
Abstract
Lionfish are successful invasive predators in the Caribbean region and inhabit a large range of habitats. Our study in the Caribbean has focused on the relationships between the biological characteristics of lionfish particularly their size, their activities and use of those different habitats. In this study, we observed a high number of lionfish individuals, focusing on the behavioural activities and biological traits in relation to different habitats and environmental characteristics. We monitored 793 individuals, recording their activities, biological traits, and habitat characteristics. Our results report that lionfish are not solitary, but frequently form groups for many activities. We provide evidence of differences between lionfish habitat use according to activity, and the size of individual fish. Considering the size is correlated with age, coral reefs appear to be the preferred habitat of older individuals, whereas the youngest lionfish use a diversity of habitats, ranging from mangroves to coral reefs. In addition, this study suggests that predation of lionfish is age-dependent strategy, and depends on time and the tone of the environment. Lionfish do not only use the head-down posture to catch prey but also horizontal and head-up postures. The youngest lionfish hunt mainly in dark areas and during the night while the older fish were observed hunting mostly during the day and in clear areas. These new aspects of lionfish ecology and behaviour are discussed in light of their invasive success.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Lionfish, Pterois volitans (Linnaeus, 1758) (Scorpaeniformes, Scorpaenidae), is an invasive species in the Caribbean that can cause serious damage to reef ecosystems (Albins and Hixon 2013). It occupies a wide range of habitats such as mangroves, sand, barrier reefs, lagoons, and it has also been reported in man-made structures and docks (Jud et al. 2011). This carnivorous fish eats small reef crustaceans and fish (Valdez-Moreno et al. 2012). The importance of crustaceans in their diet is inversely proportional to the age of the fish, where the largest lionfish prey almost exclusively on teleost fish (Morris and Akins 2009). Lionfish represent an important competitor for native carnivores (O'Farrell et al. 2014; Raymond et al. 2015), reducing the recruitment of reef fish by up to 79% (Albins and Hixon 2008) as well as the invertebrate population (Albins and Hixon 2013). Recent studies on the relationship between lionfish and its possible predators suggest that they are not efficiently controlled by sharks or large groupers, and that direct capture by fishing remains the best control measure to decrease its abundance (Hackerott et al. 2013; Anton et al. 2014; Valdivia et al. 2014). However, current control measures are based on direct capture by divers and fishermen, which has resulted in great effort with local and limited results (Barbour et al. 2011; Frazer et al. 2012).
In their original environment, lionfish are able to adapt to a wide range of habitats (Albins and Hixon 2013). For example, Cure et al. (2014) show the plasticity of lionfish to water visibility: in Guam Island (US territory in the western Pacific Ocean), P. volitans is associated with low water visibility, whereas in the Philippines it is more often found in clear water in reef-associated habitat. Bejarano et al. (2014) showed that reef complexity may influence the abundance of lionfish which tend to choose sites with high density of prey when they hunt. Anton et al. (2014) showed that in the Bahamas, lionfish prefers sites that have a low degree of exposure to wave energy, and during the hours of strongest daylight they take refuge in protected shelters. It is also an opportunist and generalist hunter (Layman and Allgeier 2011) that hunts its prey during hours of low light intensity (Green et al. 2011). Even if lionfish are generally considered as solitary (Fishelson 1975), they may predate in groups during cooperative hunting (Lönnstedt et al. 2014). Many authors mentioned that lionfish hunt preferably at low light intensity (Green et al. 2011). Fishelson (1975) mentioned that in the Red Sea, predation is a nocturnal activity for lionfish. In the Caribbean, other authors observed that the foraging activity occurs around sunrise and sunset, and that lionfish are inactive the rest of the time (Green et al. 2011; Green and Côté 2014). Fishelson (1975) describes that lionfish use their pectoral fins to flush fish and benthic invertebrates into their mouths. Lionfish also blow a jet stream of bubbles to mimic a current, tricking fish into swimming headfirst into their mouths (Albins and Lyons 2012). Jud and Layman (2012) investigated lionfish in an estuary in Florida, finding a pattern of site fidelity.
In a large variety of fishes, differences in use of habitats are related to the size of the fish which generally reflect the age, and activities, principally for reproduction, feeding and predation. For example, in the Caribbean, several species of fishes use mangrove and seagrass beds as nurseries mainly because it provides better protection against predators (Nagelkerken et al. 2001). However, according to the same authors, the dependence for a specific kind of habitat (e.g., seagrass bed, mangrove) may differ according to species. In this context, Dahlgren and Eggleston (2000) demonstrate, in the Nassau Grouper, the existence of a trade-off between growth and being predated that provokes a shift between habitats according to fish body size. Furthermore, when there is a separation between day and night activities, these can occur in different habitats or microhabitats. For example, Jadot et al. (2006) tracked salema (Sarpa salpa L. 1758; Perciformes, Sparidae) and demonstrated that this fish used different habitats according to their resting or displacement activities that to a large extent, depend on, whether it is the night or day.
According to Edwards et al. (2014) and Johnson and Swenarton (2016), the size of lionfish reflects their age. The smallest individuals are more vulnerable when faced with predators such as larger lionfish or groupers (Maljković et al. 2008; Valdez-Moreno et al. 2012). Also small lionfish may have different strategies than older and larger individuals with regard to their choice of habitats, daily activities, and social and predation behaviour.
