
Abstract
Demographic analyses were conducted for the dusky shark, Carcharhinus obscurus, in the Northwest Atlantic. Fishery-independent, fishery-dependent, and previously published data were used to estimate age specific by-catch mortality and reproductive cycle for incorporation into the analyses. Correspondence analyses were conducted for viability (dead or alive), shark length, month, soaktime, and region. Dead sharks were associated with small size, long soak times, and the spring season, while live sharks were associated with large size and shorter soak times. Data on reproduction suggested a 3-year reproductive cycle consisting of a 2-year gestation period and a 1-year resting period. Litter sizes ranged from 3–12 embryos. Decreasing age-specific fishing mortality and a 3-year reproductive cycle were used in age structured life tables which incorporated stochasticity in life history parameters through Monte Carlo simulation to estimate annual population growth. Demographic analyses showed that the dusky shark population would decline even at low levels of fishing mortality despite low natural mortality for neonates. The demographics of this species make it among the most vulnerable to excessive fishing mortality, and suggest stringent regulatory measures may be required to recover collapsed populations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The dusky shark, Carcharhinus obscurus, is a common coastal and pelagic shark that inhabits warm-temperate and tropical coastal waters of the western North Atlantic, ranging from southern New England to the Caribbean and Gulf of Mexico to southern Brazil. It avoids areas of lower salinity and is rarely found in estuarine environments (Compagno 1984; Musick and Colvocoresses 1986; Musick et al. 1993).
This species undertakes long temperature-related migrations along the east coast of the U.S. Individuals move north as water temperatures increase with the onset of spring and return south with the onset of fall (Musick and Colvocoresses 1986). Juveniles occupy highly productive coastal nurseries from New Jersey to South Carolina for several warmer months (Castro 1993).
Tagging studies have shown the dusky shark’s range in the Northwest Atlantic to extend from New England south to the Caribbean Sea and the Gulf of Mexico no farther south than the Yucatan Peninsula (Kohler et al. 1998). These data support the designation of a distinct population segment designation for the dusky shark in the Northwestern Atlantic.
The dusky shark attains large sizes, reaching 360 cm Total Length (TL) (Castro 1993). In the western North Atlantic, males are estimated to reach sexual maturity at 231 cm Fork Length (FL) and 19 years of age, while females mature at 235 cm FL and 21 years of age (Natanson et al. 1995). The oldest reported dusky from vertebral centra aging studies is 37 years, but they are believed to live to a maximum age of 40 or 50 years (Natanson et al. 1995). Vertebral aging has been validated in Australian waters using OTC (oxytetracycline) marking and tag recapture methods (Simpfendorfer et al. 2002).
The dusky shark is viviparous, giving birth to litters ranging from 2–14 pups. Reproduction in this species is not well understood, and the length of the gestation period and time of mating have not been definitively determined. However, the lack of large yolky ova in late term females indicate at least a 1-year resting period following parturition (Musick 1995; Branstetter and Burgess 1996), and Branstetter and Burgess (1996) suggested that gestation might last for 22 to 24 months.
Dusky sharks suffered dramatic population decreases during the 1980s. Relative abundance of the species declined from 20% of the total shark catch on coastal long-lines to 1–2% of the catch (Musick et al. 1993). Fishing mortality and lack of a management plan (prior to 1993) combined with slow growth and a long reproductive process has led to their decline in Northwestern Atlantic waters. These factors combined with a shark fishery that expanded from 135 metric tons landed in 1979 to 6,452 metric tons in 1991 (Castro 1997) have been the main cause of the decline of the species. The dusky shark was one of the preferred species within this fishery along with blacktip (Carcharhinus limbatus), sandbar (C. plumbeus), and spinner sharks (C. brevipinna) (Scott 1997).
Demographic analyses have been conducted for many elasmobranch populations where sufficient catch, effort, or abundance estimates are not available to conduct in depth stock assessments. In these situations, demographic analyses may provide the best available description of the population being studied (Cailliet 1992; Simpfendorfer 1999b, 2005). Life-history table or Leslie matrix approaches have been utilized to provide guidance to managers or provide priors for more in depth dynamic assessment models. Cailliet et al. (1992) used results from tagging experiments on the Pacific angel shark, Squatina californica, coupled with demographic analyses to provide management suggestions for this fishery. Stage-based and age-based approaches have been used in various demographic analyses. Mollet and Cailliet (2002) suggested an age-based approach to enable incorporation of age-specific mortality estimates and also suggested utilization of Monte Carlo methods to estimate confidence intervals for model outputs (e.g. Cortés 2002).
