
Abstract
The effects of a commercially available anionic surfactant solution (FFD-6) on growth and morphology of a common green alga (Scenedesmus obliquus) and on survival and clearance rates of the water flea Daphnia magna were studied. The surfactant-solution elicited a morphological response (formation of colonies) in Scenedesmus at concentrations of 10–100 μl l−1 that were far below the No Observed Effect Concentration (NOEC) value of 1,000 μl l−1 for growth inhibition. The NOEC-value of FFD-6 for colony-induction was 3 μl l−1. Daphnia survival was strongly affected by FFD-6, yielding LC50–24h and LC50–48h of 148 and 26 μl l−1, respectively. In addition, clearance rates of Daphnia feeding on unicellular Scenedesmus were inhibited by FFD-6, yielding a 50% inhibition (EC50–1.5h) at 5.2 μl l−1 with a NOEC of 0.5 μl l−1. When Daphnia were offered FFD-6-induced food in which eight-celled colonies (43 × 29 μm) were most abundant, clearance rates (~0.14 ml ind.−1 h−1) were only 25% the rates of animals that were offered non-induced unicellular (15 × 5 μm) Scenedesmus (~0.56 ml ind.−1 h−1). As FFD-6 concentrations in the treated food used in the experiments were far below the NOEC for clearance rate inhibition, it is concluded that the feeding rate depression was caused by the altered morphology of the Scenedesmus moving them out of the feeding window of the daphnids. The surfactant evoked a response in Scenedesmus that is similar to the natural chemically induced defensive reaction against grazers and could disrupt the natural information conveyance between these plankton organisms.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The phytoplankton–zooplankton interaction is of major importance for the functioning of freshwater systems, as it represents the crucial transfer from plant material into animal biomass. Especially members of the genus Daphnia play an important role in lentic ecosystems as phytoplankton grazers by linking limnetic primary production and higher trophic levels (Tollrian and Dodson 1999). Any effects on Daphnia abundance and feeding activity will spread out over the food web, where feeding impairment in particular might result in proliferation of phytoplankton biomass and a dramatic shift from clear water to a turbid state (Scheffer et al. 1993, 2001).
It is already known for years that filtering rates of Daphnia might be a very sensitive indicator for toxic stress (Kersting and van der Honing 1981). Because filtration requires coordination of the movements of appendages by the nervous system, especially neurotoxicants, such as insecticides, may at sub lethal concentrations cause a decrease in Daphnia filtering rates (e.g. Gliwicz and Sieniawska 1986; Fernández-Casselderry et al. 1994; Hartgers et al. 1999; Villarroel et al. 1999; Christensen et al. 2005; Barata et al. 2008; Pestana et al. 2010). However, feeding efficiency also depends on the palatability of the food source. Wealthy information exists on effects of phytoplankton size and morphology on Daphnia filtering rates, where cells exceeding the maximum size of ingestible particles will be inedible for Daphnia (Lampert 1987). When grazers such as Daphnia graze upon cells of the common green alga Scenedesmus, chemicals released during the grazing process promote the formation of colonies to sizes beyond the ingestion capacity of the grazers (Lürling 2003). This chemically induced morphological change in the algal food species not only hampers food intake, but also impairs Daphnia growth and reproduction (Lürling and Van Donk 1996).
Recently, it has been observed that the commercially available anionic surfactant FFD-6 (Skalar chemical) could transform the unicellular green alga Scenedesmus obliquus into colonies (Lürling and Beekman 2002; Lürling 2006). The surfactant FFD-6 induced a response similar to the one evoked by the natural information conveying chemicals. These natural active substances have been extracted from Daphnia and could be identified as different aliphatic sulfates and sulfamates showing strong structural similarity with synthetic anionic surfactants (Yasumoto et al. 2005, 2006, 2008a, b). Hence, despite various anionic surfactants have been classified as not dangerous to the environment with respect to their effect on Daphnia (Verge and Moreno 2000), the FFD-6-induced morphological effect might indicate potential adverse effects on the energy-flow from algae to higher trophic levels at concentrations previously thought to be safe. Therefore, in the current study we tested the hypothesis that FFD-6 induced colonies of the green alga Scenedesmus obliquus decrease the filtering rate of the grazer Daphnia magna.
Materials and methods
Organisms
The green alga Scenedesmus obliquus (Turpin) Kützing SAG 276/3a (originating from the culture collection of the University of Göttingen, Germany) was maintained in a round-glass 1.0 l chemostat system on a standard algal growth medium (Lürling and Beekman 2006). The chemostat system received algal growth medium with a dilution rate of 1.1 day−1. The culture vessel was illuminated with a constant incident irradiance of 100 μmol quanta m−2 s−1 provided by circular fluorescent tubes (Philips TLEM 40 W/33RS) and was placed in a temperature-controlled chamber at 20°C. The chemostat was aerated with sterilized (0.2-μm membrane filter) and moistened air flowing from the bottom to the top of the vessel ensuring optimal mixing of the algal population. Under these conditions, the algal population was mainly unicellular with on average 1.5 cells per colony and a mean particle volume of ~150 μm3.
The grazer Daphnia magna Straus has been isolated from Lake Zwemlust (The Netherlands) and isofemale lineages have been cultured in 1 l vessels containing artificial RT-medium (Tollrian 1993) at a constant temperature of 20°C (pH 7.5–8.0). The animals were fed three times a week ad libitum with S. obliquus harvested from the chemostat systems.