The literature provides a wide range information about the time predation occurs, solitary or grouping behaviour, and the use of different habitats by lionfish, showing the high plasticity of this species. In order to develop adequate control strategies, it is important to identify lionfish behaviour in relation to their biological traits, particularly their size, habitat characteristics and behavioural activities. To understand these relationships, we used a large variety of factors in different habitats to allow us clarify lionfish life strategies in the southern region of the Mexican Caribbean. Furthermore, we hypothesise that behavioural activities used in a large range of habitats where lionfish were observed, may differ according to age, determined as the size of these fish, the time of day and microhabitat characteristics.
Material and methods
Collecting sites
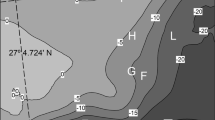
We conducted lionfish observations between May 2013 and May 2014 in three localities in Quintana Roo State, in the Mexican Caribbean (Fig. 1): Mahahual (18°42'N-87°42'W), Banco Chinchorro Biosphere Reserve (18°44'’N-87°19'W), and Xcalak Reef National Park (18°21'N-87°48'W). We surveyed lionfish through direct observations by focal observation and ad libitum records (Altmann 1974) during one-hour SCUBA dive sessions (fins and mask were used in mangroves). We also made recordings with a GoPro Hero III ™ camera. In each locality surveys occurred in different areas recovering different spots. Each site within a given locality was surveyed only once. As previous observations (personal observation, MCGR) indicated higher lionfish abundance on the barrier reef, our surveys were focused in this type of habitat, rather than on the lagoon reef, piers, mangroves and sea grasses. Dives took place between 07:00 and 21:00, to record variations throughout the day. Depth varied from 1 m to 30 m. During nocturnal dives, we used white light (led) that seemed not to affect lionfish the first 20 s, which gave us enough time to describe their behaviour.

Study sites used in this study in the Southern Mexican Caribbean: Mahahual, Banco Chinchorro Biosphere Reserve and the Xcalak National Park reef
Data collection
To optimize data collection, three observers recorded lionfish information. For each lionfish, we recorded the following behaviours and characteristics: 1) total length of fish considering three class of sizes (small: < 15 cm, medium: from 15 cm to 25 cm, and large: ≥ 25 cm 2) tone of fish considering three classes (dark: predominantly red/brown, medium: equal proportion of red/brown and white, and light: more predominance of white); 3) fish posture considering five categories (horizontal, sideways, head-up, head-down and inverted; see details in Table 1 and illustrations in Fig. 2); 4) behaviour at the moment of observation considering three kinds of behaviours (motionless, moving and hunting; see details in Table 1); 5) group composition considering solitary fish (when only one individual is present in a 1.5 m radius circle, or in a cave) vs group of individuals.

Illustrations of five postures identified in this study. Horizontal (a), sideways (b), head-up (c), head-down (d), and inverted (e) (photos by Maria Del Carmen García Rivas)
Lionfish were observed in different environments: mangrove, under docks, reef patches, and barrier reef (see details in Table 2). For each lionfish observed, we recorded the type of environment and characteristics of the microhabitat in where each fish was observed (all details were given in Table 2). Temperature, observation depth, and time were recorded using a TUSA™ IQ-800 computer.
Statistical analysis
A G-test (maximum likelihood ratio test) was used because it is considered a more powerful test than a Chi-square test and is recommended for large sample size, to test significance in frequencies for the following comparisons: 1) postures for each behaviour, 2) solitary vs group for each behaviour, 3) behaviour for each size of fish category, 4) behaviour for each fish tone. In addition, to ascertain possible relationships between activities, postures, biological traits, and environmental characteristics, we used the Self-Organizing Map algorithm (SOM, neural network).
The Self-Organizing Map algorithm (SOM; see Kohonen 2001 for details) was used to analyse non-linear data and/or variables that have skewed distributions, without an a priori transformation. Additionally, the SOM algorithm averages the input dataset using weight vectors and thus removes noise. These features were relevant in our study because we analysed a binary dataset with many zeros (presence/absence). The SOM Toolbox (version 2) for Matlab® developed by the Laboratory of Information and Computer Science at the Helsinki University of Technology (http://www.cis.hut.fi/projects/somtoolbox/) was used. The structure of the SOM for our study consists of two layers of neurons connected by weights (i.e., connection intensities): the input layer comprises 38 neurons (one per variable, but see below) connected to the 793 samples (fish), the output layer is composed of 140 neurons (visualized as hexagonal cells) organized on an array with 14 rows and 10 columns. In the output layer, the neurons act as virtual sites and approximate the probability density function of the input data. During the training, we used a mask function to give a null weight to 24 environmental variables (habitat type, habitat characteristics, time), whereas 14 biological variables or “traits” (e.g., fish activities, postures and tones) were given a weight of 1 so that the ordination process was based on the 14 biological variables only. Setting the mask value to zero for a given component removes the effect of that component on organization. The occurrence probability of a fish trait in a given area in the form of the connection intensity was visualized on the SOM map by a scale of grey, and therefore allowed us to analyse the effect of each environmental variable on the patterning input dataset (fish behaviour, traits). The map size is important in detecting the deviation of the data. Therefore, the network was trained with different map sizes (from 10 to 200 units), and the optimum map size was chosen based on local minimum values for quantization and topographic errors (for a practical example see Céréghino and Park 2009). At the end of the learning process, each individual fish is set in a SOM hexagon. Neurons that are neighbours on the grid are expected to represent neighbouring clusters of fish; consequently, fish separated by a large distance from each other, according to biological variables, are expected to be distant in the output space. A k-means algorithm was applied to cluster the trained map. The SOM units (hexagons) were divided into clusters according to the weight vectors of the neurons, and clusters were justified according to the lowest Davis Bouldin Index, i.e. for a solution with low variance within clusters and high variance between clusters.
