gamma counter (Laboratory Technologies, USA). The Cd(NO3)2 and Cd(Cys)2 contamination solutions were analysed by inductively coupled plasma spectroscopy (ICPS) in a Jobin Ivon JY70 with a Meinard C001 nebuliser. Specific activities of the two contamination solutions were assessed by comparing gamma counts with measurements obtained by ICPS.
Statistical analysis
All data were checked for normality and homoscedasticity using SigmaStat (version 3.01, SPSS, Chicago, IL, USA). Statistical analysis was carried out at a significance level of 0.05, using t tests to compare pregnancy duration and juvenile weights or one-way analysis of variance (ANOVA) with Tukey and Dunnett’s test for multiple comparisons of means to determine differences on time to reach pregnancy and number of juveniles relatively to control treatment. Whenever possible, data that was not normally distributed or whose equal variance testing failed were transformed.
The lethal time of 50% (LT50) was calculated with the Probit method using the MiniTab software (2000).
Results
Survival
During the individual exposure period no mortality was observed (Fig. 1). During the 82 days of exposure, mortality did not occur among control females while 20% of the control males died during the test. This is in accordance to the 20% mortality value that is referred in Hornung et al. (1998a) as a validity criteria for controls in isopod testing. In the Cd(Cys)2 treatment 70% of females and 90% of males died at the end of the test, whereas for the Cd(NO3)2 treatment mortality was of 10 and 70%. LT50 values for Cd(Cys)2 exposures were respectively for females and males: 65.85 days (63.97–67.89 for 95%CL) and 58.35 days (56.80–59.89 for 95%CL). For the Cd(NO3)2 exposure, only data from male survival gave a LT50 value of 70.36 days (68.44–72.57 for 95%CL). Females exposed to Cd(NO3)2 showed a low mortality rate of 10% (Fig. 1).

Number of females (a) and males (b) of P. dilatatus that survived during the 82 days of the test
Cd accumulation in isopods
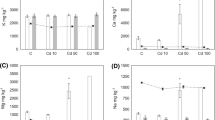
No cadmium was detected in any of the control isopods. Isopods exposed to Cd(NO3)2 and Cd(Cys)2 had equivalent rates of Cd accumulation (6.4 ± 1.9 and 7.8 ± 1.7 ng Cd/mg animal, respectively) after the individual exposure period of 28 days (Fig. 2). At the end of the experiment isopods fed with Cd(NO3)2 accumulated much more Cd (17.9 ± 2.3 ng Cd/mg animal) than those from the Cd(Cys)2 treatment (9.9 ± 2 ng Cd/mg animal).

Cadmium accumulation by P. dilatatus before the beginning of the experiment (T0), after 28 days of individual exposure (T1) and at the final of the experiment (82 days of exposure) (T2). Grey bars represent the accumulation in isopods fed with Cd(NO3)2 and the white bars represent the accumulation in isopods fed with Cd(Cys)2. Vertical error bars represent the standard error of the mean (n ranges from 4 to 12)
Time to reach pregnancy, pregnancy duration and abortions
During the individual exposure test, plus the 54 days of the reproduction test, all animals lost weight, with the exception of the only male that survived in the Cd(Cys)2 treatment, that showed a growth rate of 14.22%. There were no significant differences between treatments and within sexes (female data after exponential transformation, p = 0.117; males, p = 0.376).
As shown in Fig. 3, the average time at which the control isopods showed the first signs of pregnancy following mating was of 19 ± 2 days. This interval was narrower for isopods fed with Cd(NO3)2 (12 ± 1 days) and even more so for those in the Cd(Cys)2 treatment (10 ± 2 days). Statistically significant differences were found between the control and the two Cd treatments (p < 0.05). Average pregnancy duration was of 21 ± 1 days for the control, and decreased in Cd(NO3)2 treatment (14 ± 3 days), while for the Cd(Cys)2 treatment, and although signs of pregnancy were observed, no females were able to deliver mancae.