Surfactant
The anionic surfactant FFD-6 was obtained from Skalar BV (Breda, The Netherlands). The product (Skalar 00429040590) was delivered as a FFD-6 surfactant solution that consists of 55% water and 45% mono- and didodecyl disulphanated diphenyloxide, sodium salt (Lürling 2006). The compound was tested without the use of additional solvents.
Effect of FFD-6 on S. obliquus
The effect of FFD-6 on growth and morphology of S. obliquus was tested by running batch experiments in 100 ml Erlenmeyer flasks that contained 50 ml algal growth medium with different concentrations of FFD-6. The surfactant FFD-6 was added at final concentrations of 0, 100, 101, 102, 103 and 104 μl FFD-6 l−1. The Erlenmeyer flasks were closed with cellulose-plugs. Controls and treatments were run for 48 h in triplicates and the initial algal concentration was 1.3 × 104 particles ml−1, i.e. 2 × 106 μm3ml−1. The flasks were incubated in continuous light of 125 μmol quanta m−2s−1 at 20°C on a rotating shaking device (80 rpm). After 48 h the algal concentrations and size distributions were determined in the range 2.5–25 μm equivalent spherical diameter (ESD) using a Coulter® Multisizer II (capillary 100 μm orifice width, Coulter Electronics Limited, Luton, England). The mean particle volumes (MPV) calculated from Coulter data were statistically compared using a one-way ANOVA in the statistical tool pack SPSS with surfactant concentration as the fixed factor. In addition to measurements of MPV, the numbers of cells per colony were determined microscopically by counting at least 100 aggregates (i.e. unicells and colonies) in sub samples that were preserved with Lugol’s fixative (10% v/v). The mean numbers of cells per colony were statistically compared using a one-way ANOVA, followed by a Tukey test to distinguish the significantly different means.
Growth rates were calculated from the increase in algal biovolume measured with the Coulter Multisizer II using the equation μ = {ln(Vt) − ln(V0)} × Δt−1, where Vt = the final algal biovolume (μm3ml−1), V0 = the initial algal biovolume and Δt = the incubation time (d). Cell-based growth rates were calculated from the number of particles determined by the Coulter Multisizer II and the mean number of cells per particle determined by microscope. The chlorophyll-a concentration (μg l−1) and photosynthetic activity of the algae (ΦPSII) was measured with the PHYTO-PAM phytoplankton analyzer (Heinz Walz GmbH, Effeltrich, Germany) using a new reference spectrum for the green alga S. obliquus.
For biovolume- and cell-based growth rates, chlorophyll-a concentrations and photosynthetic activities, EC50 values were determined by non-linear regression (three parameter logistic model) using an iterative process of parameter value adjustments in the toolpack SigmaPlot 2000 for Windows version 6.00. All endpoints were statistically evaluated running one-way ANOVA followed by a Tukey test to distinguish the significantly different means in the statistical tool pack SPSS. The No Observed Effect Concentration (NOEC) was defined as the highest dose at which there was statistically no effect found, while higher dose caused an effect.
In a second biotest, the ½max-MPV values (i.e. FFD-6 concentration causing a 50% increase in the mean particle volume, MPV, of the maximum MPV reached in the treatments) for colony formation in S. obliquus was determined. Inocula of S. obliquus from the chemostat (with 1.85 ± 0.07 cells colony−1) were transferred into 100 ml Erlenmeyer flask that contained 50 ml algal growth medium without (controls) or with FFD-6 (treatments) in the range 100–101 μl FFD-6 l−1 at 1 μl l−1 intervals. The initial algal concentration was 2 × 106 μm3ml−1 and four replicates per treatment were used. At the start of the experiment and after 48 h incubation under conditions as outlined above the algal concentrations and size distributions were determined in the range 2.5–25 μm ESD using the Coulter Multisizer II. The ½max-MPV value for colony formation was determined by non-linear regression using a four parameter logistic model. Because Levene’s test indicated that equal variances could not be assumed, MPV was statistically compared by a non-parametric Kruskal–Wallis test. Dunnett’s T3 test was used for the post-hoc comparison in order to control for Type I error across the pairwise comparisons.
Effect of FFD-6 on Daphnia survival
A cohort of D. magna was created by transferring newborns from the stock cultures into new jars with fresh RT-medium. The animals were fed S. obliquus (~10 mg C l−1) from the chemostat every other day. Neonates born on the same day in these cultures were transferred to new jars with fresh RT-medium and S. obliquus food. After 3 days, the juveniles were collected on a 150 μm sieve, rinsed with RT-medium, and transferred to a beaker with fresh RT-medium. Groups of 15 specimens were selected from the beaker and placed in 250 ml beakers with 150 ml freshly prepared RT-medium without any food. In these beakers the animals were exposed for 48 h to FFD-6 in the concentrations 0, 10−1, 100, 101, 102, 103 and 104 μl FFD-6 l−1. The beakers were incubated at 20°C. The experiment was run in triplicate and the number of surviving animals was recorded after 24 h and 48 h.
