
Abstract
Long and short term investigations have revealed dramatic changes in fish communities after impoundments by reservoirs. However, the process of these changes during impoundment remains poorly studied. In 2005 and 2006, before and during the second stage of impoundment of the Three Gorges Reservoir (TGR), in the upper reaches of the Yangtze, China, we investigated fish catches to assess the impacts of reservoir filling on fish community structure. We conducted sampling at 2 sites, Wanzhou reach, which had been inundated by the first water filling in 2003, and Fuling reach, which still remained a riverine site before this water filling. The results revealed an evident and immediate shift of fish communities from lotic to lentic components. In the Wanzhou reach, during the filling, the abundance percentage of lentic fish increased to 94% compared to 69% before the filling. In the Fuling reach, after the filling, lotic fish declined in abundance from 99% to 85%, while lentic fish increased from less than 1% to 12% in abundance. Based on our investigation and the regulation scheme of the TGR, we predicted that after the completion of the Three Gorges Dam (TGD), there would be typical lacustrine, transitional and riverine zones in TGR, with their corresponding fish assemblages. We suggest that long term monitoring should be conducted to evaluate the future ecological effects, and different strategies implemented in these three zones for conservation purposes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Dams are one of the most negative anthropic activities on rivers. They change water flow regimes, alter the natural environment (Allan 1996), fragment rivers, block migration routes and destroy spawning grounds of fishes, leading to disturbance of the aquatic fauna and biodiversity (Baxter 1977; Dudgeon 2000). The impacts of dams on fish communities have been extensively investigated by many authors. Dramatic changes in fish assemblages have been found after impoundment of reservoirs, which include alternation among lentic and lotic species, and different trophical specialists. In the Warta River, Poland, Penczak and Kruk et al. (2005) found changes in fish assemblage composition after impoundment. Most lithophilous species declined and many phytolithophilous and phytophilous species became dominant. In the Kinkaid Creek drainage of southern Illinois, Taylor et al. (2001) identified a dramatic change in fish community structure that occurred, with the pre-impoundment community dominated by cyprinids and the post-impoundment community dominated by centrarchids. Choi et al. (2008) found significant changes in the fish community in a freshwater system in Korea as a result of impoundment and exotic fish species.
However, all these investigations were conducted either over long term or short term, with the process of these changes during impoundment remaining largely neglected. To diagnose the real reason for the change in fish communities and to be able to adopt suitable conservation tactics, simultaneous investigation during impoundment is needed.
Three Gorges Dam (TGD), constructed on the Yangtze River, is one of the largest dams in the world. This dam will be 193 m high and the inundation will cover over 600 km upstream on the Yangtze River, including areas with the habitat and spawning grounds of many rare, endemic, and commercial fishes, such as Chinese sucker (Myxocyprinus asiaticus), Coreius guichenoti, black carp (Mylopharyngodon piceus), and grass carp (Ctenopharyngodon idella) (Liu and Cao 1992; Ke et al. 1994; Cao 2000; Wu et al. 2004). Construction of the TGD began in 1996 and will be completed in 2009. The aquatic biodiversity has been and will be strongly affected (Wu et al. 2003; Park et al. 2003; Xie 2003).
According to the schedule, the water of the Three Gorges Reservoir (TGR) will be filled up to 175 m above the sea level by the third filling in 2009 (Huang 2004). In June of 2003, the first filling was conducted and raised the water level up to 135 m. Previous investigations have revealed changes in the structure of the aquatic fauna soon after the impoundment, such as changes in fish community, phytoplankton community and fish population density, as well as copepod and cladoceran species composition (Shao et al. 2006; Xue et al. 2006a, b; Zhang et al. 2006).
The second filling was conducted in 2006, which raised the water level up to 156 m from 20 September to 27 October 2006 during 37 days impoundment (Fig. 1). To investigate the processes changing the fish assemblages induced by this impoundment, we undertook field fishing sampling in the Three Gorges Reservoir in 2005 and 2006. Our purpose was to investigate how the changes occurred, to assess the immediate impacts of the filling on fish communities, to predict fish community changes in the future, and to give suggestions for future conservation.