Demographic analyses of dusky sharks have generated estimates of annual intrinsic rates of population increase, r, of 2.8% year−1 (Cortés 1998) and 5.6% year−1 (Sminkey 1996). These two estimates were based on a population free of fishing mortality and a 2-year reproductive cycle. The reality of heavy fishing mortality and the possibility of a 3-year reproductive cycle would lead to lower estimates of r, and the need for more conservative management strategies to protect this species.
Large coastal sharks were first protected in the Atlantic by a Secretarial Fishery Management Plan in 1993 (NMFS 1993). Subsequently, quotas were cut in 1997 after analyses indicated the original assessment had over-estimated the intrinsic rate of increase by 2.5 to 3 times. In 2000, dusky sharks were placed on the list of prohibited species (NMFS 2000), but by-catch mortality may remain a problem. The objectives of this study were: to estimate by-catch mortality of this species in the directed bottom longline fishery; to define reproductive periodicity and fecundity; and to incorporate this new information into a demographic model using stochastic techniques to better understand the response of the dusky shark population in the western North Atlantic to fishery mortality and related management requirements.
Materials and methods
Data collection
Data collected by the Florida Natural History Museum Commercial Shark Fishery Observer Program (CSFOP) from 1994–2000 in the South Atlantic and Florida region of the Gulf of Mexico were analyzed for mortality estimates and life history parameters. Observers collected data on bottom long-line sets and shark catches. Observer coverage was 35–40 days within each biannual commercial shark harvest season. Data on long-line sets included depth, soak times, number of hooks, and length of set. Biological data collected included species, fork length (FL), total length (TL), sex, hooking mortality, and reproductive status. Fork length and total length were measured from the tip of the snout to the fork of the tail and tip of snout to the tip of the caudal fin respectively. Individuals were given a reproductive category based on clasper characteristics in males and uterine condition in females. Males were designated as juvenile, sub-adult or mature. Measurements of clasper lengths on sub-adult and mature specimens were also recorded. Females were characterized as immature, maturing, mature, post-partum, or pregnant. Number of embryos and their minimum and maximum lengths were recorded for each litter. Average number of pups per female was calculated from 51 pregnant sharks. Reproductive status of 98 mature female dusky sharks was examined to determine the reproductive cycle of this species. Embryo sizes from CSFOP, the fishery independent longline survey conducted by the Virginia Institute of Marine Science (VIMS) (Musick et al. 1993), and the literature (Clark and von Schmidt 1965) coupled with reproductive states were used to determine gestation period.
Dusky shark data collected by VIMS from 1975–2000 were also analyzed for reproductive parameters. The VIMS longline shark survey operated in most years from May through October. Approximately seven longline sets were made each month at set stations along the Virginia coast. Longlines consisted of tarred nylon mainline and 100 steel yankee gangions (Musick et al. 1993). In total, 574 coastal sets were made consisting of 56,134 hooks and 2,451 soak hours.
Data analysis
Immediate hooking mortality of sharks was determined by the observers on haul-back. Post-release mortality was not determined in this study and remains unknown. For modelling purposes all live animals that were released were assumed to survive. Mortality estimates were calculated for each month and size class. Size classes for hooking mortality estimates were designated as <110 cm FL, 111–169 cm FL, 170–229 cm FL, 230–269 cm FL, and >270 cm FL. These size classes corresponded to neonates and juveniles, juveniles, sub-adults, adults, and large adults.
Correspondence analysis and multiple correspondence analyses were conducted using SAS software and the CORRESP procedure with the MCA statement to determine correlations factors associated with hooking mortality within the first shark fishing season. The second season was omitted from analyses due to the small numbers of dusky sharks landed during this time period.
Models
Age structured life tables that included probability distributions for demographic parameters were used to estimate population growth rates, generation times, and elasticities under various levels of exploitation following the methods of Cortés (2002) and Goldman (2002). Monte Carlo simulations and probability distributions were used to incorporate uncertainty in demographic parameters and to produce error estimates for these parameters.
