
Abstract
Seagrass habitats are relevant for numerous nearshore fish species, particularly as nursery grounds. Seagrass meadows are often interspersed with other habitats, what can alter the distribution and abundance of seagrass ichthyofauna. This research aimed to determine whether there is a change in the abundance and biomass of the parrotfish, Sparisoma cretense, in seagrass meadows (Cymodocea nodosa) with varying proximity from rocky reefs, specifically seagrass interiors (>200 m away from reefs) vs. seagrass adjacent to reefs (<10 m away). Sampling was undertaken using a seine net and underwater visual census through an entire annual cycle. Adults were predominantly observed in seagrass adjacent to reefs, which seem to be restricted to incursions of large-sized parrotfish from adjacent reefs. Juvenile abundance did not significantly differ between seagrass interiors and seagrass adjacent to reefs; however, juvenile biomass was greater in seagrass meadows adjacent to reefs compared to those far away from reefs. This pattern was consistent through times for both sampling techniques. These results suggest a transition of juvenile parrotfish from seagrass interiors to seagrass near reefs, so juveniles are located in the vicinity of their adult habitat, i.e. rocky reefs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Seagrass landscapes support a large number of invertebrate and fish species by providing food and shelter (Beck et al. 2001). In particular, seagrass meadows have been routinely viewed as key ‘nursery’ grounds for juveniles of many fish species, including commercially exploited species (Pollard 1984; Gillanders 2006). This results from the large structural complexity (Gullström et al. 2008) and abundance of trophic resources (Bell and Pollard 1989) provided by seagrasses. The presence of fishes in seagrasses, however, may fluctuate through time scales ranging from days to years, according to the specific peculiarities of species’ life cycles, including migrations among distinct habitats (Gillanders 2002; Elsdon and Gillanders 2003). For example, Diplodus annularis is associated with seagrasses during its complete life cycle (Brito et al. 2002), while Spondyliosoma cantharus use seagrasses as a nursery habitat (Espino et al. 2011a, b), occupying rocky reefs and sand-rocky habitats when adults (Lorenzo and González 1997). Seagrass meadows are frequently interspersed with other habitats, such as rocky-reefs, coral reefs, mangroves, and unvegetated sediments (Wernberg et al. 2006; Valentine et al. 2008; Tuya et al. 2010). At a landscape scale, the spatial arrangement and size of these habitats can exert a strong influence on movements of many organisms, affecting patterns in the distribution and abundance of species, e.g. invertebrates and fishes (Tuya et al. 2010, 2011). Several studies have focused on the influence of proximity from a range of habitats on the abundance and structure of fish assemblages inhabiting seagrass meadows, e.g. mangroves-seagrasses (Jelbart et al. 2007), seagrasses-coral reefs (Dorenbosch et al. 2005; Valentine et al. 2008), and mangroves-seagrasses-coral reefs (Nagelkerken et al. 2000; Cocheret de la Morinière et al. 2002; Aguilar-Perera and Appeldoorn 2008). For example, Scarus taeniopterus and S. iserti are species that use seagrasses as nursery habitats and experience a subsequent migration to coral reefs as adults. However, both species showed different migration pace patterns, abruptly for S. taeniopterus and more smooth for S. iserti (Cocheret de la Morinière et al. 2002). The effect of proximity from adjacent habitats on patterns of abundance of seagrass fishes may also change according to the fish body size, reflecting changes in species’ life cycles. For example, mangroves and seagrasses are daytime feeding habitats for a coral reef fish, the French grunt Haemulon flavolineatum, but exclusively for individuals within a particular body size (Verweij et al. 2006).
Different sampling techniques are used to sample fishes in coastal habitats, seagrass beds in particular. Some of the most widely used are capture by fishing gears and visual count methods. Trawls with seine nets are particularly adequate for small-sized individuals (Guest et al. 2003) that are typically overlooked by visual techniques. Underwater visual censuses, on the other hand, are predominantly recommended for large-sized fishes in clear-water habitats (Edgar et al. 2001), because they often escape from trawls. Importantly, different techniques can result in different outcomes when testing for models of spatial and temporal variability in the abundance and biomass of organisms. For example, Harmelin-Vivien and Francour (1992) found that underwater visual census recorded more abundance and biomass than bottom trawls, but trawls recorded more fish species, when studying fish assemblages associated with Posidonia oceanica meadows. As a result, many authors have recommended the combination of different, but complementary sampling methods to accurately study fish assemblages inhabiting seagrass meadows (Harmelin-Vivien and Francour 1992; Bobsien and Brandelberger 2006).