Daily change of water level of the Three Gorges Reservoir (TGR) during the second filling
Materials and methods
Study area and fish collection
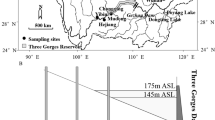
According to the different flow characters, fish sampling took place at two types of habitat in the TGR in the main channel of the upper Yangtze River in 2005 and 2006: Wanzhou reach (30°50′ N, 108°22′ E) and Fuling reach (29°42′ N, 107°23′ E). For these 2 sites, the Wanzhou reach, 286 km up of the TGD, has been inundated as a lacustrine pool by the first filling in 2003, while the Fuling reach, 492 km from the TGD, was still a riverine segment before the second filling in 2006, but was inundated after this filling (Fig. 2). We conducted the fish sampling in Wanzhou reach in October 2005 (before the second filling) and from September to October in 2006 (in the second filling), respectively. As for the Fuling reach, we sampled in September 2006 (before the second filling) and from October 2006 to November 2006 (after the second filling), respectively. The sampling generally continued for about 8–14 days at each site.

Map of China shows the location of Three Gorges Dam (TGD) and sampling sites in the Three Gorges Reservoir (TGR). These broken lines locate the end of backwater formed by the fillings (First filling raised the water level up to 135 m. Second filling raised the water level up to 156 m. Third filling raised the water level up to 175 m)
We investigated more than ten fishing boats in the Wanzhou reach and three fishing boats in the Fuling reach, respectively. Fishes were collected with the local fishing boats using 1.5 cm mesh nylon trap nets or 4, 5, and 5.5 cm mesh nylon gill nets (100–130 m long × 1.2–1.4 m high) at about 7:00 every field day. Normally, one fishing boat had one catch per day using trap nets. Fishermen set the trap nets in scattered locations in the surveyed reach at 6:00, and then collected all the fishes at 6:00 the next day. In the Fuling reach, gill nets were set in an area covering about a 20 km reach, and lasting about 7–24 h each day, according to the fishermen’s experience. In this study, catch per unit effort (CPUE) was expressed as kg·boat−1·(day and night)−1. During the survey period, we identified, counted, and weighed the fishes of each boat. The specimens were sorted and identified according to Ding (1994). Each fish was weighed to the nearest 0.1 g. At the same sampling reach, capturing locations varied somewhat as a result of changes in water level. The trap and gill net, like all fishing gears, have their inherent biases. However, in terms of the aim of this study, these fishing gears give a consistent representation of fish community at the same reach. The consistent biases among capture can mitigate the effect of sampling error when comparing the structure of fish communities.
Data analysis
The data collected after and before the filling from the same sampling sites were compared to analyze the changes. Pair-wise comparisons of fish communities and CPUE were conducted using Chi-square test and t-test, respectively. Moreover, the similarity index was used to estimate the similarity of the fish assemblages between different sampling periods. The Bray–Curtis similarity index, one of the most prevalent quantified tools in comparisons between fish communities, was used here with the following formula:
where BC jk is Bray–Curtis similarity index. n ij and n ik are the number of individuals of ith species in sampling j and k, respectively, and s is the number of species.
Additionally, for the data of each sampling site and period, we calculated its Shannon–Wiener diversity index (Shannon and Wiener 1963), Simpson’s diversity index (Simpson 1949), Buzas and Gibson’s evenness index (Harper 1999), and their 95% confidence intervals by using the program “Past” (Hammer et al. 2001), and then conducted the bootstrap comparison of the diversity indices between different sampling periods through the “compare diversities” module of “Past” (Hammer et al. 2008).