Number of days until first signs of pregnancy were detected (grey bars), and number of days between first signs of pregnancy and release of mancae (white bars) of P. dilatatus fed with Cd(Cys)2 and Cd(NO3)2 gelatine and lettuce discs. n ranges from 3 to 10. Vertical error bars represent the standard error of the mean. * Indicates a significant difference (p < 0.05) from the control (ANOVA, Dunnett’s test); ** indicates a significant difference (p < 0.05) between the control and Cd(NO3)2 treatments (t test)
In the control all females successfully reached pregnancy within the test period (Fig. 4). Only 30% of the females fed with Cd(Cys)2 became pregnant but none were able to take it through: two died and the one that survived, failed to produce offspring. In the Cd(NO3)2 treatment, half of the females successfully reached pregnancy but only 80% were able to reach term.

Percentage of successful females achieving pregnancy (grey bars) and percentage of female inconclusive pregnancies (white bars) of P. dilatatus feed with Cd(Cys)2 and Cd(NO3)2 gelatine and lettuce discs
Number of juveniles and individual juvenile weight
Significant differences were found between the number of juveniles in control and in Cd treatments (p < 0.001). Differences between the Cd(Cys)2 and the Cd(NO3)2 were not statistically significant, although no mancae were delivered in females exposed to Cd(Cys)2 (Fig. 5). In the control the average number of mancae delivered per female was 21 ± 2. In the Cd(Cys)2 none of the females were able to carry out the pregnancy and therefore, no juveniles were delivered. In the Cd(NO3)2, the number of mancae delivered per female was 7 ± 2.

Number of juveniles hatching per pregnant female (grey bars), and individual mancae weight (white bars) of P. dilatatus fed with Cd(Cys)2 and Cd(NO3)2 gelatine and lettuce discs. Vertical error bars represent the standard error of the mean. (a) and (b) Indicates a significant difference between number of juveniles produced in the treatments (ANOVA, Tukey test, p < 0.001); asterisk indicates a significant difference between juvenile individual weight (t test, p < 0.05)
The total juvenile weight was significantly different between the control and the Cd(NO3)2 treatment (ANOVA, Dunnett’s test, p < 0.001). In the control, there were more juveniles delivered and therefore the total weight (6.8 ± 0.31 mg) was higher (ANOVA, Dunnett’s test, p < 0.001) than in that exposed to Cd(NO3)2 (3.3 ± 0.96 mg). The decrease in the number of mancae per female in this later treatment lead to an individual mancae weight increase. There were significant differences between individual weight of the control juveniles and those in the Cd(NO3)2 treatment (ANOVA, Tukey test, p = 0.007). Although more juveniles were delivered in the control, their individual weight was lower (0.338 ± 0.015 mg) than of those exposed to Cd(NO3)2 (t test, p < 0.05), where fewer juveniles with higher individual weight (0.435 ± 0.029 mg) were produced.
Discussion
Besides being considered very toxic, Cd has no known biological function (Odendaal and Reinecke 1999). Effects on the environment have been mostly studied in the aquatic compartment, whereas in the terrestrial compartment most ecotoxicological tests have been performed using soluble Cd2+ salts (ECB 2007). Life-history theory predicts that habitat disturbances will increase mortality, select for early reproduction and increase reproductive effort (Donker et al. 1993). From the survival data, there was an unexpected differential response between sexes in both exposures. In several studies, differentiation between sexes is not considered (Calhoa et al. 2006; Jansch et al. 2005; Loureiro et al. 2009) due to the expected similarity on behaviour and survival in terrestrial isopods. The inexistence of differences between sexes in isopods was described in the study where the aquatic isopod Idotea baltica was exposed to zinc, copper and lead and no differences were observed between males and females’ LT50 values (Bat et al. 1999). In the present work, males exposed to Cd(Cys)2 died earlier than those exposed to Cd(NO3)2, showing higher acute toxicity as a response to long-term exposure.