The parrotfish Sparisoma cretense (Linnaeus, 1758) is one of the few temperate species of the Scaridae family (Bernardi et al. 2000). This species is distributed in the eastern coasts of the Mediterranean Sea, the Macaronesian archipelagos (Azores, Madeira, Canaries and Cape Verde) and the northwest coast of Africa (González 1993). It is a necto-benthic species, inhabiting rocky bottoms and seagrass beds on shallow waters to about 50 m depth (Guidetti and Boero 2002). Sparisoma cretense is a daytime feeder, scraping algae, seagrasses and small invertebrates from the substrate with its fused, beak-like jaws. In the Canarian Archipelago, S. cretense is a demersal species, usually observed inhabiting rocky bottoms, especially vegetated reefs, but also in mixed rocky-sandy bottoms, while juveniles can be found in seagrass beds (Mena et al. 1993; Brito et al. 2002; Tuya et al. 2006a; Espino et al. 2011a, b). This species is highly prized for both local recreational and commercial fisheries (Bortone et al. 1991; Bas et al. 1995). This study aimed to: a) determine whether proximity from rocky reefs, here seagrass interiors versus seagrass adjacent to reefs, affects the abundance and biomass of S. cretense inhabiting seagrass meadows; b) determine the temporal and spatial consistency of these patterns via two different sampling techniques, underwater visual census vs. seine nets; and c) assess whether patterns might be altered by the use of different sampling techniques.
Methods
Study area and sampling design
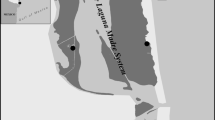
Gran Canaria is an almost circular island (~50 km in diameter) in the middle of the Canarian archipelago, which is located in the north-western African coast. The oceanographic conditions are characterized by the Canary current that flows to the southwest and dominant trade winds that blows from the northeast. Sea surface temperature typically ranges from 18 °C in winter to 24 °C in summer (Navarro-Pérez and Barton 2001). For the purpose of this work, we selected three seagrass meadows constituted by the seagrass Cymodocea nodosa; two meadows are located in the southeast, ca. 2 km apart, while the other meadow is located in the southwest part of the island, ca. 50 km apart (Fig. 1). The area covered by these seagrass meadows ranges between 98,417 and 243,580 m2 at 6–15 m of depth (Ministerio de Medio Ambiente 2002). Each meadow was visited four times through an entire annual cycle: February 2011, May 2011, August 2011 and November 2011. Dates were separated to encompass conditions encountered throughout an entire year. At each meadow two sites, separated by 100 s m, were randomly selected. At each site, fish were sampled at to distances: seagrass adjacent to reefs (ca. <10 m away from reefs) and in seagrass interiors (>200 m away from reefs). Other habitats that have been shown to be relevant during some phases of the life cycle of other parrotfish species are not present in the study region.

Overview of the study area, including the island of Gran Canaria (27°58′N, 15° 36′W) and the location of the three seagrass meadows
Fish and seagrass sampling
Parrotfish were sampled using two techniques: underwater visual censuses (hereafter UVC) and seine nets (hereafter SN). At each site and distance, six replicated 25 m long and 4 m wide transects were firstly laid out randomly during daylight hours, typically between 10:00 and 12:00. The transects were made by two SCUBA divers (3 each diver) simultaneously, surveying different areas to avoid fish to be counted twice. The abundance (total number of individuals) and size (to the nearest 1 cm of total length) of all individuals of S. cretense were recorded on waterproof paper. In the study area, this procedure provides optimal precision and accuracy to account for the abundance and size-structure of both rocky-reef and seagrass fishes (Tuya et al. 2004, 2006a), S. cretense in particular (Tuya et al. 2006b). Secondly, three replicated, 25 m long and 4 m wide, bottom trawls were carried out using a seine net, in approximately the same transect lines where the UVC were previously made. This technique has been successfully applied to study small fish inhabiting seagrass beds in the study area, exclusively capturing small-sized (juvenile) parrotfish (Espino et al. 2011a, b). Thirdly, three descriptors of the seagrass physical structure at each transect line were registered: (1) shoot density (by counting seagrass shoots in n = 6, 25 × 25 cm, quadrats that were deployed in the middle of each transect line), (2) leaf length (by measuring the average leaf length in 20 shoots randomly selected within each quadrat) and (3) seagrass coverage during UVC performance (by registering the distance under a 1 cm × 25 m flexible line transect covered by the seagrass C. nodosa to the nearest cm and subsequent calculation of % coverage, n = 6).