Results
We collected a total of 12,952 fishes representing 57 species of 10 families in the Wanzhou reach in this study (Table 1). Chi-square test analysis indicated significant differences in the structure of fish communities during the filling (P < 0.05). This change could be explained by many aspects, e.g. species richness, taxonomic groups, dominant species, and ecological groups. Species richness evidently increased from 34 species before the second filling to 49 species during the filling. Composition of taxonomic groups also changed due to this impoundment. Before the filling, 26 species (76%) were Cyprinidae; Bagridae and Siluridae each comprised two species (6%); Ictaluridae, Centrarchidae, Gobiidae, and Serranidae comprised 1 species (3%), respectively. During the filling, 36 species (73%) were Cyprinidae; 4 species (8%) Bagridae; 3 species (6%) Cobitidae; Ictaluridae and Siluridae each comprised 2 species (4%); Cichlidae and Catostomidae comprised 1 species (2%), respectively (Fig. 3). Filling also resulted in the replacement of dominant species. Before the filling, Hemiculter leucisculus and Saurogobio dabryi were the dominant species, accounting for 61% and 30% of the abundance, respectively. During the filling, H. leucisculus was still the dominant species but its abundance percentage decreased to 40%. With its abundance increasing to 39%, Hemiculter bleekeri became the dominant species (Table 1). Changes in ecological groups were shown by the increasing number of lentic fishes during filling. Lentic fishes were already the dominant group before the second filling in the Wanzhou reach, which held 69% of the abundance. The abundance proportion increased to 94% during the filling (Fig. 4). In contrast to this, those fishes suited to general habitats decreased sharply during the filling (Fig. 4). The changes in the fish community were also indicated by various community indices between the sampling periods. The qualitative similarity index indicated 47% similarity between the periods before and during the filling. Compared with the fish community before the second filling, there was an increase of 0.53 in the Shannon–Wiener diversity index, 0.15 in the Simpson’s diversity index, and 0.02 in the Buzas and Gibson’s evenness index during the filling (Table 3). Bootstrap comparison revealed that the fish community structure in the Wanzhou reach was altered significantly by the second filling (P < 0.05).

Compositions of taxonomic groups in the different stages in the Wanzhou reach (a) and the Fuling reach (b)

Abundance compositions of ecological groups in the different stages in the Wanzhou reach (a) and the Fuling reach (b)
There were 2,451 fishes belonging to 22 species in 6 families sampled in the Fuling reach (Table 2). Chi-square test indicated that the structure of fish community differed significantly after the filling (P < 0.05). This was shown by the changes in species richness, taxonomic groups, dominant species, and ecological groups. As a result of the filling, species richness increased from 12 to 21 species. Changes in composition of taxonomic groups were evident between sampling periods. Before the filling, 6 species (50%) were Cyprinidae; Bagridae, Cobitidae, and Homalopteridae each comprised of 2 species (17%). After the filling, 13 species (62%) were Cyprinidae; 3 species (14%) Bagridae; 2 species (10%) Homalopteridae; Siluridae, Cobitidae, and Serranidae each comprised 1 species (5%). The abundance proportion of the dominant species decreased after the filling. Coreius guichenoti (43%), Coreius heterodon (28%), and Rhinogobio ventralis (18%) were the dominant species before the filling. After the filling, those species still predominated, but made up 45%, 12%, and 12% of the community, respectively (Fig. 3). It is worth noting that we found clear evidence of the switch between different ecological groups. Lotic fish declined from 99% to 85% in abundance due to the filling (Fig. 4). Conversely, lentic fish increased from 0.4% to 12% on abundance (Fig. 4). We also calculated the ecological community indices for comparison of changes between the sampling periods as well. The Bray–Curtis qualitative similarity index indicated 69% similarity for the fish community after the filling. The Shannon–Wiener diversity index increased by 0.43 and Simpson’s diversity index increased by 0.05 after the filling. However, the Buzas and Gibson’s evenness index decreased by 0.04 (Table 3). So, the significant differences in the diversity indices also indicated that the structure of fish community in the Fuling reach differed significantly after the second filling (P < 0.05).
In addition, we also compared differences of CPUE between the sampling periods. The results showed that no significant differences in CPUE induced by the impoundment (P > 0.05) (Table 4) in either the Wanzhou or the Fuling reach.