Isopods fed with Cd(NO3)2 and Cd(Cys)2 had equivalent Cd accumulation (6.4 and 7.8 ng Cd/mg animal, respectively) after the 28 days of individual exposure. It was provided a higher concentration of Cd in Cd(Cys)2 gelatine discs to try to equal metal accumulation by isopods fed with both Cd species, because in previous experiments (Calhoa et al. 2011) isopods fed with Cd(Cys)2 accumulated less Cd than those exposed to Cd(NO3)2 contaminated food. It was important to have similar accumulation rates for a reproduction test in order to compare toxicity responses between both Cd species. But at the end of the experiment isopods fed with Cd(NO3)2 contaminated food accumulated higher Cd concentration (17.9 ng Cd/mg animal) compared to those that received the Cd(Cys)2 treatment (9.9 ng Cd/mg animal). Studies made by Vijver et al. (2006) on Cd accumulation on the isopod Porcellio scaber showed similar accumulation rates (5.77 mg Cd/kg dry weight) after 32 days of exposure to contaminated soil and food with 109Cd.
In a previous study, Calhoa et al. (2011) showed that isopods fed with Cd(Cys)2 gelatine discs had a higher storage level of Cd in the metal-sensitive fractions. It is hypothesized that Cd in organelles and in heat denatured proteins can be considered potentially vulnerable fractions, and in the present study it is demonstrated that when Cd is predominantly present in these fractions it will lead to increased toxicity. The Cd(Cys)2 treatment induced the highest mortality probably due to higher availability of the ingested Cd that impaired physiological processes. Isopods fed with Cd(NO3)2 stored Cd in metal-rich granules as a detoxification strategy so they were more efficient at detoxifying Cd, reflecting in an increased metal body burden, though being less toxic to the isopod (Calhoa et al. 2011). In this way, Cd within granules is not available for the physiological processes, thus becoming less toxic. This could be also related to metal tolerance and resistance that may be attributed to the ability of isopods to compartmentalize Cd in the hepatopancreas, acting as a detoxification mechanism and contributing to tolerance to high Cd levels.
Parental care by female isopods is a behavioural strategy that contributes to increase fitness of progeny but is energetically costly (Lardies et al. 2004). Terrestrial isopods have an extensive parental care since they carry the eggs and juveniles in a ventral marsupium, that is provisioned with fluid from the mother and allows early development to take place, independently of an external water source (Lardies et al. 2004). The tradeoff between energy losses with reproduction/maternal care and those from detoxification processes due to Cd exposure might justify a shorter isopod pregnancy, to allow shifting energy for detoxification.
Jones and Hopkin (1996) showed that terrestrial isopods (P. scaber and Oniscus asellus) in metal polluted sites had higher reproductive investment, suggesting that they were able to redirect resources from other functions, like growth, to meet the physiological costs of metal detoxification. Donker et al. (1993) also showed that isopods from a zinc smelter started to reproduce earlier and that metal contamination compromised adult survival, while the reproductive effort was higher when compared to isopods in reference sites. In our experiment, isopods were previously exposed to Cd for 28 days in order to evaluate the effects of Cd accumulation on both the isopod reproduction ability (fecundity) and on the egg fertilization phase.
During the 28 days of the individual exposure test plus the 54 days of the reproduction test all animals lost weight, probably due to the long duration of this test in which a long period of stress was involved. Growth inhibition among isopods is a commonly reported consequence of Cd exposure (Odendaal and Reinecke 2004), and in this study may reflect the increased cost of detoxification of Cd (Zidar et al. 2003). Considering the high rates of mortality observed in this study and although isopods exposed to Cd(NO3)2 accumulated more Cd, higher female mortalities were observed in the Cd(Cys)2 treatments, showing that Cd(Cys)2 induced higher acute toxicity.
In the control, all females became pregnant and delivered mancae within the 54 days of the reproduction test. These values exceed those in the few reports available (e.g. Hornung and Warburg 1994; van Brummelen et al. 1996) with other isopod species but are in accordance with Lemos et al. (2010) that reported the same percentage of successfully pregnancy (100%) for P. scaber females.
Control females showed the first signs of pregnancy later than that in the Cd treatments which is in accordance with life-history theory that predicts a selection for early reproduction and increase reproductive effort when stressed (Donker et al. 1993).