Results
A total of 796 fish (224 small, 335 medium, 237 large) were recorded over a period of one year, of which 62 came from Mahahual, 357 from the Banco Chinchorro Biosphere Reserve, and 374 from the Xcalak Reef National Park. The great majority of individuals (81%; n = 644) were recorded during hours of daylight, 12% (n = 92) at dusk, and finally 7% (n = 57) during the night. Sixty-seven percent of fish (n = 530) were observed at low depths, 27% (n = 216) were found at intermediate depths, and only 6% (n = 47) at high depths. The visibility at day was always greater than 10 m, and the average water temperature was 28.5 ± 1.5 °C.
Relationship among behaviour, posture and lionfish characteristics
The most common and significant (G = 606.2, DF = 2, P < 0.001) behaviour of the lionfish was “motionless”, which was observed for 75% of individuals. The “moving” behaviour was observed for 18% (n = 139) of individuals, and the “hunting” behaviour was observed for only 7% of fish. Out of the five postures observed, the most common is the horizontal position (75% of individuals; Fig. 2a), principally observed for “motionless” and “moving” fish (Fig. 3). The sideways posture (Fig. 2b) is only observed for “motionless” fish (Fig. 3). The head-up posture (Fig. 2c) is observed for all behaviours (Fig. 3), whereas although the head-down posture (Fig. 2d) is observed for all three behaviours, it was generally more associated with “hunting” fish (Fig. 3). Finally, the inverted posture (Fig. 2e) is predominantly observed for “motionless” fish (Fig. 3). Our study shows that three postures were used by lionfish while hunting; however, the head-down posture was dominant. We did not observe a significant difference (G = 0.2, DF = 1, P > 0.05) between the number of solitary individuals (n = 224) and lionfish moving in groups (n = 235 groups). The number of individuals found in groups varies from two to a maximum of eight lionfish. Groups of two individuals were the most frequently observed (57%). There is a significant preference to conduct hunting behaviour solitarily (66%; 31 individuals of 47) (Fig. 4). With respect to the other two behaviours, although there is a tendency for “moving” behaviour to be conducted solitarily and “motionless” behaviour to be conducted as a group, none of these tendencies is significant (Fig. 4). Even if all fish size categories present all behaviours (Fig. 5), our results show that for all size categories, all fish were observed significantly more frequently conducting “motionless” behaviour, while hunting was the least frequent behaviour.

Percentage of the five postures described for lionfish for each behaviour observed in the south of Quintana Roo, Mexico. Horizontal (white), sideways (grey), head-up (black), head-down (hatched), inverted (waves). Results from G-test for each behaviour: “motionless” (G = 238, DF = 3, P < 0.001), “moving” (G = 882, DF = 4, P < 0.001), and “hunting” (G = 33, DF = 2, P < 0.001)

Percentage of the group composition of lionfish for each behaviour. Solitary (white) and in groups (black). Results of G-test for each behaviour: moving (G = 3.3, DF = 1, ns), motionless (G = 1.2, DF = 1, ns), and hunting (G = 4.8, DF = 1, P = 0.002)

Percentage of individuals conducting each behaviour for each size category. Motionless (white), moving (grey) and hunting (black). Results of the G-test: small fish (G = 236.9, DF = 2, P < 0.0001), medium fish (G = 217.08, DF = 2, P < 0.0001), and large fish (G = 179.2, DF = 2, P < 0.0001)
In relation to the tone of lionfish, the majority (54%; n = 424) presented a medium tone, 25% a dark tone (n = 191) and 21% a light tone (n = 162). There is a clear dominance of medium tone fish for all behaviours (Fig. 6). However, during hunting behaviour light colour individuals were more frequent than dark ones (30.4% and 17.8% respectively) while there were no differences between the proportion of individuals with light and dark tones for moving (26.3% and 20.5% respectively) and motionless (20.6% and 16.9% respectively) behaviours (Fig. 6).

Percentage of tone characteristics of lionfish, dark (black), medium (grey) and light (white) tones for each behaviour. Results of the G-test: moving (G = 48.67, DF = 2, P < 0.001), motionless (G = 100.7, DF = 2, P < 0.001), and hunting (G = 9.89, DF = 2, P = 0.007)
Relationship between fish behaviour, biological traits and habitat characteristics
In our study, the large majority (85%; n = 675) of lionfish were observed in the wall reef, while only 4% (n = 33) were found in patchy reef and docks, 2% on sand and rocks, and less than 0.5% in mangroves and sea grass. We distinguish six clusters (Fig. 7a) from the SOM analysis (Fig. 7a, b, c, d), which allow us to relate the three behaviours identified in this study with postures, biological traits, and habitat/environment characteristics. Each behaviour can be associated with one or more clusters: “motionless” behaviour is associated with clusters A, B, C and D, “moving” and “hunting” behaviours with clusters E and F with two clearly separated groups reflecting two characteristic strategies that will be specified through SOM analysis (Fig. 7b, c, d). It appears that lionfish size is an important characteristic that separates fish into different clusters: larger fish are associated with clusters F and D; smaller lionfish are found principally in cluster B and in cluster F but in different SOM units than larger lionfish, and intermediate sized fish are regrouped in clusters A, C, E.