Effect of FFD-6 on Daphnia clearance rates
A grazing experiment was performed to study the effect of FFD-6 on clearance rates of D. magna (cf. Lürling and Verschoor 2003). Juvenile three day old D. magna were collected on a 150 μm sieve, rinsed with RT-medium, and transferred to a 250 ml beaker with 150 ml fresh RT-medium. Animals were randomly selected from the beaker and pipetted individually in a drop of medium to 3 ml vials on 24-welled culture plates that contained 2.5 ml of S. obliquus in RT-medium (20 μg chlorophyll-a l−1) and FFD-6 concentrations of 0, 0.5, 5, 50, 500 and 5000 μl l−1. Each vial contained one animal. The algae were mainly unicellular with a mean particle volume of 70 μm3. For each concentration three Daphnia were selected, while one vial received only a drop of medium from the beaker and served as animal-free controls. The culture plates were incubated at 20°C in the dark for 1.5 h. Every 30-min. settled material was resuspended by gently using a pipette and blow some water with it. Initially and after 1.5 h, the samples were analyzed on chlorophyll-a content using the PHYTO-PAM. Clearance rates (CR, in ml ind.−1h−1) were calculated using the equation CR = {ln(AC) − ln(AT)} × ∆t−1 × V/N where AC is the algal concentration (chlorophyll-a in μg l−1) in controls, AT is the algal concentration in the treatments, Δt is the time (1.5 h), V is the culture volume (2.5 ml) and N is the number of animals (=1 per vial). The concentration causing 50% inhibition of the clearance rates (EC50 value) was determined by non-linear regression. The body-length of each animal was measured under a dissecting microscope (40× magnification) from just above the eye until the base of the tail spine. CR were compared by Kruskal–Wallis test and Dunnett’s T3 test was used as follow up test for the pairwise comparisons.
Effect of FFD-6 induced colonies on Daphnia clearance rates
A second grazing experiment was conducted to investigate the effect of unicellular S. obliquus and surfactant-induced colonial S. obliquus on D. magna clearance rates. Colonial S. obliquus were obtained by incubating inocula from the chemostat culture in 100 ml Erlenmeyer flasks containing 50 ml medium in the absence (controls) or presence of FFD-6 (10 μl l−1). Controls and treatments were run for 48 h in quadruplicates as outlined above with identical initial algal concentration of (1.3 × 104 particles ml−1, i.e. 2 × 106 μm3ml−1). Both mean particle volumes (MPV, in μm3) and the number of cells per colony were statistically compared using a t-test. Length and width dimensions of unicells, four-celled colonies and eight-celled colonies were measured using a Leica Quantimet 500 MC image analyzer coupled with a Nikon light-microscope at 500× magnification.
The grazing experiment was performed as described previously, but with two food concentrations (0.1 and 0.2 mg C l−1) and two food types (unicellular and colonial). The experiment was performed with lower food concentrations than the previous one to have maximal clearance rates and to avoid any potential influence of FFD-6 transported with the algal suspensions, i.e. maximally 0.05 and 0.1 μl FFD-6 l−1 was transferred. For each treatment, five Daphnia were selected from a cohort of juvenile 2-day-old D. magna, and five other vials received only a drop of medium from the beaker and served as animal-free controls. The culture plates were incubated for 1.5 h in the dark at 20°C, and hereafter clearance rates were calculated as outlined above. Clearance rates were statistically compared running a two-way ANOVA with food concentration and food morphology (i.e. unicells or colonies) as the fixed factors in the statistical tool pack SPSS.
Results
Effect of FFD-6 on S. obliquus
Exposure of S. obliquus to a concentration gradient of FFD-6 yielded significant differences in the volume- (one-way ANOVA: F 5,12 = 24.9; P < 0.001) and cell-based (F 5,12 = 15.0; P < 0.001) growth rates (Fig. 1). Tukey’s test disclosed two homogenous groups for each endpoint: (1) the concentrations 0, 100, 101, 102, and 103 μl FFD-6 l−1 and (2) the treatments with a concentration of 104 μl FFD-6 l−1. Volume-based growth rates in the range 0–103 μl FFD-6 l−1 were similar (1.04 ± 0.08 d−1), but they were significantly lower in the 104 μl FFD-6 l−1 treatment (0.52 ± 0.08 d−1). A tendency of higher cell multiplication rates in the range 0–10 μl FFD-6 l−1 was observed (Fig. 1), but cell-based growth rates were similar in 0–103 μl FFD-6 l−1 (0.90 ± 0.13 d−1), and only significantly lower in 104 μl FFD-6 l−1 (0.37 ± 0.03 d−1). Chlorophyll-a concentrations were significantly higher in 0, 100, 101, and 102 FFD-6 l−1 (1395 ± 80 μg l−1) than in the 104 μl FFD-6 l−1 treatment (574 ± 11 μg l−1). The chlorophyll-a concentration at 103 μl FFD-6 l−1 (1063 ± 32 μg l−1) was significantly different from the 0, 100 and 104 μl FFD-6 l−1 treatments. The ΦPSII was similar in the range 0 to 103 μl FFD-6 l−1 (0.72 ± 0.01), but significantly lower in the 104 μl FFD-6 l−1 treatment (0.47 ± 0.03). The corresponding EC50 values for volume- and cell-based growth rates as well as those based on PHYTO-PAM derived chlorophyll-a concentrations and photosynthetic activities are given in Table 1. The volume- and cell based growth rates had a NOEC of 103 μl FFD-6 l−1, chlorophyll-a concentration a NOEC of 102 μl FFD-6 l−1 and ΦPSII a NOEC > 104 μl FFD-6 l−1.