Pregnancy duration was higher for the control treatments and decreased in Cd(NO3)2 treatment where juveniles were released 7 days earlier (as mean time). As Cd induced a decrease on the brood period, females exposed to Cd(NO3)2 had a reduced length of time to provide for their developing young. A similar observation was recorded in females of Armadillidium vulgare exposed to predatory ants (Castillo and Kight 2005), as well as in females of Porcellio leavis under physical stress (5 min of continuous locomotion daily throughout the brooding period) (Kight and Nevo 2004), thus exhibiting significantly shorter brooding periods compared with the controls and releasing juveniles almost 48 h earlier.
The isopod marsupium physically limits the area available for egg attachment, and abnormal egg growth may lead to overcrowding and consequent egg loss or reabsorption, following-on fewer offspring (Lardies et al. 2004). This phenomenon may had occurred in the present experiment in regard to the Cd(NO3)2 treatment. The oocyte resorption takes place when the albumin is stopped from entering the oocyte and no vitellogenin synthesis takes place. This process leaves no visible traces after the resorption has been completed. In the Cd(Cys)2 none of the females were able to carry out a successful pregnancy (most animals died before breeding) and the only survivor did not deliver mancae. Hornung and Warburg (1994) reported for the first time the oosorption in an ovary of an oniscid isopod (P. ficulneus) and Farkas et al. (1996) showed that P. scaber under stressful conditions increased disruption of oocytes within the ovarium during vitellogenesis (oosorption), resulting in fewer juveniles.
Upon Cd(NO3)2 exposure it was observed a decrease in the number of mancae delivered per female with an increase in juveniles’ individual weight, which might be a response for their investment in quality rather than in quantity of juveniles. Among crustaceans, fecundity has been demonstrated to be particularly sensitive to dietary metal exposure (Hook and Fisher 2001; Mann and Hyne 2008; Mann et al. 2009). Hence we suggest that females in the Cd(NO3)2 treatment, balanced the benefits of having smaller clutch decreasing the costs of providing parental care, which implies less energy for maintenance.
Conclusion
Different metal species has been shown in previous studies to affect the AE and compartmentalization of Cd in isopods, strongly suggesting that Cd accumulates in the organism differently depending on the form it is provided in food. In this case the impairment of physiological processes by Cd in P. dilatatus induced a decrease in time to reach pregnancy, pregnancy duration, and also the occurrence of inconclusive pregnancies on the Cd(Cys)2 treatment.
Cd(Cys)2 showed to be more toxic than Cd(NO3)2 in long term exposure, demonstrating that different metal species affects differently female and male survival and that also impacts on the reproduction efforts in isopods.
The species P. dilatatus showed to be a good model test organism in this kind of studies where accumulation and possible trophic transfer are evaluated.
References
Bat L, Sezgín M, Gundogdu A, Çulha M (1999) Toxicity of zinc, copper and lead to Idotea baltica (Crustacea, Isopoda). Turk J Biol 23:465–472
Calhoa CF, Soares A, Mann RM (2006) Cadmium assimilation in the terrestrial isopod, Porcellio dilatatus—Is trophic transfer important? Sci Total Environ 371(1–3):206–213. doi:10.1016/j.scitotenv.2006.09.013