Classification of all lionfish to establish a congruent relation between behaviours (see Table 1 for description of behaviours and postures), biological traits (see material and methods for description), and habitat/environmental characteristics (see Table 2 for description of the characteristics) using the Self-Organizing Map algorithm (SOM). Number of clusters construct by SOM analysis (a), SOM map for postures, biological traits and for lionfish behaviours (in bold) (b), SOM map for habitat characteristics (c), and SOM map for environmental characteristics (d)
To clarify the complexity of SOM analysis, we will describe each cluster considering the associated behaviours, postures, biological traits, habitats characteristics and environmental conditions. Cluster A regroups lionfish that are intermediate sized, motionless with a horizontal posture, and with dark or light tones. The fish in this cluster are present in dome structures on the reef, at a depth of between 21 and 30 m, and during the morning. Cluster B regroups lionfish that are small, motionless, mainly in a horizontal posture, but also in inverted postures. These fish have intermediate tones. This is the only cluster that contrasts negatively with the reef habitat (white SOM units) and other habitats such as lagoons, docks, sand and rocks. In these habitats, the small lionfish are in inverted postures, observed in dome structures and mostly associated with rocks, dark support, and half-light in the morning. The SOM units associated with small fish, particularly for sand, lagoon and docks habitats are characterized by no current, low depth (< 10 m), and darkness (night and morning). Cluster C regroups exclusively intermediate size fish with intermediate tone, horizontal, sideways, head-up, inverted postures and motionless. Fish were encountered on the reef and in sea grass habitats. In reef habitats, fish were found at 21–30 m deep, in dome structures with a wide variability of postures, while in sea grass habitat, fish were found at low depths (10 to 20 m) and only in a horizontal posture. The cluster D regroups large motionless fish that only display horizontal postures. In this cluster, fish were found with different categories of tone, but associated with different habitats. The dark tone fish are associated with sand habitat while the intermediate and light tone ones are regrouped with reef habitats that also experience strong currents. This cluster also regroups SOM units with dark support associated with reef habitat, dome structures and intermediate tone fish. Finally, this cluster is associated with activity during the morning. Cluster E regroups exclusively intermediate size lionfish with intermediate tones, and the SOM units are clearly divided between moving behaviour in a horizontal posture, and hunting behaviour with a head-down posture, but all SOM units fit with the reef habitat. Moving behaviour is mostly associated with simple structures, and to a lesser extent with complex ones, with light substrate (clear support), low depth and during the morning. Hunting behaviour is associated with simple structures, light substrate, no current, low depth, during the afternoon, and under conditions of light. Cluster F regroups small light tone and large intermediate tone lionfish that are hunting or moving. All individuals that presented moving behaviour were in a horizontal posture while hunting behaviour fish displayed a head-down posture. However, some larger lionfish hunted with a head-up posture. In both cases, small and large fish, always hunted in complex structures and reef habitat, and to a lesser extent, small fish were observed in sand habitats. Moving behaviour was associated with simple structure in mangroves for small fish, and sand habitat for large lionfish. Small fish were also clearly associated with low depth (< 10 m), no current and dark conditions. The large lionfish were characterized by low depth when hunting. Large fish were active (hunting or moving) during the afternoon and under conditions of light, while small fish were hunting mostly during night and moving during the morning. All large fish were associated with light support (clear substrate) and simple structure when moving, and mostly with complex structure when hunting. All of these results are presented in Table 3.
Discussion
The lionfish were observed at the same frequencies alone or in a group. One of the most important findings from our observations concerns differences between habitat use according to lionfish activities and their size that is considered as equivalent to their age according to several authors (Edwards et al. 2014; Johnson and Swenarton 2016).
The three principal behavioural activities identified for lionfish are: motionless, moving and hunting. Our study confirms that lionfish hunt mainly in the head-down posture, as observed previously by Fishelson (1975) and Green et al. (2011). Older fish also hunt with a head-up posture, and all size of fish may also use a horizontal posture. The head-up and horizontal postures are described for the first time for hunting lionfish. Hunting occurs principally at low depth areas, in reef habitat, generally in complex structures, and on light substrates. Large and intermediate fish hunt during the afternoon in the light, and small or younger lionfish during the night. The latter category is the only lionfish that hunts in other habitats, specifically areas of sand. These results suggest that the hunting strategy of lionfish is size-dependent and therefore, age-dependent, where small/young lionfish use different postures, hours of the day, and sometimes different habitats than older and larger fish. Generally, lionfish were observed resting, and very few individuals were observed hunting. Smaller lionfish were, in fact, rarely observed hunting; they have a higher proportion of crustaceans included in their diet at this stage of their development (Morris and Akins 2009; Layman et al. 2014). Also in younger fish, hunting behaviour is less conspicuous than for older fish. As previously observed in the bluegill sunfish Lepomis macrochirus Rafinesque, 1810 (Perciformes, Centrarchidae) (Mittelbach 1981), younger lionfish may have to learn hunting techniques and are thus unsuccessful piscivores. This diet switch of lionfish from a diet based principally on