Contributions of specific life-stages and ages to population growth were estimated using elasticity analyses (Heppell et al. 1999). Sensitivities of the models to perturbations in demographic parameters were also calculated. Life-tables were constructed for females using a yearly time step to produce estimates of population growth rate (λ) and intrinsic rate of increase (r, where \(\lambda = e^r \)), generation times (\(\overline A \)), net reproductive rate (R 0 ), reproductive value (v x ), and stable age distribution (c x ). Population doubling or halving time was also calculated as
Vital rates used in the model were those found in the literature and those produced by this study.
Uncertainty was included in the model to compensate for possible variation in reported life history parameters (Cortés 2002; Goldman 2002). Mills et al. (1999) suggested incorporating a range of values for life history parameters when conducting elasticity analysis combined with simulation techniques. This method also produces confidence intervals for output values such as population growth and population doubling time.
Probability distributions were created for the following parameters: maximum age (ω), proportion mature at age, fecundity (m x = number of female pups per female/length of reproductive cycle) and survivorship at age (S x ) (Table 1). The probability distribution for maximum age (ω) was centered on reported values. Natanson et al. (1995) used tag recapture data to produce a value of 45 years as maximum age (ω), thus 45 was given the greatest probability. Fecundity estimates used in the model were derived from this study. An extreme value probability distribution was fit to litter size frequency data from this study. A sex ratio of 1:1 within litters was assumed (McAuley et al. 2007). Female specific fecundity or m x was calculated as the product of the number of females per female per reproductive cycle, in this case 3 years, and the proportion of mature females at age calculated as
where a and b are regression descriptors. The estimate of b was held constant within the simulations. This maintained the average shape of the maturity ogive. A normal probability distribution was used to describe the a parameter.
Six methods were used for determining the range of survivorship for all age classes except the age-0 cohort in model simulations: Peterson and Wroblewski (1984), Hoenig (1983), Pauly (1980), Chen and Watanabe (1989), Jensen based on age at maturity (t mat ) and based on k (Jensen 1996). Probability distributions for survivorship were bounded by minimum and maximum values calculated from the 6 methods.
These methods did not address age-0 survivorship and little data exist for survivorship of young of the year elasmobranchs nor do they address the possibility of decreased survivorship at maturity due to reproductive costs. Gruber et al. (2001) estimated juvenile annual survival rates for lemon sharks, Negaprion brevirostris, to range from 38% to 65% under greater and lesser population densities respectively. Bounds for the probability distribution for age-0 survivorship were estimated by setting natural mortality to a value that would create a population growth rate of λ = 1 year−1 in a population free of fishing mortality (Vaughan and Saila 1976; Hoenig and Gruber 1990; Saether and Bakke 2000). The upper limit of this distribution was estimated by setting age-0 mortality to a value equal to the mean mortality estimated for the age-1 cohort to portray possible compensation within the fished population. A uniform probability distribution was then created and bounded by these two values. Given the lack of information for age-0 survival this appeared to be a viable method to estimate age-0 survivorships. Many K-selected species display disparity between age-0 survivorship and subsequent age classes (Olesiuk et al. 1990; Weimerskirch et al. 1992; Clapham et al. 2003). All other estimates for natural mortality were assumed to be within the correct range.
The effect of changes in age-0 survivorship on population growth rates was calculated using a Leslie matrix under a range of natural mortality estimates and levels of fishing mortality. Natural mortality estimates used ranged from 0.0–0.8 year−1. Fishing mortality was applied to all age classes equally and ranged from 0.0–0.5 year−1.
The net reproductive rate (R 0 ) was calculated as
Annual population growth rates (\(\lambda = e^r \)) were calculated from rates of population increase (r) by iteratively solving the Euler equation:
Generation time (\(\overline A \)) was calculated as
The distribution of reproductive values (v x ) was calculated as
where v o is the reproductive value at birth (v 0=1) and t represents all ages a female will attain through x to ω. The stable age distribution (c x ) was calculated as
Elasticities were calculated as
following the methods of Caswell (2001) and Cortés (2002). Elasticities were calculated for young of year or fertility, 1–3 age classes, 4–9 age classes, 10–20 age classes, 21–28 age classes and 29–50 age classes. The sensitivity of λ to changes in model parameters was calculated as
following the methods of Caswell (2001). Mean life expectancy was calculated as follows
P x is the mean survivorship of the probability distribution for age x. Monte Carlo simulation was used to randomly sample from probability distributions created for demographic parameters that possessed a level of uncertainty within the reported values. Confidence intervals were reported as the 2.5th and 97.5th percentiles after simulations had been run.