Discussion
Shift in species composition
Previous investigations have shown that fish assemblage structure is correlated with environmental characteristics, mainly water discharge in rivers. Alternation of flow regimes may cause a large variety of biotic responses, such as the loss of fishes adapted to turbid river habitats, elimination of pelagic spawning fishes, and loss of migratory fish species. Agostinho et al. (1994) described changes in the fish fauna and fisheries in the Itaipu Reservoir (Brazil) between pre- and post-impoundment: only 83 species were caught after impoundment, compared to 113 prior to inundation; fish species diversity increased initially but declined thereafter. Characiformes and Siluriformes were the dominant groups both before and after impoundment. Two species previously sporadic in the catch, the planktivore Hypophthalmus edentatus and the insectivore Auchenipterus nuchalis, became prolific in the reservoir. Our results also revealed clear changes in fish communities from lotic to lentic species during the filling of the reservoir. Depending on the different areas in the TGR, the fish compositions were different, the changes of environmental conditions due to inundation were different, and so were the changes of the fish compositions after inundation. However, in both sites, the impoundment led to a more lacustrine environment, resulting in changes of fish communities more adapted to this habitat.
Since the first filling in 2003, the Wanzhou reach has become lacustrine. Our results indicated that lentic fishes were already the dominant group after the first filling, e.g. Hemiculter species. More lentic species appeared during the second filling. Indeed, the fish community in the Fuling reach differed from that in the Wanzhou reach because the Fuling reach was not inundated by the first filling and still remained lotic. Despite of the significant changes in fish community structure, the composition still mainly consisted of lotic species after the second filling, including C. heterodon, C. guichenoti, R. ventralis, and Rhinogobio cylindricus. The change was mainly the increase of small lentic or generalist species, such as Pelteobagrus vachelli, Pelteobagrus nitidus, Silurus meridionalis, Abbottina obtusirostris, S. dabryi, Carassius auratus, Cyprinus carpio, Hemibarbus labeo, Hemibarbus maculates, and Siniperca kneri, while the lotic species, such as C. heterodon, R. ventralis, Rhinogobio typus, and R. cylindricus, which were not adapted to the slow flow, moved upstream to more lotic habitats.
Compared with the long term monitoring of the impacts of impoundment, our results show that the shift of fish assemblages was very quickly, since our investigations were conducted during the water filling period or immediately after the filling. We believe that further changes will soon appear after completion of the TGD.
Productivity change
It is interesting to note that, between the sampling periods, the changes in CPUE in the Wanzhou reach and the Fuling reach were not statistically different though their fish community structures have changed. Generally speaking, impoundment may change the hydraulic characteristics of the inundated area, and result in changes in productivity. In the present case, however, since the time period was too short, the difference of CPUE could not be accounted by the change in primary productivity. We suggest this may be due to the resettlement of the fishes. The non-significant differences of CPUE may indicate that a very quick shift of fish component occurred during the impoundment.
Prediction of fish assemblage in the future
Oliveira et al. (2005) analyzed the structures of fish assemblages in the Itaipu Reservoir, on the Brazil–Paraguay border, and described three zones, riverine, transitional and lacustrine, along a longitudinal or river-dam gradient. After the full completion, the flow regulation of the TGD is designed to lower the reservoir level to 145 m (the flood control level, FCL) during the flood season (from May to September), and raise the reservoir level again to 175 m, the normal pool level (NPL) from October to April (dry season). Thus, we predict that there will be a large transitional zone in the future. So, after the completion the TGD, there should be typical lacustrine, transitional and riverine zones, and their corresponding fish assemblages.
The replacement of lotic by lentic fish assemblages after impoundment has been reported in many cases, for example in the Dalešice reservoir in the Czech Republic (Peňáz et al. 1999), and the Fuchunjiang, Xinanjiang, and Danjiangkou reservoirs in China (Zhong and Power 1996). Normally, those hydroelectric reservoirs were characterized by riverine and lacustrine zones. However, Gomes and Miranda (2001) suggested that the short term water retention created neither riverine nor lacustrine fish assemblages within the reservoir. Considering the flow regulation scheme of the TGR, we predict that the alternation of water level every year may seriously affect the structure of fish community. A transitional zone may form along the reach where the water level will alternate between 145 m and 175 m. In this transitional zone, a very rapid shift between different fish assemblages will occur each year in the future.