Growth rates (μ, d−1) based on the increase in biovolume (filled symbols) and on the increase in cell numbers (open symbols) for Scenedesmus obliquus exposed for 48 h to different concentrations of the surfactant FFD-6. Error bars indicate one standard deviation (N = 3), while similar symbols (a,b and A,B) indicate homogeneous groups that are not different at the 95% level (Tukey test)
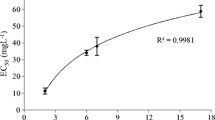
Not only growth rates, but also S. obliquus morphology was affected by the exposure to FFD-6 as significant differences were found in MPV (F 5,12 = 25.9; P < 0.001) and the mean number of cells per colony (F 5,12 = 32.7; P < 0.001) (Fig. 2). Both the MPV and the mean number of cells per colony were significantly higher at 10–1000 μl FFD-6 l−1 compared to controls and the 1 and 104 μl FFD-6 l−1 treatments (Fig. 2). Focusing on the range between 1 and 10 μl FFD-6 l−1 revealed a sigmoid shaped response curve for colony formation in S. obliquus (Fig. 3). The Kruskal–Wallis test indicated significant differences in MPV (χ 2 = 32.6; P < 0.001). Because a significant increase in MPV was found at 4 μl FFD-6 l−1 compared to the control, but not at 3 μl FFD-6 l−1, the latter dose was considered the NOEC for colony formation (Fig. 3). The non-linear model yielded a strong fit and the ½max-MPV value (3.98 μl FFD-6 l−1) was much lower than those based on the other four endpoints (Table 1).

Mean particle volumes (MPV in μm3; filled bars) and mean number of cells per colony (open bars) in Scenedesmus obliquus after 48 h exposure to different concentrations of the surfactant FFD-6. Error bars indicate one standard deviation (N = 3), while similar symbols (a,…, d and A, B) indicate homogeneous groups that are not different at the 95% level (Tukey test)

Mean particle volumes (MPV, μm3) of Scenedesmus obliquus exposed to different concentrations (0–10 μl l−1) of the surfactant FFD-6. The solid line represents the fit of the logistic model: MPV = 177.5 + 246.8/{1 + ([FFD-6]/3.98)−4.93} (r 2 = 0.967). Error bars indicate one standard deviation (N = 4). Similar symbols (a,…, e) indicate homogeneous groups that are not different at the 95% level (Dunnett’s T3 test)
Effect of FFD-6 on Daphnia survival
In controls and FFD-6 concentrations of 10−1 and 100 μl l−1 between 93 and 97% of the test animals had survived after 24 h and 48 h, while all had died in concentrations of 103 and 104 μl FFD-6 l−1. The corresponding LC50–24h and LC50–48h were 148 and 26 μl l−1, respectively (Table 1).
Effect of FFD-6 on Daphnia clearance rates
The clearance rate of similarly sized D. magna (1.34 ± 0.11 mm) feeding on unicellular Scenedesmus was significantly depressed by FFD-6 (Kruskal–Wallis test: χ 2 = 14.1; P = 0.007) (Fig. 4). Dunnett’s T3 test revealed three homogeneous groups: (1) 0–5; (2) 5 and 50 and (3) 0.5, 50, 500 and 5000 μl FFD-6 l−1. The four parameter logistic model yielded an EC50-value of 5.2 μl FFD-6 l−1 (Fig. 4; Table 1).

Concentration–response (clearance rates, CR) curve for Daphnia magna exposed for 1.5 h to the anionic surfactant FFD-6 in the range 0–5000 μl l−1. Error bars indicate one standard deviation (N = 3). The solid line represents the fit of a four parameter logistic model: CR = 0.013 + 0.593/{1 + ([FFD-6]/5.20)−0.96} (r 2 = 0.992). Similar symbols (a, b, c) indicate homogeneous groups that are not different at the 95% level (Dunnett’s T3 test)
Effect of FFD-6 induced colonies on Daphnia clearance rates
Juvenile D. magna (body-length 0.97 ± 0.03 mm) had significantly lower clearance rates (F 1,16 = 68.1; P < 0.001) on FFD-6 induced colonial S. obliquus than on the unicellular S. obliquus food in the controls (Fig. 5). The two-way ANOVA revealed no difference in clearance rates at the two food concentrations of 0.1 and 0.2 mg C l−1 (F 1,16 = 1.07; P = 0.317) and no statistical interaction effect (F 1,16 = 0.65; P = 0.432). The control populations had on average 1.99 (± 0.35) cells per colony, whereas the colonial populations had 7.09 (±0.39) cells per colony. The corresponding MPVs were 145.1 (±0.9) μm3 and 588.5 (±37.1) μm3 for unicells and colonies, respectively. Length and width dimensions of unicells, four- and eight-celled colonies are given in Table 2.