Calhoa CF, Monteiro MS, Soares AMMV, Mann RM (2011) The influence of metal speciation on the bioavailability and sub-cellular distribution of cadmium to the terrestrial isopod, Porcellio dilatatus. Chemosphere 83:531–537
Caseiro I, Sousa JP, Santos S, Nogueira AJA, Soares AMVM (2000) Optimization of culture conditions of Porcellio dilatatus (Crustacea: Isopoda) for laboratory test development. Ecotoxicol Environ Saf 47(3):285–291. doi:10.1006/eesa.2000.1982
Castillo ME, Kight SL (2005) Response of terrestrial isopods, Armadillidium vulgare and Porcellio laevis (Isopoda: Oniscidea) to the ant Tetramorium caespitum: morphology, behavior and reproductive success. Invertebr Reprod Dev 47(3):183–190
Donker MH, Koevoets P, Verkleij JAC, Vanstraalen NM (1990) Metal-binding compounds in the hepatopancreas and hemolymph of Porcellio scaber (Isopoda) from contaminated and reference areas. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol 97(1):119–126
Donker MH, Zonneveld C, Vanstraalen NM (1993) Early reproduction and increased reproductive allocation in metal-adapted populations of the terrestrial isopod Porcellio scaber. Oecologia 96(3):316–323
Drobne D (1997) Terrestrial isopods—a good choice for toxicity testing of pollutants in the terrestrial environment. Environ Toxicol Chem 16(6):1159–1164
Drobne D, Strus J (1996) Moult frequency of the isopod Porcellio scaber, as a measure of zinc-contaminated food. Environ Toxicol Chem 15(2):126–130
Drobne D, Blazic M, Van Gestel CAM, Leser V, Zidar P, Jemec A, Trebse P (2008) Toxicity of imidacloprid to the terrestrial isopod Porcellio scaber (Isopoda, Crustacea). Chemosphere 71(7):1326–1334
ECB (2007) European Union Risk Assessment Report: cadmium oxide and cadmium metal. Vol Part I—Environment Institute for Health and Consumer Protection. European Chemicals Bureau, Ispra
Engenheiro EL, Hankard PK, Sousa JP, Lemos MF, Weeks JM, Soares AMM (2005) Influence of dimethoate on acetylcholinesterase activity and locomotor function in terrestrial isopods. Environ Toxicol Chem 24(3):603–609
Farkas S, Hornung E, Fischer E (1996) Toxicity of copper to Porcellio scaber Latr (Isopoda) under different nutritional status. Bull Environ Contam Toxicol 57(4):582–588
Godet J-P, Demuynck S, Waterlot C, Lemière S, Souty-Grosset C, Scheifler R, Douay F, Leprêtre A, Pruvot C (2011) Growth and metal accumulation in Porcellio scaber exposed to poplar litter from Cd-, Pb-, and Zn-contaminated sites. Ecotoxicol Environ Saf 74(3):451–458
Hames CAC, Hopkin SP (1991) Assimilation and loss of Cd-109 and Zn-65 by the terrestrial isopods Oniscus asellus and Porcellio scaber. Bull Environ Contam Toxicol 47(3):440–447
Hook SE, Fisher NS (2001) Reproductive toxicity of metals in calanoid copepods. Mar Biol 138(6):1131–1140
Hopkin SP (1990) Species-specific differences in the net assimilation of zinc, cadmium, lead, copper and iron by the terrestrial isopods Oniscus asellus and Porcellio scaber. J Appl Ecol 27(2):460–474
Hornung E, Warburg MR (1994) Oosorption and oocyte loss in a terrestrial isopod under stressful conditions. Tissue Cell 26(2):277–284
Hornung E, Fischer E, Farkas S (1997) Isopod reproduction as a tool for sublethal-toxicity tests. In: 4th Symposium on the biology of terrestrial isopods, Haifa, Israel, 03–07 Feb 1997, pp 445–450
Hornung E, Farkas S, Fischer E (1998a) Tests on the isopod Porcellio scaber. In: Løkke H, Van Gestel CAM (eds) Handbook of soil invertebrate toxicity tests. Wiley, Chichester
Hornung E, Fischer E, Farkas S (1998b) Isopod reproduction as a tool for sublethal-toxicity tests. Isr J Zool 44(3–4):445–450