All fish captured by the SNs were preserved in a 10 % of formalin/seawater solution and carried to the laboratory, where the total length (TL ± 1 mm) and weight (W ± 0.001 g) was annotated. Biomasses for fish counted through UVCs were calculated using an available length–weight relationship for the Canarian archipelago (W = 0.0135 L3.0222, González 1991), when total length was ≥20 cm. Biomasses of fish with a TL < 20 cm were calculated using an unpublished length–weight relationship (W = 0.0155 L3.0158, R 2 = 0.9916, n = 312) from our own data. Individuals were classified as adults (TL ≥ 20 cm) and juveniles (TL < 20 cm) using the first maturity size for the study region (González 1991; González and Lozano 1992). This is pertinent, as adult and juvenile S. cretense have different movement capacity and degree of association with seagrasses, similar to other parrotfish species (Ogden and Zieman 1977; Hyndes et al. 2003; Maciá and Robinson 2005).
Statistical analysis
The three structural descriptors of seagrass structure (shoot density, leaf length and seagrass coverage) were analysed by means of 4-way ANOVA models to test if proximity from reefs affected seagrass structure. The model incorporated the factors: (1) ‘Time’ (fixed factor with four levels); (2) ‘Meadow’ (random factor with three levels and orthogonal to ‘Time’); (3) ‘Site’ (random factor with two levels nested within ‘Meadow’ and ‘Time’); and (4) ‘Distance’ (fixed factor with two levels and orthogonal to the previous factors). In particular, analyses focused on the main effects of ‘Distance’ and the interaction term between ‘Distance’ and ‘Time’, to unconfound patterns of parrotfish abundance and biomass with changes in seagrass structure with varying proximity from reefs. The abundance and biomass of adults and juveniles were analysed by means of 4-way ANCOVA models to test for differences among times, meadows, sites within meadows, and distance from reefs. The model followed the same criteria outlined before. The three seagrass structural descriptors were included as covariates to remove variation within the residual term, and so to increase the power to detect significant differences. Prior to the analyses, the Cochran’s test was used to check for homogeneity of variances. Data from UVCs were Ln (X + 1) transformed and rendered homogenous variances for adult biomass (C = 0.0731, P > 0.05), juvenile abundance (C = 0.0947, P > 0.05) and juvenile biomass (C = 0.0421, P > 0.05), but not for adult abundance (C = 0.4324, P < 0.01). In the latter case, the significance level was set at the 0.01, instead of the 0.05 level, as ANOVA is robust to heterogeneous variances for large, balanced, experiments (Underwood 1997). Abundance and biomass data from SNs were transformed to Ln (X + 1) and rendered homogenous variances for both cases (Cochran’s tests, C = 0.122 and C = 0.1334, P > 0.05, respectively). When appropriate, SNK a-posteriori multiple comparison tests were performed (Underwood 1997), particularly when either ‘Time’ or ‘Distance’ were involved as an interaction or as a main effect. The size structure of the parrotfish was compared between seagrass meadows adjacent to reefs and seagrass interiors through a χ 2.
Results
Patterns of abundance and biomass of S. cretense between seagrass interiors and seagrass adjacent to reefs were unconfounded by changes in seagrass structure with varying proximity from reefs: shoot density (‘D’, F = 0.43, P = 0.578; ‘S × D’, F = 0.47, P = 0.713), leaf length (‘D’, F = 0.13, P = 0.751; ‘S × D’, F = 1.19, P = 0.399) and seagrass coverage (‘D’, F = 0.3, P = 0.639; ‘S × D’, F = 0.36, P = 0.786).
UVCs: abundance and biomass patterns with varying proximity from reefs
A total of 644 individuals were observed (20.03 % were adults and 79.97 % juveniles). Adult parrotfish were majorly (88.37 % of all adults individuals) observed in seagrass adjacent to reefs, especially at some sites of two meadows (‘Cabrón’ and ‘Risco’) (Fig. 2a–d). This resulted in a significant ‘D × Si (T × M)’ interaction term for the abundance (P = 0.007, Table 1) and biomass (P = 0.003, Table 1) of adult parrotfish; this masked overall differences in adult abundance and biomass between seagrass interiors and seagrass adjacent to reefs through times (‘D’, ‘T × D’, P > 0.05, Table 1). Juvenile abundance and biomass also changed inconsistently between seagrass adjacent and far away from reefs between sites within meadows and times (‘D × Si (T × M)’, P = 0.001 in both cases). Importantly, the biomass of juveniles overall differed between seagrass interiors and seagrass adjacent to reefs (‘D’, P = 0.038, Table 1, Fig. 3e–h), independently of times (‘T × D’, P > 0.05, Table 1); indeed, juvenile biomass was 1.9 times larger at seagrass adjacent to reefs than in seagrass interiors (Fig. 3e–h). Juvenile abundance was 2.35 times larger at seagrass adjacent to reefs than in seagrass interiors (Fig. 2e–h); however, differences were marginally non-significant (‘D’, P = 0.056, Table 1). Larger juvenile abundance and biomass at seagrass near reefs than seagrass interiors were consistent through times (‘T × D’, P > 0.05 for both juvenile abundances and biomasses, Table 1). The size structure of the juvenile parrotfish significantly differed between seagrass meadows near reefs and seagrass interiors (χ 2 = 17.18, d.f. = 7, P = 0.016) (Fig. 6a).