crustaceans to one based on fish (Morris and Akins 2009; Valdez-Moreno et al. 2012), may explain the two different size-dependent strategies observed in our study. Similar size-dependent diet changes have been reported for other carnivorous reef fish (Lukoschek and McCormick 2001). Additionally, lionfish favour areas with low or no current, probably to facilitate the hunt with the aspiratory system and prey detection. Hunting in open areas, under twilight conditions (as observed for medium and large lionfish), probably involves vision. Lionfish have good visual capacity (Guthrie 1986). However, when hunting, small lionfish present a tendency to remain in the dark. So, under dark conditions, other senses may be used to detect prey, such as tactile capacity or the lateral line to detect crustaceans in the coral structure as demonstrated for other scorpaenids (Montgomery and Hamilton 1996). Another hypothesis to explain why younger lionfish hunt during the night, in the dark and sometimes in different habitats than larger fish, could be that they try to avoid being eaten by larger lionfish, as suggested by stomach content analysis (Valdez-Moreno et al. 2012), which show a high quantity of lionfish remains in our area of observation. The complexity of the habitat is also a good natural defence against predators’ detections and attacks (Crowder and Cooper 1982; Grabowski 2004). Large lionfish can also use horizontal or head-up postures as strategies for the hunting and capture of fish included in their diet (Morris and Akins 2009; Valdez-Moreno et al. 2012) in clear and open areas. These areas present a greater risk to lionfish as they are much more visible to potential predators. The head-down posture makes lionfish more vulnerable to predators in these zones while the horizontal or head-up postures allow lionfish to detect possible threats. The success of the invasion of the lionfish in the Caribbean, and the limited impact of potential native predators (Hackerott et al. 2013; Valdivia et al. 2014) may be related with these evasion strategies. Furthermore, prey fish appear to fail to detect lionfish even in their native range (Lönnstedt and McCormick 2013), so piscivorous larger lionfish may feed during the day in the Caribbean because they are inconspicuous, even in bright day light, due to fish naiveté towards lionfish (Anton et al. 2016). The size and natural defence of larger and older lionfish allow them to display their predatory behaviour in open areas, and are more detectable by possible lionfish´ predators during the day. However, the efficacy of the defence capacities of the lionfish is not well known (Morris and Whitfield 2009).
A specific use of habitat to carry out different behaviours, such as the covered shelters for resting, is common for nocturnal reef fish such as cardinalfish (Perciformes, Apogonidae) to avoid predation (Harmelin-Vivien 1975). The marine current is a determinant factor in the behaviour of other fish species, and it can be related to the choice of staying in one place to limit the energetic cost of displacement in high marine currents (Hobson and Chess 1978). Some studies (Anton et al. 2014; Valdivia et al. 2014) demonstrate that lionfish avoid areas with high swell to limit the loss of energy necessary to maintain their position. Furthermore, these authors suggest that strong currents decrease hunting success because lionfish hunt by squirting water at their prey in order to confuse them, and a strong sea current would affect this hunting strategy. In fact, no hunting behaviour of lionfish was reported in sites with high currents (Anton et al. 2014).
One important and interesting result is that around 50% of observed lionfish were associated with conspecifics. Little is known about the social structure of scorpaenids. Fishelson (1975) mentions that Pterois miles (Bennett, 1828) is a solitary animal. McTee and Grubich (2014) found a greater association among conspecifics in P. miles compared to Pterois radiata (Cuvier, 1829). This association suggests an aggregative behavioural tendency, due to attraction for the same shelter characteristics, particularly during rest behaviour, as observed by Dendrochirus zebra (Cuvier, 1829) (Scorpaeniformes, Scorpaenidae) commonly known as the Zebra lionfish (Moyer and Zaiser 1981). This association could provide a better defence against predators during movement, also hunting in groups could increase the probability of capturing prey (Rizzari and Lönnstedt 2014) but also may lead to intraspecific competition according to Benkwitt (2016). Considering that Pterois spp. are low-speed predators (Albins and Lyons 2012), we propose that hunting in groups may increase their feeding success. In addition, it may be possible that by staying in groups during resting, lionfish may have more chance of detecting predators as frequently observed for various animals including fish (Hoare et al. 2004). Similarly, we regularly observed these fish observing congeners and adapting their behaviour according to others lionfish behaviour when trying to capture one individual of the group (personal observations).
Lionfish can be identified by the intensity of its tone from dark to light with a majority of observed fish presenting an intermediate tone. Our results show that the dark and intermediate tones of the fish are related with inactivity on a dark background or shelter, while the light tones are related to hunting activity on a pale background. Those differences could be adaptive, as observed for some poecilids in South America (Endler 1984). Do lionfish change their tone according to activities and habitat tonalities? Different mechanisms may be proposed to explain this process: 1) physical process by the presence of melanophores, xanthophores and iridophores (Hawkes 1974), 2) physiological process, such as colour changes in parrotfish related to the hormonal level (Cardwell and Liley 1991), or 3) behavioural processes as observed for octopus (Packard and Sanders 1971). Our results suggest that lionfish change colour according to activity and site. However, further studies are needed to understand the relationship between fish tone, activities and habitat tonality. Occasionally, we observed lionfish bleaching (a change from a dark to light colour), when captured by fishermen with a harpoon (personal observation).