Jansch S, Garcia M, Rombke J (2005) Acute and chronic isopod testing using tropical Porcellionides pruinosus and three model pesticides. In: 6th International symposium on the biology of terrestrial isopods, Aveiro, Portugal, 12–15 Jul, pp 143–152. doi:10.1016/j.ejsobi.2005.09.010
Jemec A, Drobne D, Tisler T, Sepcic K (2010) Biochemical biomarkers in environmental studies-lessons learnt from enzymes catalase, glutathione S-transferase and cholinesterase in two crustacean species. Environ Sci Pollut Res 17(3):571–581. doi:10.1007/s11356-009-0112-x
Jones DT, Hopkin SP (1996) Reproductive allocation in the terrestrial isopods Porcellio scaber and Oniscus asellus in a metal-polluted environment. Funct Ecol 10(6):741–750
Kautz G, Zimmer M, Topp W (2000) Responses of the parthenogenetic isopod, Trichoniscus pusillus (Isopoda: Oniscidea), to changes in food quality. Pedobiologia 44(1):75–85
Kight SL, Nevo M (2004) Female terrestrial isopods, Porcellio laevis Latreille (Isopoda: Oniscidea) reduce brooding duration and fecundity in response to physical stress. J Kans Entomol Soc 77(3):285–287
Lardies MA, Cotoras IS, Bozinovic F (2004) The energetics of reproduction and parental care in the terrestrial isopod Porcellio laevis. J Insect Physiol 50(12):1127–1135
Lemos MFL, van Gestel CAM, Soares A (2010) Reproductive toxicity of the endocrine disrupters vinclozolin and bisphenol A in the terrestrial isopod Porcellio scaber (Latreille, 1804). Chemosphere 78(7):907–913. doi:10.1016/j.chemosphere.2009.10.063
Loureiro S (2005) Isopod biology—guest-editorial note. Eur J Soil Biol 41(3–4):55–56. doi:10.1016/j.ejsobi.2005.11.003
Loureiro S, Sampaio A, Brandao A, Nogueira AJA, Soares A (2006) Feeding behaviour of the terrestrial isopod Porcellionides pruinosus Brandt, 1833 (Crustacea, Isopoda) in response to changes in food quality and contamination. Sci Total Environ 369(1–3):119–128
Loureiro S, Amorim MJB, Campos B, Rodrigues SMG, Soares A (2009) Assessing joint toxicity of chemicals in Enchytraeus albidus (Enchytraeidae) and Porcellionides pruinosus (Isopoda) using avoidance behaviour as an endpoint. Environ Pollut 157(2):625–636. doi:10.1016/j.envpol.2008.08.010
Mann RM, Hyne RV (2008) Embryological development of the Australian amphipod, Melita plumulosa Zeidler, 1989 (Amphipoda, Gammaridea, Melitidae). Crustaceana 81(1):57–66
Mann RM, Matos P, Loureiro S, Soares AMV (2005) Foundation studies for cadmium accumulation studies in terrestrial isopods—diet selection and diet contamination. Eur J Soil Biol 41(3–4):153–161. doi:10.1016/j.ejsobi.2005.09.013
Mann RM, Hyne RV, Spadaro DA, Simpson SL (2009) Development and application of arapid amphipod reproduction test for sediment-quality assessment. Environ Toxicol Chem 28(6):1244–1254
Minitab Statistical Software (2000) Minitab, Ver 13.0. State College, PA, USA
Monteiro MS, Santos C, Soares A, Mann RM (2008) Does subcellular distribution in plants dictate the trophic bioavailability of cadmium to Porcellio dilatatus (Crustacea, Isopoda)? Environ Toxicol Chem 27(12):2548–2556
Ndayibagira A, Sunahara GI, Robidoux PY (2007) Rapid isocratic HPLC quantification of metallothionein-like proteins as biomarkers for cadmium exposure in the earthworm Eisenia andrei. Soil Biol Biochem 39(1):194–201
Odendaal JP, Reinecke AJ (1999) The toxicity of sublethal lead concentrations for the woodlouse, Porcellio laevis (Crustacea, Isopoda). Biol Fertil Soils 29(2):146–151
Odendaal JP, Reinecke AJ (2004) Effect of metal mixtures (Cd and Zn) on body weight in terrestrial isopods. Arch Environ Contam Toxicol 46(3):377–384. doi:10.1007/s00244-003-2303-7
Paoletti MG, Hassall M (1999) Woodlice (Isopoda : Oniscidea): their potential for assessing sustainability and use as bioindicators. Agric Ecosyst Environ 74(1–3):157–165