Mean abundance of adult (a, b, c, d) and juvenile (e, f, g, h) parrotfish Sparisoma cretense recorded through UVCs (n = 6), at seagrass interiors (white bars) and seagrass adjacent to reefs (black bars) at each of two sites within each seagrass meadow and time. Errors bars are standard errors of mean values

Mean biomass of adult (a, b, c, d) and juvenile (e, f, g, h) parrotfish Sparisoma cretense recorded through UVCs (n = 6), at seagrass interiors (white bars) and seagrass adjacent to reefs (black bars), at each of two sites within each seagrass meadow and time. Errors bars are standard errors of mean values
SNs: abundance and biomass patterns with varying proximity from reefs
A total of 312 individuals were collected; all were juveniles. Parrotfish abundance and biomass varied among times inconsistently from meadow to meadow (‘T × M’, Table 2, P < 0.1, SNK tests, Figs. 4 and 5). For example, parrotfish abundance and biomass did not vary among times for ‘Cabrón’, while the other two meadows had larger parrotfish abundance and biomass in summer (Figs. 4 and 5, Table 3). Parrotfish abundance did not overall differed with varying proximity from reefs (‘D’, P = 0.854, Table 2, Fig. 4). However, a larger parrotfish biomass (1.86 times) was observed in seagrass adjacent to reefs than in seagrass interiors (‘D’, P = 0.021, Table 2, Fig. 5). Differences in parrotfish abundance and biomass between distances away from reefs were consistent through times (‘T × D’, P > 0.05, Table 2). Juveniles in seagrass meadows near reefs had a larger mean total length that those in seagrass interiors (8.90 ± 0.27 and 7.12 ± 0.26 cm; mean ± standard error, respectively) (Fig. 6b). In turn, the size structure of the parrotfish significantly differed between seagrass meadows near reefs and seagrass interiors (χ 2 = 46.78, d.f. = 7, P < 0.001).

Mean abundance of the parrotfish Sparisoma cretense at seagrass interiors (white bars) and seagrass adjacent to reefs (black bars), recorded through SNs (n = 3), at each of two sites within each seagrass meadow and time. Errors bars are standard errors of mean values

Mean biomass of the parrotfish Sparisoma cretense at seagrass interiors (white bars) and seagrass adjacent to reefs (black bars), recorded through SNs (n = 3), at each of two sites within each seagrass meadow and time. Errors bars are standard errors of mean values

Size structure (total length, cm) of the parrotfish, Sparisoma cretense, recorded by means of a UVCs and b SNs at seagrass adjacent to reefs (black bars) and seagrass interiors (grey bars)
Discussion
Patterns of adult abundance and biomass with varying proximity from reefs
Despite UVCs did not detect consistent differences in the abundance and biomass of adult S. cretense between seagrass interiors and seagrass adjacent to reefs. This result was likely caused by the low number of registered adults. In any case, the majority (88.37 %) of adults were found in seagrasses juxtaposed with reefs. In the study region, S. cretense is particularly abundant on shallow rocky bottoms, predominantly on reefs covered by macroalgal canopies (Tuya et al. 2006b), while juveniles also occur on C. nodosa seagrass meadows (Mena et al. 1993; Brito et al. 2002; Tuya et al. 2006a; Espino et al. 2011a, b). Therefore, the presence of S. cretense adults in C. nodosa seagrass meadows seems to be restricted to incursions from adjacent reefs, most likely to feed. This is supported by, firstly, the presence of adult parrotfish inhabiting shallow-water rocky reefs in the study region (Tuya et al. 2006b). Secondly, a similar pattern has also been detected in the Mediterranean, where S. cretense is found on rocky reefs, feeding on macroalgae, yet occasional displacements towards seagrass meadows dominated by Posidonia oceanica can also occur to feed on seagrass epiphytes (Bernardi et al. 2000; Kalogirou et al. 2010). In tropical areas, distinct parrotfish species inhabit coral reefs; intermittent displacements of adults towards neighboring seagrasses are common to feed (Randall 1965; Ogden and Zieman 1977; Tribble 1981; Macintyre et al. 1987). This pattern is attributed to the inability of large-sized parrotfish (20–40 cm) to constantly use seagrass canopies as refuges, while small-sized parrotfish (<15 cm) freely move across seagrass canopies (Ogden and Zieman 1977). The same outcome was identified when parrotfish grazing patterns were analyzed (Maciá and Robinson 2005). Adults did not show any consistent temporal pattern in their abundances and biomasses, as observed for individuals on rocky reefs (Tuya et al. 2006b). The low abundance of adult parrotfish somehow reinforces the notion of the large fishing pressure this species is suffering in Gran Canaria (Tuya et al. 2006b).