Suggestions for conservation
After fully completion, the TGR will cover a 600 km reach of the upper Yangtze River. The regulation of water flow, the alternation of water level, and the operation of the hydroelectric power station will create serious impacts on fish communities upstream of the dam. Therefore, conservation measures for fishes should be taken as soon as possible. Considering the results of our present investigation and some previous research, we suggest that different conservation strategies should be employed for the different zones.
The impounded lacustrine zone can be a large area for nursery and feeding by commercial species (Jackson and Marmulla 2007). In the TGR, M. piceus, C. idella, Hypophthalmichthys molitrix, and Aristichthys nobilis, known as the “four major Chinese carps”, will mainly use this area. However, it has been noticed that their juveniles have been evidently overfished by trap nets of small mesh size (Institute of Hydrobiology, unpublished data), which may possibly lead to recruitment failure. In terms of conservation interest for fishes, we suggest that this gear should be banned. Moreover, some exotic species have successfully colonized the lacustrine zone due to unauthorized introduction, such as Ameiurus melas, Tinca tinca, and Ictalurus punctatus. Those species will threaten the native fishes through competing habitats and prey (Gehrke et al. 2002; Chen et al. 2007). Therefore, the fishery department should seriously examine the use of these introduced species in aquaculture and legally prohibit any unauthorized introductions.
After completion of the TGD, a relatively natural flow regime will remain in the riverine zone, which would become the last refuge for the endemic fishes of upper Yangtze River. Due to the flow regulation in transitional and lacustrine zones, the endemic fishes would move upstream to the riverine zone. In particular, during or after the filling of water each year, the endemic fishes would rapidly congregate in the riverine zone where they would be exposed to heavy exploitation by net fisheries. Therefore, we suggest that catch sizes and catch limit should be established as part of capture quotas to avoid overfishing in riverine zone, especially during the water filling period.
In contrast to other zones, the fish community in the transitional zone could be more severely impacted by frequent flow regulation. Though we predicted the shift between lotic and lentic fish assemblages in our study, we still have very limited knowledge in this complex zone. This leads to difficulty in establishing a conservation strategy for this zone. We suggest that long term surveys should be conducted in this zone to investigate the ecological effects of the flow regulation, and establish suitable conservation measures.
References
Allan JD (1996) Stream ecology: structure and function of running waters. Chapman & Hall, London
Agostinho AA, Julio HF Jr, Petrere M Jr (1994) Itaipu reservoir (Brazil): impacts of the impoundment on the fish fauna and fisheries. In: Cowx IG (ed) Rehabilitation of freshwater fishries. Fishing News Books, Bodmin, pp 171–184
Baxter RM (1977) Environmental effect of dams and impoundments. Annu Rev Ecol Syst 8:255–283
Cao W (2000) The construction of the nature reserves of the endemic fishes in the upper reaches of the Yangtze River and the consideration of the correlative problem. Resour Environ Yangtze Basin 9:131–132 (in Chinese)
Chen P, Wiley EO, Mcnyset KM (2007) Ecological niche modeling as a predictive tool: silver and bighead carps in North America. Biol Invasions 9:43–51
Choi JS, Choi JK, Noakes DLG (2008) The impacts of exotic species and stream impoundment on a freshwater lake fish community: Lake Soyang, Korea. J Sci Technol Human 6:85–98
Ding R (1994) The fishes of Sichuan, Chian. Sichuan Publishing House of Science and Technology, Chengdu (in Chinese)
Dudgeon D (2000) Large-scale hydrological changes in tropical Asia: Prospects for Riverine Biodiversity. Bioscience 50:793–806
Gehrke PC, Gilligan DM, Barwick M (2002) Changes in fish communities of the Shoalhaven River 20 years after construction of Tallowa Dam, Australia. River Res Appl 18:265–286