Morphological appearance of the green alga Scenedesmus obliquus after 48 h in the absence (upper left panel a; control) and presence of the anionic surfactant FFD-6 (upper right panel, b; FFD-6). The lower panel c presents the mean clearance rates of juvenile Daphnia magna on the food two types (unicells from controls; open bars, and colonies from FFD-6 treatments; filled bars) at two food concentrations. Error bars indicate one standard deviation (N = 5). Similar symbols (a, b) indicate homogeneous groups that are not different at the 95% level (Tukey test)
Discussion
The results of the current study are in favor of the hypothesis that FFD-6 induced colonies of the green alga Scenedesmus obliquus decrease the filtering rate of the grazer Daphnia magna. The surfactant caused a shift from algal populations dominated by unicells (with average sizes of 15 × 5 μm) to populations in which eight-celled colonies (43 × 29 μm) were most abundant. Forming large colonies moved S. obliquus out of the feeding window of the daphnids. This is corroborated by several other studies that have shown that the formation of colonies allowed Scenedesmus to reach sizes beyond the ingestion capacity of the grazers, which was reflected in lower clearance rates of Daphnia on colonial Scenedesmus compared to unicells (Hessen and Van Donk 1993; Lürling and Van Donk 1996; Lürling et al. 1997; Lürling 2003).
The anionic surfactant FFD-6 promoted the formation of colonies in S. obliquus at concentrations far below those that inhibited growth, which is in concordance with previous studies (Lürling and Beekman 2002; Lürling 2006). The ½max-MPV value of 3.98 μl FFD-6 l−1 for colony induction was far below the NOEC value of 103 μl FFD-6 l−1 for growth inhibition. Thus, the morphological appearance of Scenedesmus was a more sensitive endpoint than the traditionally determined growth inhibition.
The tendency of higher cell multiplication rates in the range 0–10 μl FFD-6 l−1 (see Fig. 1) coincided with the formation of colonies. However, growth in terms of biomass acquisition (biovolume-based) was not affected. This phenomenon has also been observed for the grazer-induced formation of colonies in Scenedesmus (Lampert et al. 1994). For example, the regression obtained for the grazer-induced formation of colonies between volume and cells per colony [log (MPV) = 2.127 + 0.726 × log (cells colony−1); Lampert et al. 1994] is in close match with that of the current study [log (MPV) = 2.235 + 0.662 × log (cells colony−1)]. In both regressions, the slope is less than one, which indicates that individual cell sizes decreased with increasing colony size.
The colonies were formed at doses of 10–103 μl FFD-6 l−1, but not at 104 μl FFD-6 l−1. At the highest dose, FFD-6 impeded growth and the reduced growth rate implied that resources needed for the production of daughter cells, and thus potentially colonies, were acquired at rates too low to trigger the unicell-colony transformation within the experimental period. Those Scenedesmus cultures would theoretically need about 4–5 days to acquire sufficient resources to become colonial too. A similar observation has been made when Scenedesmus was exposed simultaneously to FFD-6 and the growth-(photosynthesis) inhibiting herbicide metribuzin (Lürling 2011).
The EC50 value for (inhibition of) the clearance rate of D. magna was 5.2 μl FFD-6 l−1, which was lower than the LC50–48h (26 μl FFD-6 l−1). This fits in a consistent pattern as for several other toxicants, such as metals and pesticides, EC50 values for feeding inhibition were 5–130 times lower than LC50 values (Gliwicz and Sieniawska 1986; Hartgers et al. 1999; McWilliam and Baird 2002). Since acute studies might provide a good indication of population level effects using an extrapolation factor of 10 between the LC50 and the NOEC (Roex et al. 2000), extrapolation would yield a chronic Daphnia population level NOEC of 2.6 μl FFD-6 l−1, which is about 1.5 mg FFD-6 l−1. Such chronic NOEC value would be lower than the EC50 value for feeding inhibition, but in good agreement with reported NOEC values for chronic exposure of Daphnia to anionic surfactants (Lewis 1991). Nonetheless, prolonged exposure to a toxicant might also further depress Daphnia feeding rates (Hartgers et al. 1999). The acute LC50–48h (26 μl FFD-6 l−1, i.e. ~15 mg l−1) was also in good agreement with literature data for acute toxicity of LAS to D. magna 0.7–270 mg l−1 (Verge and Moreno 2000; Verge et al. 2001; Hodges et al. 2006), and D. similis; ~14 mg l−1 for C12-LAS (Da Silva Coelho and Rocha 2010).
The similarity between the ½max-MPV value and the feeding EC50 value indicates that lower Daphnia clearance rates could also result from FFD-6 transported from the algal suspensions into the grazing vials. However, the low food concentrations employed in the grazing experiment ensured that FFD-6 concentrations were below the NOEC value for feeding inhibition. According to our results, it can be expected that if the organisms are co-cultured at FFD-6 concentrations eliciting the morphological response in S. obliquus, there will also be a direct adverse effect on the feeding activity of the daphnids. Surfactants influence the surface chemistry of food particles and an increase in wettability has been shown causing more food particles to escape the filtering apparatus of the Daphnia (Gerritsen and Porter 1982). Therefore, several mechanisms may act in concert in exerting adverse effects of FFD-6 on the energy flow from S. obliquus to D. magna. Hence, the overall effect on feeding inhibition might have been underestimated in our experiments. Inasmuch as in our experiment with FFD-6 induced colonies a direct FFD-6 effect on clearance rates can be refuted, the most probable explanation for the lower clearance rates of D. magna is that they were caused by the altered morphology of the algal food rather than by toxicity.