Adult abundance was larger at meadows ‘Cabrón’ and ‘Risco’, which are nearby to reefs covered by frondose macroalgal canopies at shallow water (e.g. Cystoseira spp. and Sargassum spp., mean coverage ± SE = 85.56 ± 16.67 %, Rodríguez et al. 2008), relative to ‘Veneguera’ meadow, which is juxtaposed to reefs lacking macroalgal canopies (Rodríguez et al. 2008). The abundance of S. cretense on hard bottoms of the study region is enhanced by frondose algal communities that provide food, as well as habitat, for associated epifauna, which are widely found in gut contents of S. cretense (Llinás et al. 1990; González 1991).
Patterns of juvenile abundance and biomass with varying proximity from reefs
Despite a lack of statistical significance, the abundance of juvenile S. cretense detected by means of UVCs was larger (2.35 times) in seagrass near than far away from reefs. Importantly, the biomass of juvenile S. cretense was consistently larger (1.90 times) in seagrass adjacent to reefs than in seagrass interiors through times and meadows. The abundance of juvenile S. cretense registered through SNs did not differ between seagrass adjacent and away from reefs. However, the biomass of juvenile S. cretense was larger (1.86 times) in seagrass adjacent to reefs than in seagrass interiors, consistently through times and meadows. Juveniles inhabiting seagrass adjacent to reefs were, in fact, larger than those in seagrass interiors. This observation complements results from UVCs, and seems to point out towards a transition of juveniles from seagrass interiors to seagrass near reefs as juveniles increase in size, so juveniles are located in the vicinity of their habitat as adults (reefs).
In the study region, S. cretense juveniles display a high degree of association with C. nodosa seagrass, remaining within the canopy to avoid predation; in turn, movements away this canopy are uncommon (F. Espino pers. obs.). Similarly, S. cretense juveniles are tightly associated with vegetation in the Mediterranean; S. cretense juveniles are, indeed, considered as migrant juveniles in seagrass meadows during their early life stages (Kalogirou et al. 2010). In the Canary Islands, most C. nodosa meadows near reefs are separated by narrow sandy corridors, usually about 10–20 m wide, similar to unvegetated ‘haloes’ around coral reefs (Randall 1965; Ogden 1976; Ogden and Zieman 1977). Most likely, this limits the movement of juveniles from seagrass to reefs and vice versa, and helps out to explain the ‘concentration’ of large-sized juveniles in seagrass in the vicinity of reefs. The capacity of juveniles from a range of parrotfish species to move through seagrass canopies has been previously acknowledge (Ogden and Zieman 1977; Hyndes et al. 2003). Individuals around 15–20 cm TL likely have a sufficient movement capacity to move towards reefs. This would imply that juvenile parrotfish remain, in average, ~1.43 years in C. nodosa seagrass meadows before moving to reefs, according to Lozano and González (1993).
Patterns of juvenile abundance and biomass through times
Sparisoma cretense is a gonochorist species that, in the study region, mainly spawns from the end of June to the end of October (González 1991, 1993), so the presence of juveniles is particularly large in late summer and autumn. The spawning period, however, can extend from May to December, depending on environmental conditions. In this context, the release of gametes can be fractionated through several spawning events, so S. cretense is considered as a heterochronic species (Pérez 1979; González 1991, 1993). This wide reproductive period of S. cretense explains the presence of juveniles throughout an entire annual cycle on C. nodosa seagrass meadows, including juveniles ranging from 1 to 19.2 cm TL. This pattern contrast with the recruitment periodicity of some fish species of the Sparidae family (e.g. Spondyliosoma cantharus, Pagellus erythrinus), which recruit on C. nodosa seagrass meadows in shorter periods and, subsequently, show clear cohorts of individuals within the same size range (Espino et al. 2011a, b). A similar outcome was observed in the Mediterranean (Petrakis and Papaconstantinou 1990), where S. cretense recruits were detected all year round, including several cohorts.