Three scenarios were established to examine the effects of fishing mortality. Scenario one assumed a population at equilibrium, and no fishing mortality was included in this scenario. Fishing mortality was included in scenarios two and three to estimate the effect of certain levels of fishing mortality on population growth and to simulate age specific hooking mortality induced by the commercial shark fishery in the winter fishing season (January to June). Recent stock assessments of the shark fishery for the large coastal complex estimated F to range from F = 0.07–0.21 year−1 (NMFS 2003). These estimates of fishing mortality were included in the models in scenarios two and three. In scenario two an annual mortality rate of 0.068% year−1 was divided by two and in scenario three 0.19% year−1 was divided by two. Annual mortality estimates were halved to compensate for F being applied to the first 6 months of the year. Age specific hooking mortality (expressed as percent landed that were dead) was estimated for sharks for which length and mortality data were available using von Bertalanffy growth parameters established by Natanson et al. (1995). A regression was fit to age-specific hooking mortality rates for ages 0–31. Little data were available for sharks greater than 31 years of age and thus they were excluded from the analyses. Mortality frequencies were also calculated for each Julian day. Annual mortality rates were then multiplied by percent dead for each age-class to establish age-specific fishery by-catch mortality rates. Ages were assigned using the age-growth relationship derived by Natanson et al. (1995) through vertebral analyses. Fishing mortality was applied only to ages 0–31 due to the lack of hooking mortality data for animals greater than 31 years of age.
Ratios of juvenile survival elasticity to age-zero survival elasticity were calculated to provide estimates of required compensatory responses following increased juvenile mortality following the methods of Heppell et al. (1999) and Cortés (2002).
Deterministic post-breeding projection matrices were used to display responses to conditions set in the models. An initial population of 1,000,000 animals was created following the stable age distribution calculated in scenario one. The population was projected over 100 years for each scenario beginning at equilibrium conditions.
Results
Hooking mortality
Hooking mortality increased as shark size decreased (Table 2, Fig. 1). Hooking mortality for dusky sharks less than 110 cm FL was 79%. Mature animals (>230 cm FL) experienced the lowest hooking mortality (37%). Soak time also influenced hooking mortality (Table 3). Mortality for North Carolina winter sets (January–April) was 79% when soak times were greater than 15 h. Mortality increased to 85% for soak times greater than 20 h. Mortality was 57% when soak times were less than 15 h. Soak times of less than 10 h had the lowest hooking mortality (5%).

Age specific catch frequency and age specific hooking mortality (r 2 = 0.53) for animals that data were available
Correspondence analysis of soak time with alive or dead observations yielded similar results. Longer soak times associated with dead sharks and the shorter soak times associated with live sharks. Spring months associated with the highest proportions of dead sharks. The smallest two size classes were also associated with dead sharks, while the largest size classes were located in the same quadrant as live sharks. The smallest size classes of dusky sharks closely corresponded to catches in North Carolina.
Reproduction
Females averaged 7.13 (n = 51) pups per litter and ranged from 3–12 pups (Fig. 2). Minimum and maximum values for litter sizes were 3 pups and 12 pups respectively. A minimum extreme probability distribution provided the best fit to the data. Litter sizes of 8 pups were given the highest probability within the models. Age at 50% maturity was estimated as 20 years of age (Fig. 3). Embryo size throughout the year indicated a gestation period of approximately 20 months (Fig. 4). Development appeared to last for 20–22 months following fertilization sometime between July and September and ending with birth between May and June. Time of mating is still not known and may not indicate time of fertilization due to the ability of this species to retain viable spermatozoa in the oviducal glands (Pratt 1993).

Frequency of litter sizes for dusky sharks from CSFOP data

Age-based maturity ogive. \({\text{Proportion mature}}_x = \frac{1}{{\left( {1 + e^{ - \left( { - 19.76 + 0.99x} \right)} } \right)}}\)

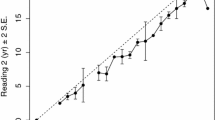
Minimum and maximum fork lengths of each litter over time from CSFOP, VIMS, and Clark and von Schmidt (1965)
CSFOP data on uterine condition show approximately 66% of mature females were pregnant from January through April (Fig. 5). Post-partum females accounted for 37% and 29% of the mature animals in July and September respectively. Pregnant females accounted for 21% and 29% of mature animals in July and September respectively. This suggests a resting period of a year following parturition and a 3-year reproductive cycle.