The effects of FFD-6 on Scenedesmus morphology and grazer feeding efficiency point towards a reduction of the energy-flow along the food chain for the Scenedesmus–Daphnia system. Lower food intake by daphnids is reflected in lower reproduction and reduced population growth (Lynch 1989), which has also been demonstrated for small-bodied Daphnia on a feed of Scenedesmus colonies (Lürling and Van Donk 1996; Lürling et al. 1997). In this particular plankton system, Scenedesmus responds with the formation of protective colonies to a high threat of being consumed by Daphnia, which is advertised by chemical cues released from the grazers (e.g. Lürling 2003; Von Elert and Franck 1999). This grazer-induced formation of protective colonies is only activated when needed, to avoid unnecessary costs. Therefore, a reliable cue is an important requirement to make chemically induced defenses an evolutionary stable strategy (Harvell 1990) and a stabilizing factor in predator–prey interactions (Lürling et al. 2005). Although the anti-grazing response has also been observed in the green algae Chlorella (Boraas et al. 1998), Chlamydomonas (Lürling and Beekman 2006) and Coelastrum (Van Donk et al. 1999), the marine prymnesiophyte Phaeocystis (Tang 2003) and the cyanobacteria Microcystis (Yang et al. 2006) and Cyanobium (Jezberová and Komarková 2007), it remains unknown how these organisms respond to surfactants. Moreover, results from simple two species plankton systems cannot be extrapolated to multi-species systems or to natural communities (Kratina et al. 2007). Because in nature a wide variety of food items will be available to the grazers, further research should be directed towards a scaling up of experiments to more complex systems to elucidate the community level consequences.
Recently, the chemical nature of the natural colony-inducing chemicals released from Daphnia has been elucidated and the active substances were identified as different aliphatic sulfates (Yasumoto et al. 2005; 2006; 2008a, b). These compounds are closely related to commonly used manmade anionic surfactants. It is therefore not surprising that two commercially available anionic surfactants (sodium dodecyl sulphate and FFD-6) triggered the unicell-colony transformation in S. obliquus as did centrifuged water from a zooplankton culture; and in that way the surfactant effect resembled the natural-induced colony formation (Lürling and Beekman 2002; Yasumoto et al. 2005).
The FFD-6-mimicry of the natural chemical messenger might be far from a reliable proxy of the actual predation risk as advertised by the natural cue. When colonies are induced under low grazing pressure, the benefit, a reduced mortality from grazing, will not outbalance the costs, such as enhanced sinking out of the euphotic zone (Lürling and Van Donk 2000). Consequently, those surfactant-induced colonies can be viewed as maladapted phenotypes with potentially adverse effects on population dynamics of the algae and grazers, where the later might suffer from aggravated anorexia. However, it should be noted that FFD-6 is mainly used in standard spectrophotometric analysis of phosphorus in water (e.g. EPA 1996; Skalar 2002). Thus, environmental concentrations of this particular surfactant will be extremely low. However, FFD-6 belongs to the class of linear alkylbenzene sulfonates (LAS), which are the major anionic surfactants currently on the global market. The structural similarity of many LAS with the 13 different aliphatic sulfates and sulfamates, which have been identified as natural colony-inducing compounds (Yasumoto et al. 2008b), might point toward a more widespread mimicry than solely restricted to sodium dodecyl sulfate and FFD-6. Hence, further research should be directed to study if colony-induction in Scenedesmus is a more general feature of anionic surfactants. In general, anionic surfactant concentrations in surface water are below 100 μg l−1(e.g. Ding et al. 1999; Lara-Martín et al. 2008), but overall these surfactants also cause growth inhibition in green algae at concentrations of an order of magnitude lower than that for FFD-6 (Pavlić et al. 2005).
The surfactant triggered a response in Scenedesmus similar to the natural grazing-associated chemicals, and through effects on Daphnia feeding rates it might also affect production/release of chemical messengers by the grazers. Such disruption of the natural info-chemical system between organisms shows high similarity with endocrine-disrupters that disturb the normal endocrine feedback inside organisms. Disturbance of information exchange between organisms, such as mimicry, amplification or inhibition of natural signals, has the potential of causing organisms to behave/respond maladaptive. Negative effects on key species, such as Daphnia, will not only influence their population dynamics, but could also be spread out over the entire system, through biological interactions (Hanazato 1999). The information component in those interactions is often of major importance in which chemicals are the most dominant form of information carriers. Organisms intensively use such chemical messengers to locate food, find a mate, recognize close kin, mark a territory or detect enemies (Dicke and Takken 2006). With increasing evidence that aquatic organisms exchange information through chemical messengers, the potential disturbance by anthropogenic chemicals, presents an ecological risk (Lürling and Scheffer 2007), which is not covered by standard toxicity tests.