UVCs vs. SNs: methodological considerations
Despite both sampling techniques showed a similar pattern for the abundance and biomass of juveniles, two clear differences were detected. First, adults were exclusively registered by UVCs, as adults can easily escape from bottom trawls due to their large swimming capacity. Second, the size distribution of juveniles was different, as small-sized individuals (TL < 10 cm) were underestimated by UVCs. Small fish hide within seagrass canopies and are difficult to spot through UVCs, while an accurate size measure is, at the same time, harder to obtain relative to SNs. The accurate distribution size recorded by SNs reinforces results obtained for the biomass of juveniles; juvenile in seagrass near reefs exhibited a larger mean total length than those at seagrass interiors.
References
Aguilar-Perera A, Appeldoorn RS (2008) Spatial distribution of marine fishes along a cross-shelf gradient containing a continuum of mangrove-seagrass-coral reefs off southwestern Puerto Rico. Estuar Coast Shelf Sci 76:378–394
Bas C, Castro JJ, Hernández-García V, Lorenzo JM, Moreno T, Pajuelo JM, González-Ramos AJ (1995) La Pesca en Canarias y áreas de influencia. Ediciones del Cabildo de Gran Canaria, Las Palmas de Gran Canaria
Beck MW, Heck KL Jr, Able KW, Childers DL, Eggleston DB, Gillanders BM, Halpern B, Hays CG, Hoshino K, Minello TJ, Orth RJ, Sheridan PF, Weinstein MP (2001) The identification, conservation, and management of estuarine and marine nurseries for fish and invertebrates. Bioscience 51:633–641
Bell JD, Pollard DA (1989) Ecology of fish assemblages and fisheries associated with seagrasses. In: Larkum AWD, McComb AJ, Shepherd SA (eds) Biology of seagrasses: a treatise on the biology of seagrasses with special reference to the Australian region. Elsevier, New York, pp 565–609
Bernardi G, Robertson DR, Clifton KE, Azzurro E (2000) Molecular systematics, zoogeography, and evolutionary ecology of the Atlantic parrotfish genus Sparisoma. Mol Phylogen Evol 15:292–300
Bobsien IC, Brandelberger H (2006) Comparison of an enclosure drop trap and a visual diving census technique to estimate fish populations in eelgrass habitats. Limnol Oceanogr Methods 4:130–141
Bortone SA, Van Tassell J, Brito A, Falcón JM, Bundrick CM (1991) A visual assessment of the inshore fishes and fishery resources off El Hierro, Canary Islands: a baseline survey. Sci Mar 55:529–541
Brito A, Pascual PJ, Falcón JM, Sancho A, González G (2002) Peces de las islas Canarias. Catálogo comentado e Ilustrado. Francisco Lemus Editor, La Laguna
Cocheret de la Morinière E, Pollux BJA, Nagelkerken I, van der Velde G (2002) Post-settlement life cycle migration patterns and habitat preference of coral reef fish that use seagrass and mangroves habitats as nurseries. Estuar Coast Shelf Sci 55:309–321
Dorenbosch M, Grol MGG, Nagelkerken I, van der Velde G (2005) Distribution of coral reef fishes along a coral reef-seagrass gradient: edge effects and habitat segregation. Mar Ecol Prog Ser 299:277–288
Edgar GJ, Mukai H, Orth RJ (2001) Fish, crabs, shrimps and other mobile epibenthos: measurements methods for their biomass and abundance in seagrass. In: Short FT, Coles RG (eds) Global seagrass research methods. Elsevier, Amsterdam, pp 255–270
Elsdon TS, Gillanders BM (2003) Reconstructing migratory patterns of fish based on environmental influences on otolith chemistry. Rev Fish Biol Fish 13:217–235
Espino F, Tuya F, Brito A, Haroun RJ (2011a) Ichthyofauna associated with Cymodocea nodosa meadows in the Canarian Archipelago (central eastern Atlantic): community structure and nursery function. Cien Mar 37:157–174
Espino F, Tuya F, Brito A, Haroun RJ (2011b) Spatial variability in the structure of the ichthyofauna associated with Cymodocea nodosa seagrass meadows across the Canary Islands, north-eastern subtropical Atlantic. Rev Biol Mar Oceanogr 46:391–403
Gillanders BM (2002) Connectivity between juvenile and adult populations: do adults remain near their recruitment estuaries? Mar Ecol Prog Ser 240:215–223