Gomes LC, Miranda LE (2001) Riverine characteristics dictate composition of fish assemblages and limit fisheries in reservoirs of the upper Paraná River basin. Regul Rivers Res Manage 17:67–76
Huang Z (2004) Eco-environmental monitoring and protection measures for the Three Gorges Project. Sci Technol Rev 12:26–30 (in Chinese)
Jackson DC, Marmulla G (2007) The influence of dams on river fisheries. In: Marmulla G (ed) Dams, fish and fisheries. FAO Fisheries Technical Paper, Rome, p 419
Ke F, Wei Q, Luo J, Yang W (1994) Effect of Changjiang River Three-Gorges projects on fishery resources and remedy measures. Freshwater Fish 24(1):6–9 (in Chinese)
Liu J, Cao W (1992) Fish resources of the Yangtze River basin and the tactics for their conservation. Resour Environ Yangtze Basin 1(1):17–23 (in Chinese)
Oliveira EF, Minte-Vera CV, Goulart E (2005) Structure of fish assemblages along spatial gradients in a deep subtropical reservoir (Itaipu Reservoir, Brazil–Paraguay border). Environ Biol Fish 72(3):283–304
Park YS, Chang J, Lek S, Cao W, Brosse S (2003) Conservation strategies for endemic fish species threatened by the Three Gorges Dam. Conserv Biol 17:1748–1758
Peňáz M, Baruš V, Prokeš M (1999) Changes in the structure of fish assemblages in a river used for energy production. Regul Rivers Res Manage 15:169–180
Penczak T, Kruk A (2005) Patternizing of impoundment impact (1985–2002) on fish assemblages in a lowland river using the Kohonen algorithm. J Appl Ichthyol 21(3):169–177
Harper DAT (1999) Numerical palaeobiology. Wiley, Chichester
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electronica 4(1):9 http://palaeo-electronica.org/2001_1/past/issue1_01.htm
Hammer Ø, Harper DAT, Ryan PD (2008) PAST-palaeontologia statistic, ver. 1.81. http://folk.uio.no/ohammer/past/past.pdf
Shannon CE, Wiener W (1963) The mathematical theory of communication. University of Illinois Press, Urbana 117 pp
Shao X, Li D, Cai Q (2006) The composition of the fish community in Xiangxi Bay and resources evaluation. Acta Hydrobiol Sin 30(1):70–74 (in Chinese)
Simpson EH (1949) Measurement of diversity. Nature 163:688
Taylor CA, Knouft JH, Hiland TM (2001) Consequences of stream impoundment on fish communities in a small North American drainage. Regul Rivers Res Manage 17:687–698
Wu J, Huang J, Han X, Gao X, He F, Jiang M, Jiang Z, Primack RB, Shen Z (2004) The Three Gorges Dam: an ecological perspective. Front Ecol Environ 2(5):241–248
Wu J, Huang J, Han X, Xie Z, Gao X (2003) Three-Gorges Dam—experiment in habitat fragmentation. Science 300:1239–1240
Xie P (2003) Three-Gorges Dam: risk to ancient fish. Science 302:1149
Xue J, Ye L, Cai Q (2006a) Variation of copepod from maoping to Guizhou in the Gorges Reservoir before and after impoundment. Acta Hydrobiol Sin 30(1):113–115 (in Chinese)
Xue J, Ye L, Cai Q, Liu J (2006b) Variation of cladocerans from Maoping to Guizhou in the Three Gorges Reservoir before and after impoundment. Acta Hydrobiol Sin 30(1):58–63 (in Chinese)
Zhang Y, Zheng B, Liu H (2006) Characteristics of Phytoplankton composition with analysis of its impact factors after impounding of the Three Gorges Reservoir. Resour Environ Yangtze Basin 15(2):254–258 (in Chinese)
Zhong Y, Power G (1996) Environmental impacts of hydroelectric projects on fish resources in China. Regul Rivers Res Manage 12:81–98
Acknowledgements
This study was funded by the key project of CAS (KZCX1-YW-08-01), the National Natural Science Foundation of China (NSFC 40432003), and the Three Gorges Project eco-environmental monitoring system (SX[2006]-013). Also, we are very grateful to the anonymous reviewers for the advice to our work, especially one who corrected English for the draft of this paper in detail.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gao, X., Zeng, Y., Wang, J. et al. Immediate impacts of the second impoundment on fish communities in the Three Gorges Reservoir. Environ Biol Fish 87, 163–173 (2010). https://doi.org/10.1007/s10641-009-9577-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-009-9577-1