References
Barata C, Alañon P, Gutierrez-Alonso S, Riva MC, Fernández C, Tarazona JV (2008) A Daphnia magna feeding bioassay as a cost effective and ecological relevant sublethal toxicity test for Environmental Risk Assessment of toxic effluents. Sci Tot Env 405:78–86
Boraas ME, Seale DB, Boxhorn JE (1998) Phagotrophy by a flagellate selects for colonial prey: a possible origin of multicellularity. Evol Ecol 12:153–164
Christensen BT, Lauridsen L, Ravn HW, Bayley M (2005) A comparison of feeding efficiency and swimming ability of Daphnia magna exposed to cypermethrin. Aquat Toxicol 73:210–220
Da Silva Coelho K, Rocha O (2010) Assessment of the potential toxicity of a linear alkylbenzene sulfonate (LAS) to freshwater animal life by means of cladoceran bioassays. Ecotoxicology 19:812–818
Dicke M, Takken W (2006) Chemical ecology. From gene to ecosystem. Wageningen UR Frontis Series 16. Springer, The Netherlands
Ding W-S, Tzing S-H, Lo J-H (1999) Occurrence and concentrations of aromatic surfactants and their degradation products in river waters of Taiwan. Chemosphere 38:2597–2606
EPA (1996) Recommended guidelines for sampling and analyses in the Chesapeake Bay monitoring program. EPA 903-R-96-006
Fernández-Casselderry A, Ferrando MD, Andreu-Moliner E (1994) Effect of sublethal concentrations of pesticides on the feeding behavior of Daphnia magna. Ecotox Environ Saf 27:82–89
Gerritsen J, Porter KG (1982) The role of surface chemistry in filter feeding by zooplankton. Science 216:1225–1227
Gliwicz MZ, Sieniawska A (1986) Filtering activity of Daphnia in low concentrations of a pesticide. Limnol Oceanogr 31:1132–1138
Hanazato T (1999) Anthropogenic chemicals (insecticides) disturb natural organic chemical communication in the plankton community. Environ Poll 105:137–142
Hartgers EM, Heugens EHW, Deneer JW (1999) Effect of lindane on the clearance rate of Daphnia magna. Arch Environ Contam Toxicol 36:399–404
Harvell CD (1990) The ecology and evolution of inducible defenses. Q Rev Biol 65:323–340
Hessen DO, Van Donk E (1993) Morphological changes in Scenedesmus induced by substances released from Daphnia. Arch Hydrobiol 127:129–140
Hodges G, Roberts DW, Marshall SJ, Dearden JC (2006) The aquatic toxicity of anionic surfactants to Daphnia magna—a comparative QSAR study of linear alkylbenzene sulphonates and ester sulphonates. Chemosphere 63:1443–1450
Jezberová J, Komarková J (2007) Morphological transformation in a freshwater Cyanobium sp. induced by grazers. Environ Microbiol 9:1858–1862
Kersting K, van der Honing H (1981) Effect of the herbicide dichlobenil on the feeding and filtering rate of Daphnia magna. Verh Int Verein Limnol 21:1135–1140
Kratina P, Vos M, Anholt BR (2007) Species diversity modulates predation. Ecology 88:1917–1923
Lampert W (1987) Feeding and nutrition in Daphnia. In: Peters RH, De Bernardi R (eds) “Daphnia” Memorie dell’Istituto Italiano di Idrobiologia, vol 45, pp 143–192
Lampert W, Rothhaupt KO, von Elert E (1994) Chemical induction of colony formation in a green alga (Scenedesmus acutus) by grazers (Daphnia). Limnol Oceanogr 39:1543–1550
Lara-Martín PA, Gómez-Parra A, González-Mazo E (2008) Sources, transport and reactivity of anionic and non-ionic surfactants in several aquatic ecosystems in SW Spain: a comparative study. Environ Poll 156:36–45
Lewis MA (1991) Chronic and sublethal toxicities of surfactants to aquatic animals: a review and risk assessment. Wat Res 25:101–113
Lürling M (2003) Phenotypic plasticity in the green algae Desmodesmus and Scenedesmus with special reference to the induction of defensive morphology. Ann Limnol Int J Lim 39:85–101
Lürling M (2006) Effects of a surfactant (FFD-6) on Scenedesmus morphology and growth under different nutrient conditions. Chemosphere 62:1351–1358
Lürling M (2011) Metribuzin impairs the unicell-colony transformation in the green alga Scenedesmus obliquus. Chemosphere 82:411–417
Lürling M, Beekman W (2002) Extractable substances (anionic surfactants) from membrane-filters induce morphological changes in the green alga Scenedesmus obliquus (Chlorophyceae). Environ Toxicol Chem 21:1213–1218
Lürling M, Beekman W (2006) Palmelloids formation in Chlamydomonas reinhardtii: defence against rotifer predators? Ann Limnol Int J Lim 42:65–72
Lürling M, Scheffer M (2007) Info-disruption: pollution and the transfer of chemical information between organisms. Trends Ecol Evol 22:374–379
Lürling M, Van Donk E (1996) Zooplankton-induced unicell-colony transformation in Scenedesmus acutus and its effect on growth of herbivore Daphnia. Oecologia 108:432–437
Lürling M, Van Donk E (2000) Grazer-induced colony formation in Scenedesmus: costs of being colonial? Oikos 88:111–118
Lürling M, Verschoor AM (2003) F0-spectra of chlorophyll fluorescence for the determination of zooplankton grazing. Hydrobiologia 491:145–157