Gillanders BM (2006) Seagrasses, fish, and fisheries. In: Larkum AWD, Orth RJ, Duarte CM (eds) Seagrasses: biology, ecology and conservation. Springer, Netherlands, pp 503–536
González JA (1991) Biología y pesquería de la vieja, Sparisoma (Euscarus) cretense (Linnaeus, 1758) (Osteichthyes, Scaridae), en las Islas Canarias. PhD thesis. Universidad de La Laguna, Tenerife, Spain
González JA (1993) Sparisoma (Euscarus) cretense (L.) (Pisces, Scaridae): Zoogeografía y ciclo reproductor en las Islas Canarias. Cour Forsch Senckenberg 159:429–435
González JA, Lozano IJ (1992) Determinación de curvas de madurez sexual en la vieja, Sparisoma (Euscarus) cretense (L.) (Osteichthyes, Scaridae), de Canarias. Bol Inst Esp Oceanogr 8:271–297
Guest MA, Connolly RM, Loneragan NR (2003) Seine nets and beam trawls compared by day and night for sampling fishes and crustaceans in shallow seagrass habitats. Fish Res 64:185–196
Guidetti P, Boero F (2002) Spatio-temporal variability in abundance of the parrotfish, Sparisoma cretense, in SE Apulia (SE Italy, Mediterranean Sea). Ital J Zool 69:229–232
Gullström M, Bodin M, Nilsson PG, Öhman MC (2008) Seagrass structural complexity and landscape configuration as determinants of tropical fish assemblage composition. Mar Ecol Prog Ser 363:241–255
Harmelin-Vivien ML, Francour P (1992) Trawling or visual censuses? Methodological bias in the assessment of fish populations in seagrass beds. Mar Ecol 13:41–51
Hyndes GA, Kendrick AJ, MacArthur LD, Stewart E (2003) Differences in the species- and size-composition of fish assemblages in three distinct seagrass habitats with different plant and meadow structure. Mar Biol 142:1195–1206
Jelbart JE, Ross PM, Connolly RM (2007) Fish assemblages in seagrass beds are influenced by the proximity of mangrove forest. Mar Biol 150:993–1002
Kalogirou S, Corsini-Foka M, Sioulas A, Wennhage H, Pihl L (2010) Diversity, structure and function of fish assemblages associated with Posidonia oceanica beds in an area of the eastern Mediterranean Sea and the role of non-indigenous species. J Fish Biol 77:2338–2357
Llinás O, Moreno E, Rueda MJ (1990) Estudio de la participación de los vegetales en la dieta de la vieja Sparisoma cretense (L., 1758). Secretariado de Publicaciones de la Universidad de La Laguna. Homenaje al Prof. Dr. Telesforo Bravo. Tomo I, Madrid, pp 509–516
Lorenzo JM, González JM (1997) Edad y crecimiento de la chopa Spondyliosoma cantharus (Linnaeus, 1758) (Sparidae) en las islas Canarias. Bol Inst Esp Oceanogr 13:67–73
Lozano IJ, González JA (1993) Determinación del crecimiento de Sparisoma cretense (L.) (Osteichthyes, Scaridae) en Canarias a partir del análisis de frecuencias de talla. Publ Espec Inst Esp Oceanogr 11:73–82
Maciá S, Robinson MP (2005) Effects of habitat heterogeneity in seagrass beds on grazing patterns of parrotfishes. Mar Ecol Prog Ser 303:113–121
Macintyre IG, Grauss RR, Reinthal PN, Littler MM, Littler DS (1987) The Barrier Reef sediment apron: Tobacco Reef, Belize. Coral Reefs 6:1–12
Mena J, Falcón JM, Brito A, Rodríguez FM, Mata M (1993) Catálogo preliminar de la ictiofauna de las praderas de fanerógamas marinas de la isla de Tenerife, islas Canarias. Publ Espec Inst Esp Oceanogr 11:217–222
Ministerio de Medio Ambiente (2002) Estudio Ecocartográfico del Arco Sur de la isla de Gran Canaria
Nagelkerken I, Dorenbosch M, Verberk WECP, Cocheret de la Morinière E, van der Velde G (2000) Importance of shallow-water biotopes of a Caribbean bay for juvenile coral reef fishes: patterns in biotope association, community structure and spatial distribution. Mar Ecol Prog Ser 202:175–192
Navarro-Pérez E, Barton ED (2001) Seasonal and interannual variability of the Canary Current. Sci Mar 65:205–213
Ogden JC (1976) Some aspects of herbivore–plant relationships on Caribbean reefs and seagrass beds. Aquat Bot 2:103–116
Ogden JC, Zieman JC (1977) Ecological aspects of coral reef-seagrass bed contacts in the Caribbean. Proceedings of the 3rd International Coral Reef Symposium 1:377–382