Our study on a large number of individuals confirm that lionfish were found in a great diversity of habitats, not only on reefs, as previously shown by Fishelson (1975), where they are more abundant. We found them also under docks, by rocks, sand, patch reefs, and sometimes in mangroves and seagrass beds (Barbour et al. 2010; Biggs and Olden 2011; Claydon et al. 2011; Pimiento et al. 2013), as well as coastal estuaries (Jud et al. 2011). This confirms previous results (Claydon et al. 2011) that demonstrate that lionfish adapt themselves to a great range of habitats. Barbour et al. (2010) suggest that lionfish use mangrove as nurseries. Claydon et al. (2011) proposed that lionfish move from shallow habitats to deep reefs in Turks and Caicos; however, Pimiento et al. (2013) proved in the Bahamas that in both habitats lionfish had a similar size and diet. In our study, lionfish display the three behaviours mostly on the reef, and use other types of habitats to move, or as a rest zone, but practically never to hunt. This observation suggests that in non-reef habitats lionfish are generally in transit, looking for new reefs or similar environment. Moreover, in the Caribbean it seems that habitats other than coral reef are mostly used by younger lionfish and cannot be considered as alternative habitats but as nurseries.
Our results showed that the immobility of lionfish is related mostly to horizontal posture, and this position is usually accompanied by deployment of the pectoral fins. We agree with Fishelson (1975), that this cryptic posture hinders recognition of the lionfish as a predator, by potential prey. Lönnstedt and McCormick (2013) demonstrated, experimentally, that damselfish Chromis viridis (Cuvier, 1830) (Perciformes, Pomacentridae) do not detect lionfish, when compared with other predators. Furthermore, numerous prey fail to recognize the lionfish as predator (Marsh-Hunkin et al. 2013; Black et al. 2014; Anton et al. 2016). This could suggest that a cryptic strategy may be also used by lionfish against predators and explains why lionfish have few predators in the Caribbean (Maljković et al. 2008; Diller et al. 2014).
Our study provides evidence of the importance of understanding lionfish behaviour and the use of its habitat to allow the development of better management strategies for this invasive species. For example, differences in habitat use between younger and older lionfish may be linked with different strategies for their management, such as capture methods or developing traps. In our area of study, this information could allow lionfish to be caught at the right moment and in the right place. Furthermore these results help explain the success of the lionfish invasion, such as an ability to use different habitats according to their age, a capability of adapting themselves to the environment in relation to foraging activity, and escaping from potential predators including cannibalism. In addition, because commonly observed in groups and not only hunting but during resting in shelters, we may expect they are also less vulnerable to predators. This group strategy may also explain the low predation that those fish suffer from potential local predators and their success as invasive species.
References
Albins MA, Hixon MA (2008) Invasive Indo-Pacific lionfish Pterois volitans reduce recruitment of Atlantic coral-reef fishes. Mar Ecol Prog Ser 367:233–238. https://doi.org/10.3354/meps07620
Albins MA, Hixon MA (2013) Worst case scenario: potential long-term effects of invasive predatory lionfish (Pterois volitans) on Atlantic and Caribbean coral-reef communities. Environ Biol Fish 96(10–11):1151–1157. https://doi.org/10.1007/s10641-011-9795-1
Albins MA, Lyons PJ (2012) Invasive red lionfish Pterois volitans blow directed jets of water at prey fish. Mar Ecol Prog Ser 448:1–5. https://doi.org/10.3354/meps09580
Altmann J (1974) Observational study of behaviour: sampling methods. Behaviour 49(3–4):465–472
Anton A, Simpson MS, Vu I (2014) Environmental and biotic correlates to lionfish invasion success in Bahamian Coral Reefs. PLoS One 9(9):e106229. https://doi.org/10.1371/journal.pone.0106229
Anton A, Cure K, Layman CA, Puntila R, Simpson MS, Bruno JF (2016) Prey naiveté to invasive lionfish Pterois volitans on Caribbean coral reefs. Mar Ecol Prog Ser 544:257–269. https://doi.org/10.3354/meps11553
Barbour AB, Montgomery ML, Adamson AA, Díaz-Ferguson E, Silliman BR (2010) Mangrove use by the invasive lionfish Pterois volitans. Mar Ecol Prog Ser 401:291–294. https://doi.org/10.3354/meps08373
Barbour AB, Allen MS, Frazer TK, Sherman KD (2011) Evaluating the potential efficacy of invasive lionfish (Pterois volitans) removals. PLoS One 6(5):e19666. https://doi.org/10.1371/journal.pone.0019666
Bejarano S, Lohr K, Hamilton S, Manfrino C (2014) Relationships of invasive lionfish with topographic complexity, groupers, and native prey fishes in little Cayman. Mar Biol 162(2):253–266
Benkwitt CE (2016) Invasive lionfish increase activity and foraging movements at greater local densities. Mar Ecol Prog Ser 558:255–266. https://doi.org/10.3354/meps11760
Biggs CR, Olden JD (2011) Multi-scale habitat occupancy of invasive lionfish (Pterois volitans) in coral reef environments of Roatan, Honduras. Aquat Invasions 6(3):447–453
Black AN, Weimann SR, Imhoff VE, Richter ML, Itzkowitz M (2014) A differential prey response to invasive lionfish, Pterois volitans: prey naïveté and risk-sensitive courtship. J Exp Mar Biol Ecol 460:1–7. https://doi.org/10.1016/j.jembe.2014.06.002
Cardwell JR, Liley NR (1991) Hormonal control of sex and color change in the stoplight parrotfish, Sparisoma viride. Gen Comp Endocrinol 81(1):7–20. https://doi.org/10.1016/0016-6480(91)90120-U
Céréghino R, Park YS (2009) Review of the self-organizing map (SOM) approach in water resources: commentary. Environ Model Softw 24(8):945–947. https://doi.org/10.1016/j.envsoft.2009.01.008
Claydon JAB, Calosso MC, Traiger SB (2011) Progression of invasive lionfish in seagrass, mangrove and reef habitats. Mar Ecol Prog Ser 448:119–129
Crowder LB, Cooper WE (1982) Habitat structural complexity and the interaction between bluegills and their prey. Ecology 63(6):1802–1813. https://doi.org/10.2307/1940122
Cure K, McIlwain JL, Hixon MA (2014) Habitat plasticity in native Pacific red lionfish Pterois volitans facilitates successful invasion of the Atlantic. Mar Ecol Prog Ser 506:243–253. https://doi.org/10.3354/meps10789
Dahlgren CP, Eggleston DB (2000) Ecological processes underlying ontogenetic habitat shifts in a coral reef fish. Ecology 81(8):2227–2240.