Monthly uterine condition of mature female dusky sharks from CSFOP
The largest immature female was 225 cm FL. The smallest mature female was 221 cm FL. All females greater than 226 cm FL were classified as mature. Springer (1940) reported a larger size at maturity (231 cm FL).
Intrinsic rates of population increase
Under zero fishing mortality and a P 0 value of 0.40, intrinsic rate of increase was 0% year−1. This translated to a natural mortality rate of 0.9 year−1. This was the greatest value for age-0 natural mortality that the population could sustain given the conditions established in the model. The greatest rate of population increase under zero fishing pressure attainable was 3.3% year−1. This assumed zero natural mortality at age-0 and a population free of fishing mortality.
Given the variability in parameters employed in the model under zero fishing pressure, intrinsic rates of population increase ranged from –1.9% and 6.7% year−1. The fishing scenarios investigated yielded negative rates of population increase for both fishing mortality scenarios (Table 4). Under the conditions of scenario two (F = 0.07 year−1), the intrinsic rate of population increase ranged from −6.0% to 7.6%. Under the conditions in the scenario three (F = 0.21 year−1) intrinsic rates of population increase ranged from −9.7% to 3.2% year−1.
Life expectancy
Mean life expectancy changed from 11.22 years in scenario 1 (F = 0) to 9.9 in the F = 0.21 scenario. Under fishing mortality conditions the probability of surviving to maturity was extremely low. Population halving time for Scenario 2 was 693 years. Population halving time for scenario 3 was 18 years.
Stable age distribution
Age-0 accounted for 15–17% of the population in all three scenarios. Contributions decreased with each increase in age. Assuming age at maturity is 21 and maximum age is 45, the female breeding population accounted for 10% of the total population and immature females accounted or 90% of the population.
Elasticities
The elasticity of λ to survival was greatest for the juvenile stage under all model scenarios. Elasticities for mature females decreased with age. The importance of the breeding population increased as fishing mortality increased and the contribution of younger age classes to population growth decreased (Table 4). Mean elasticities for juvenile survival and age-zero survival under conditions set in scenario one were 69.73 and 3.65 respectively. The mean elasticity ratio of juvenile survival to age-0 survival is 69.73/3.65 = 19.10. The two fishing mortality scenarios represent an average decrease in juvenile survival of 2.4% (scenario two) and 7.2% (scenario three) from scenario 1. Under the conditions in scenario two, age-0 survival or fecundity would have to increase by 46% to offset the decreased juvenile survival (Heppell et al. 1999; Cortés 2002). A 46% increase in fecundity translates to an increase of four pups from mean litter size (eight). The possibility of 12 pups per litter is reasonable and recovery may be possible. Scenario three represents a decrease in juvenile survival of 7.2%. Age-0 survival or fecundity would have to increase by 138%. This is not possible given mean values for fecundity and survivorship. An increase of 138% in fecundity translates to 19 pups per litter. An increase of 138% for survival and fecundity is greater than is physically possible.
Model sensitivity
Population growth rate was most sensitive to perturbations in survivorship of the youngest age classes, age at maturity and maximum age. As fishing mortality increased the model became less sensitive to age at maturity and maximum age. Sensitivity decreased as age increased for all juvenile ages.
Although the models were most sensitive to age-0 survival, when values for age-0 natural mortality were varied within the models, little change occurred in population growth rates. When age-zero natural mortality was completely removed from the model, population increase for scenario two increased to 1.2% (Fig. 6). When natural mortality was removed from the age-0 cohort in scenario three, rate of population increase was –2.6%.

Effects of natural mortality estimates on population growth rates under conditions set in scenarios
Deterministic matrix projections
The numerical projection of population size in scenarios two and three predicted large declines in population. Under scenario three the population decreased by 80% over 30 years and placed the population at extremely low levels in 100 years (Fig. 7). The conditions of scenario one project slight increases in population over 100 years.