Lürling M, De Lange HJ, Van Donk E (1997) Changes in food quality of the green alga Scenedesmus induced by Daphnia infochemicals: biochemical composition and morphology. Freshwater Biol 38:619–628
Lürling M, Arends H, Beekman W, Vos M, Van der Stap I, Mooij WM, Scheffer M (2005) Effect of grazer-induced morphological changes in the green alga Scenedesmus obliquus on growth of the rotifer Brachionus calyciflorus. Verh Int Verein Limnol 29:698–703
Lynch M (1989) The life-history consequences of resource depression in Daphnia pulex. Ecology 70:246–256
McWilliam RA, Baird DJ (2002) Postexposure feeding depression: a new toxicity endpoint for use in laboratiry studies with Daphnia magna. Environ Toxicol Chem 21:1198–1205
Pavlić Ž, Vidaković-Cifrek Ž, Puntarić D (2005) Toxicity of surfactants to green microalgae Pseudokirchneriella subcapitata and Scenedesmus subspicatus and to marine diatoms Phaeodactylum tricornutum and Skeletonema costatum. Chemosphere 61:1061–1068
Pestana JLT, Loureiro S, Baird DJ, Soares AMVM (2010) Pesticide exposure and inducible antipredator responses in the zooplankton grazer, Daphnia magna Straus. Chemosphere 78:241–248
Roex EWM, Van Gestel CAM, Van Wezel AP, Van Straalen NM (2000) Ratios between acute aquatic toxicity and effects on population growth rates in relation to toxicant mode of action. Environ Toxicol Chem 19:685–693
Scheffer M, Hosper SH, Meijer ML, Moss B (1993) Alternative equilibria in shallow lakes. Trends Ecol Evol 8:275–279
Scheffer M, Carpenter SR, Foley JA, Folke C, Walker B (2001) Catastrophic shifts in ecosystems. Nature 413:591–596
Skalar (2002) Skalar methods: phosphate in wastewater; Skalar, The Netherlands
Tang KW (2003) Grazing and colony size development in Phaeocystis globosa (Prymnesiophyceae): the role of a chemical signal. J Plankton Res 25:831–842
Tollrian R (1993) Neckteeth formation in Daphnia pulex as an example of continuous phenotypic plasticity: morphological effects of Chaoborus kairomone concentration and their quantification. J Plankton Res 15:1309–1318
Tollrian R, Dodson SI (1999) Inducible defenses in cladocera: constraints, costs, and multiple predator environments. In: Tollrian R, Harvell CD (eds) The ecology and evolution of inducible defenses. Princeton University Press, New Jersey, pp 177–202
Van Donk E, Lürling M, Lampert W (1999) Consumer induced changes in phytoplankton, inducibility, costs, benefits and the impact on grazers. In: Tollrian R, Harvell CD (eds) The ecology and evolution of inducible defenses. Princeton University Press, New Jersey, pp 89–103
Verge C, Moreno A (2000) Effects of anionic surfactants on Daphnia magna. Tenside Surf Det 37:172–175
Verge C, Moreno A, Bravo J, Berna JL (2001) Influence of water hardness on the bioavailability and toxicity of linear alkylbenzene sulphonate LAS). Chemosphere 44:1749–1757
Villarroel MD, Ferrando MD, Sancho E, Andreu E (1999) Daphnia magna feeding behavior after exposure to tetradifon and recovery from intoxication. Ecotox Environ Saf 44:40–46
Von Elert E, Franck A (1999) Colony formation in Scenedesmus: grazer-mediated release and chemical features of the infochemical. J Plankton Res 21:789–804
Yang Z, Kong F, Shi X, Cao H (2006) Morphological response of Microcystis aeruginosa to grazing by different sorts of zooplankton. Hydrobiologia 563:225–230
Yasumoto K, Nishigami A, Yasumoto M, Kasai F, Okada Y, Kusumi T, Ooi T (2005) Aliphatic sulfates released from Daphnia induce morphological defense of phytoplankton: isolation and synthesis of kairomones. Tetrahedron Lett 46:4765–4767
Yasumoto K, Nishigami A, Kasai F, Kusumi T, Ooi T (2006) Isolation and absolute configuration determination of aliphatic sulfates as the Daphnia kairomones inducing morphological defense of a phytoplankton. Chem Pharm Bull 54:271–274
Yasumoto K, Nishigami A, Aoi H, Tsuchihashi C, Kasai F, Kusumi T, Ooi T (2008a) Isolation and absolute configuration determination of aliphatic sulfates as the Daphnia kairomones inducing morphological defense of a phytoplankton—Part 2. Chem Pharm Bull 56:129–132
Yasumoto K, Nishigami A, Aoi H, Tsuchihashi C, Kasai F, Kusumi T, Ooi T (2008b) Isolation of new aliphatic sulfates and sulfamate as the Daphnia kairomones inducing morphological change of a phytoplankton Scenedesmus gutwinskii. Chem Pharm Bull 56:133–136
Acknowledgments
The research of ML was made possible by a Fellowship from the Royal Netherlands Academy of Arts and Sciences.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Lürling, M., de Lange, H.J. & Peeters, E.T.H.M. Effects of an anionic surfactant (FFD-6) on the energy and information flow between a primary producer (Scenedesmus obliquus) and a consumer (Daphnia magna). Ecotoxicology 20, 1881–1889 (2011). https://doi.org/10.1007/s10646-011-0726-x
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10646-011-0726-x