Pérez F (1979) El crecimiento y otros datos biológicos de la ‘vieja’ (Sparisoma cretense Linné, 1758). MsC thesis. Departamento de Ciencias, Universidad de La Laguna
Petrakis G, Papaconstantinou C (1990) Biology of Sparisoma cretense in the Dodecanese (Greece). J Appl Ichthyol 6:14–23
Pollard DA (1984) A review of ecological studies on seagrass-fish communities, with particular references to recent studies in Australia. Aquat Bot 18:3–42
Randall JE (1965) Grazing effect on sea grasses by herbivorous reef fishes in the West Indies. Ecology 46:255–260
Rodríguez M, Pérez O, Ramos E, Monterroso O, Riera R, Sánchez J, Sacramento A, Gil-Rodríguez MC, Cruz A, Morales T, Sangil C, Domínguez S (2008) Estudio de la distribución y tamaño de la población de la especie Cystoseira abies-marina (S. G. Gmelin) C. Agardh, 1820 en Canarias. CIMA Technical Report 29
Tribble GW (1981) Reef-based herbivores and the distribution of two seagrasses (Syringodium filiforme and Thalassia testudinum) in the San Blas Islands (Western Caribbean). Mar Biol 65:277–281
Tuya F, Boyra A, Sánchez-Jerez P, Barberá C, Haroun R (2004) Relationships between rocky-reef fish assemblages, the sea urchin Diadema antillarum and macroalgae throughout the Canarian Archipelago. Mar Ecol Prog Ser 278:157–169
Tuya F, Martín JA, Luque A (2006a) Seasonal cycle of a Cymodocea nodosa seagrass meadow and of the associated ichthyofauna at Playa Dorada (Lanzarote, Canary Islands, eastern Atlantic). Cienc Mar 32:695–704
Tuya F, Ortega-Borges L, Sánchez-Jerez P, Haroun R (2006b) Effect of fishing pressure on the spatio-temporal variability of the parrotfish, Sparisoma cretense (Pisces: Scaridae), across the Canarian Archipelago (eastern Atlantic). Fish Res 77:24–33
Tuya F, Vanderklift MA, Hyndes GA, Wernberg T, Thomsen MS, Hanson C (2010) Proximity to rocky reefs alters the balance between positive and negative effects on seagrass fauna. Mar Ecol Prog Ser 405:175–186
Tuya F, Vanderklift MA, Wernberg T, Thomsen MS (2011) Gradients in the number of species at reef-seagrass ecotones explained by gradients in abundances. PLoS ONE 6:e20190. doi:10.1371/journal.pone.0020190
Underwood AJ (1997) Experiments in ecology: their logical design and interpretation using analysis of variance. Cambridge University Press, Cambridge
Valentine JF, Heck KL Jr, Blackmon D, Goecker ME, Christian J, Kroutil RM, Kirsch KD, Peterson BJ, Beck M, Vanderklift MA (2008) Impacts of exploited species on food web interactions along the coral reef-seagrass interface: a comparison using fished and no-take zones in the Florida Keys National Marine Sanctuary. Ecol Appl 18:1501–1515
Verweij MC, Nagelkerken I, Wartenbergh SLJ, Pen IR, van der Velde G (2006) Caribbean mangroves and seagrass beds as daytime feeding habitats for French grunts, Haemulon flavolineatum. Mar Biol 149:1291–1299
Wernberg T, Vanderklift MA, How L, Lavery PS (2006) Export of detached macroalgae from reefs to adjacent seagrass beds. Oecologia 147:692–701
Acknowledgments
Financial support was received from the Government of the Canary Islands and the EU ERDF within the framework of the PCT MAC 2007–2013 programme, project GESMAR, in close liaison with the insular administration of Cabildo de Gran Canaria. F. Tuya was supported by the MINECO ‛Ramón y Cajal programme. We would like to thank L. Ortega, A. Ulibarri, H. Hernández-Zerpa and T. Sánchez for assisting during collection of field data. This research was conducted under the current environmental and fishery regulations of the Canary Islands.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Espino, F., González, J.A., Haroun, R. et al. Abundance and biomass of the parrotfish Sparisoma cretense in seagrass meadows: temporal and spatial differences between seagrass interiors and seagrass adjacent to reefs. Environ Biol Fish 98, 121–133 (2015). https://doi.org/10.1007/s10641-014-0241-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-014-0241-z