Prasad MNV (1995) Cadmium toxicity and tolerance in vascular plants. Environ Exp Bot 35(4):525–545
Ribeiro S, Sousa JP, Nogueira AJA, Soares A (2001) Effect of endosulfan and parathion on energy reserves and physiological parameters of the terrestrial isopod Porcellio dilatatus. Ecotoxicol Environ Saf 49(2):131–138
Roosens NH, Leplae R, Bernard C, Verbruggen N (2005) Variations in plant metallothioneins: the heavy metal hyperaccumulator Thlaspi caerulescens as a study case. Planta 222(4):716–729. doi:10.1007/s00425-005-0006-1
Santos M, Ferreira N, Soares A, Loureiro S (2010) Toxic effects of molluscicidal baits to the terrestrial isopod Porcellionides pruinosus (Brandt, 1833). J Soils Sediments 10(7):1335–1343. doi:10.1007/s11368-010-0246-y
Sousa JP, Vingada JV, Loureiro S, da Gama MM, Soares A (1998) Effects of introduced exotic tree species on growth, consumption and assimilation rates of the soil detritivore Porcellio dilatatus (Crustacea: Isopoda). Appl Soil Ecol 9(1–3):399–403
Stanek K, Drobne D, Trebse P (2006) Linkage of biomarkers along levels of biological complexity in juvenile and adult diazinon fed terrestrial isopod (Porcellio scaber, Isopoda, Crustacea). Chemosphere 64(10):1745–1752
Udovic M, Drobne D, Lestan D (2009) Bioaccumulation in Porcellio scaber (Crustacea, Isopoda) as a measure of the EDTA remediation efficiency of metal-polluted soil. Environ Pollut 157(10):2822–2829
UK-EA (2002) Soil guideline values for cadmium contamination. Department for Environment, Food and Rural Affairs. The UK Environment Agency, Bristol
van Brummelen TC, van Gestel CAM, Verweij RA (1996) Long-term toxicity of five polycyclic aromatic hydrocarbons for the terrestrial isopods Oniscus asellus and Porcellio scaber. Environ Toxicol Chem 15(7):1199–1210
Vijver MG, Vink JPM, Jager T, van Straalen NM, Wolterbeek HT, van Gestel CAM (2006) Kinetics of Zn and Cd accumulation in the isopod Porcellio scaber exposed to contaminated soil and/or food. Soil Biol Biochem 38(7):1554–1563
Vink K, Kurniawati TA (1996) Iteroparous reproduction, body growth and energy reserves in a tropical population of the isopod, Porcellionides pruinosus. Pedobiologia 40(5):467–476
Wallace WG, Lopez GR (1996) Relationship between subcellular distribution in prey and cadmium transfer to a predator cadmium trophic. Estuaries 19:923–930
Wallace WG, Lopez GR (1997) Bioavailability of biologically sequestered cadmium and the implications of metal detoxification. Mar Ecol Prog Ser 147(1–3):149–157
Zalups RK, Ahmad S (2003) Molecular handling of cadmium in transporting epithelia. Toxicol Appl Pharmacol 186(3):163–188. doi:10.1016/s0041-008x(02)00021-2
Zidar P, Drobne D, Strus J, Blejec A (2003) Intake and assimilation of zinc, copper, and cadmium in the terrestrial isopod Porcellio scaber Latr. (Crustacea, Isopoda). Bull Environ Contam Toxicol 70(5):1028–1035. doi:10.1007/s00128-003-0086-1
Acknowledgments
This work was supported through the project TROPHA-Trophic assimilation of inorganic and organic pollutants in terrestrial invertebrate, financed by Fundação para a Ciência e Tecnologia (FCT/POCTI/BSE/48757/2002), and a PhD fellowship awarded to C. F. Calhôa (FCT/SFRH/BD 18942/2004), also from FCT.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Calhôa, C.F., Soares, A.M.V.M. & Loureiro, S. Effects on survival and reproduction of Porcellio dilatatus exposed to different Cd species. Ecotoxicology 21, 48–55 (2012). https://doi.org/10.1007/s10646-011-0762-6
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-011-0762-6