Diller JL, Frazer TK, Jacoby CA (2014) Coping with the lionfish invasion: evidence that naïve, native predators can learn to help. J Exp Mar Biol Ecol 455:45–49. https://doi.org/10.1016/j.jembe.2014.02.014
Edwards MA, Frazer TK, Jacoby CA (2014) Age and growth of invasive lionfish (Pterois spp.) in the Caribbean Sea, with implications for management. Bull Mar Sci 90(4):953–966. https://doi.org/10.5343/bms.2014.1022
Endler JA (1984) Natural and sexual selection on color patterns in poeciliid fishes. In: Zaret TM (ed) Evolutionary ecology of Neotropical freshwater fishes. Springer, New York, pp 95–111. https://doi.org/10.1007/978-94-015-7682-6_7
Fishelson L (1975) Ethology and reproduction of pteroid fishes found in the Gulf of Aqaba (Red Sea), especially Dendrochirus brachypterus (Cuvier), (Pteroidae, Teleostei). Pubb Stn Zool Napoli 39:635–656
Frazer TK, Jacoby CA, Edwards MA, Barry SC, Manfrino CM (2012) Coping with the lionfish invasion: can targeted removals yield beneficial effects? Rev Fish Sci 20(4):185–191. https://doi.org/10.1080/10641262.2012.700655
Grabowski JH (2004) Habitat complexity distrups predator-prey interactions but not the trophic cascade on oyster reefs. Ecology 85(4):995–1004. https://doi.org/10.1890/03-0067
Green SJ, Côté IM (2014) Trait-based diet selection: prey behavior and morphology predict vulnerability to predation in reef fish communities. J Anim Ecol 83(6):1451–1460. https://doi.org/10.1111/1365-2656.12250
Green SJ, Akins JL, Côté IM (2011) Foraging behaviour and prey consumption in the indo-Pacific lionfish on Bahamian coral reefs. Mar Ecol Prog Ser 433:159–167. https://doi.org/10.3354/meps09208
Guthrie DM (1986) Role of vision in fish behaviour. In: Pitcher TJ (ed) Behaviour of teleost fishes, 2nd edn. Chapman & Hall, London, pp 75–113. https://doi.org/10.1007/978-1-4684-8261-4_4
Hackerott S, Valdivia A, Green SJ, Côté IM, Cox CE, Akins L, Layman CA, Precht WF, Bruno JF (2013) Native predators do not influence invasion success of Pacific lionfish on Caribbean reefs. PLoS One 8(7):e68259. https://doi.org/10.1371/journal.pone.0068259
Harmelin-Vivien ML (1975) Place of Apogonid fish in the food webs of a Malagasy coral reef. Micronesica 11:185–198
Hawkes JW (1974) The structure of fish skin. Cell Tissue Res 149(2):159–172. https://doi.org/10.1007/BF00222271
Hoare DJ, Couzin ID, Godin JGJ, Krause JK (2004) Context-dependent group size choice in fish. Anim Behav 67(1):155–164. https://doi.org/10.1016/j.anbehav.2003.04.004
Hobson E, Chess J (1978) Trophic relationships among fishes and plankton in the lagoon at Enewetak atoll, Marshall Islands. Fish Bull 76:133–153
Jadot C, Donnay A, Acolas ML, Cornet Y, Bégout Anras ML (2006) Activity patterns, home-range size, and habitat utilization of Sarpa salpa (Teleostei: Sparidae) in the Mediterranean Sea. Mark Sci 63:128–139
Johnson EG, Swenarton MK (2016) Age, growth and population structure of invasive lionfish (Pterois volitans/miles) in northeast Florida using a length-based, age-structured population model. PeerJ 4:e2730. https://doi.org/10.7717/peerj.2730
Jud ZR, Layman CA (2012) Site fidelity and movement patterns of invasive lionfish, Pterois spp., in a Florida estuary. J Exp Mar Biol Ecol 414:69–74
Jud ZR, Layman CA, Lee JA, Arrington DA (2011) Recent invasion of a Florida (USA) estuarine system by lionfish Pterois volitans/P. miles. Aquat Biol 13(1):21–26. https://doi.org/10.3354/ab00351
Kohonen T (2001) Self-organizing maps, 3rd edn. Springer, Berlin. https://doi.org/10.1007/978-3-642-56927-2
Layman CA, Allgeier JE (2011) Characterizing trophic ecology of generalist consumers: a case study of the invasive lionfish in The Bahamas. Mar Ecol Prog Ser 448:131–141
Layman CA, Jud ZR, Nichols P (2014) Lionfish alter benthic invertebrate assemblages in patch habitats of a subtropical estuary. Mar Biol 161(9):2179–2182. https://doi.org/10.1007/s00227-014-2491-x
Lönnstedt OM, McCormick MI (2013) Ultimate predators: lionfish have evolved to circumvent prey risk assessment abilities. PLoS One 8(10):e75781. https://doi.org/10.1371/journal.pone.0075781
Lönnstedt OM, Ferrari MCO, Chivers DP (2014) Lionfish predators use flared fin displays to initiate cooperative hunting. Biol Lett 10(6):20140281. https://doi.org/10.1098/rsbl.2014.0281
Lukoschek V, McCormick MI (2001) Ontogeny of diet changes in a tropical benthic carnivorous fish, Parupeneus barberinus (Mullidae): relationship between foraging behaviour, habitat use, jaw size, and prey selection. Mar Biol 138(6):1099–1113. https://doi.org/10.1007/s002270000530
Maljković A, Van Leeuwen TE, Cove SN (2008) Predation on the invasive red lionfish, Pterois volitans (Pisces: Scorpaenidae), by native groupers in the Bahamas. Coral Reefs 27(3):501. https://doi.org/10.1007/s00338-008-0372-9
Marsh-Hunkin KE, Gochfeld DJ, Slattery M (2013) Antipredator responses to invasive lionfish, Pterois volitans: interspecific differences in cue utilization by two coral reef gobies. Mar Biol 160(4):1029–1040. https://doi.org/10.1007/s00227-012-2156-6
McTee SA, Grubich JR (2014) Native densities, distribution, and diurnal activity of Red Sea lionfishes (Scorpaenidae). Mar Ecol Prog Ser 508:223–232. https://doi.org/10.3354/meps10847
Mittelbach GG (1981) Foraging efficiency and body size: a study of optimal diet and habitat use by bluegills. Ecology 62(5):1370–1386. https://doi.org/10.2307/1937300
Montgomery CJ, Hamilton AR (1996) Sensory contributions to nocturnal prey capture in the dwarf scorpion fish (Scorpaena papillosus). Mar Freshw Behav Physiol 30(4):209–223
Morris JA Jr, Akins JL (2009) Feeding ecology of invasive lionfish (Pterois volitans) in the Bahamian archipelago. Environ Biol Fish 86(3):389–398. https://doi.org/10.1007/s10641-009-9538-8
Morris JA Jr, Whitfield PE (2009) Biology, ecology, control and Management of the Invasive Indo-Pacific Lionfish: an updated integrated assessment. NOAA Technical Memorandum NOS NCCOS 99
Moyer JT, Zaiser MJ (1981) Social organization and spawning behavior of the pteroine fish Dendrochirus zebra at Miyake-jima, Japan. Jpn J Ichthyol 28:52–69
Nagelkerken I, Kleijnen S, Klop T, van den Brand RACJ, Cocheret de la Morinière E, van der Velde G (2001) Dependence of Caribbean reef fishes on mangroves and seagrass beds as nursery habitats: a comparison of fish faunas between bays with and without mangroves/seagrass beds. Mar Ecol Prog Ser 214:225–235. https://doi.org/10.3354/meps214225
O'Farrell S, Bearhop S, McGill RA, Dahlgren CP, Brumbaugh DR, Mumby PJ (2014) Habitat and body size effects on the isotopic niche space of invasive lionfish and endangered Nassau grouper. Ecosphere 5(10):art123
Packard A, Sanders GD (1971) Body patterns of Octopus vulgaris and maturation of the response to disturbance. Anim Behav 19(4):780–790. https://doi.org/10.1016/S0003-3472(71)80181-1
Pimiento C, Nifong JC, Hunter ME, Monaco E, Silliman BR (2013) Habitat use patterns of the invasive red lionfish Pterois volitans: a comparison between mangrove and reef systems in San Salvador, Bahamas. Mar Ecol 36(1):28–37
Raymond WW, Albins MA, Pusack TJ (2015) Competitive interactions for shelter between invasive Pacific red lionfish and native Nassau grouper. Environ Biol Fish 98(1):57–65. https://doi.org/10.1007/s10641-014-0236-9
Rizzari JR, Lönnstedt OM (2014) Cooperative hunting and gregarious behaviour in the zebra lionfish, Dendrochirus zebra. Mar Biodivers 44(4):467–468. https://doi.org/10.1007/s12526-014-0215-6
Valdez-Moreno M, Quintal-Lizama C, Gómez-Lozano R, García-Rivas MC (2012) Monitoring an alien invasion: DNA barcoding and the identification of lionfish and their prey on coral reefs of the Mexican Caribbean. PLoS One 7(6):e36636. https://doi.org/10.1371/journal.pone.0036636
Valdivia A, Bruno JF, Cox CE, Hackerott S, Green SJ (2014) Re-examining the relationship between invasive lionfish and native grouper in the Caribbean. PeerJ 2:e348. https://doi.org/10.7717/peerj.348
Acknowledgements
We would like to thank CONANP (Comisión Nacional de Areas Naturales Protegidas, Mexico) as well as Banco Chinchorro Biosphere Reserve for the important support provided during fieldwork, and two anonymous reviewers for their helpful commentaries. We are also grateful to Estefanía Medina Bastarrachea for creating the map.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
García-Rivas, M.d.C., Machkour-M’Rabet, S., Pérez-Lachaud, G. et al. Age-dependent strategies related to lionfish activities in the Mexican Caribbean. Environ Biol Fish 101, 563–578 (2018). https://doi.org/10.1007/s10641-018-0718-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-018-0718-2