Deterministic population projections for all three scenarios
Discussion
Few elasmobranch demographic models have accounted for fishery removals and their impact on population growth or the subsequent compensatory reactions of the population. Fishing mortality was incorporated in the models to investigate what levels of fishing mortality the population could withstand. Under minimal fishing pressure models returned negative values for population increase. Population growth was most sensitive to age-0 mortality in the fishing scenarios. When natural mortality of this age class was set at zero, population growth was 1.2% year−1 within scenario two (F = 0.07 year−1). Although population growth was most sensitive to age-0 mortality within the fishery scenarios, reducing natural mortality to zero did not compensate for fishing mortality in scenario three (F = 0.21 year−1) and moved population growth to slightly above zero in scenario two (F = 0.07 year−1).
Previous studies of the demography of the dusky shark have calculated r-values of 2.0% (Smith et al. 1998), 2.8% (Cortés 2002), and 5.57% (Sminkey 1996). Only one of these studies has accounted for the variability in natural mortality rates and none have included the effects of fishing mortality within the analysis (Cortés 2002). Most researchers have employed a single method for calculating natural mortality across all age classes. This approach does not account for differences in natural mortality as the species in question grows. A shark of 2 meters will likely have a lower natural mortality than a shark of lesser length (Peterson and Wroblewski 1984).
McAuley et al. (2007) estimated population growth rates for the dusky shark in Northwest Australia to be 2.5% year−1 for a population free of fishing mortality. Simpfendorfer (1999a) estimated a population increase of 4.2% year−1 in a population free of fishing. Our estimates are slightly lower (1.7%) due to the method of estimation for the age-0 survivorship and the assumption of a 3-year reproductive cycle. Simpfendorfer (1999a) suggested that fishing was sustainable as long as it was constrained to the youngest age classes. McAuley et al. (2007) found that even slight fishing mortality experienced by many age classes was not sustainable. These results are similar to ours and support our findings that fishing across many age classes may have severe repercussions.
Age-0 natural mortality
Survivorship of the age-0 cohort was established assuming the demographic parameters employed within the model represented a population experiencing fishing mortality. As such it is likely that compensatory mechanisms are in play. The probability distribution for age-0 mortality was bounded by values that would create a population at equilibrium and a population showing slight increases in population, possibly due to compensatory increased survival in the age-0 cohort. It is likely that survivorship could increase to the level of the age-1 cohort as a compensatory mechanism to offset exploitation (Gedamke et al. 2007). Various studies have found compensatory responses in elasmobranch populations following exploitation. Sminkey and Musick (1995) found significant changes in length at age of several age classes of juvenile sandbar sharks, Carcharhinus plumbeus, following exploitation. Carlson and Baremore (2002) reported decreases in length and age at maturity in the Atlantic sharpnose shark, Rizoprionodon terraenovae, following exploitation. Cassof et al. (2007) reported lower ages at maturity in the porbeagle, Lamna nasus, as a result of exploitation. Exploitation of the dusky shark was greatest in the early 1980s. Compensatory responses to fishing pressure are likely to be slow and would not be realized until 2000–2004 due to the late age at maturity and slow growth rates. Growth rate parameters employed in the models were derived in the mid 1990s and may not represent the current status of the population (Sminkey and Musick 1995). Given this possibility the models may have underestimated the population growth potential due to changes in growth rate and survivorship estimates over the past decade. The models did not account for the probable compensatory changes in growth rates and increased survivorship of the youngest age classes due to decreases in population densities (Gruber et al. 2001).
Heppell et al. (2000b) suggested utilizing demographic parameters of like species when demographic parameters for the study animal are difficult to determine or unavailable. Unfortunately, data for age-0 survivorship of elasmobranchs are scarce for many reasons. Few studies have attempted to estimate natural mortality through direct methods for coastal sharks. Simpfendorfer (1999b) estimated mortality rates of 0.561–0.698 year−1 through catch-curve analysis of the sharpnose shark in Australian waters. Natural mortality of young of the year blacktip sharks was estimated to be 0.52 year−1 using acoustic techniques within the nursery area (Heupel and Simpfendorfer 2002). Gruber et al. (2001) estimated juvenile annual survival rates for lemon sharks, Negaprion brevirostris, to range from 38% to 65% year−1 under greater and lesser population densities respectively.
Reproduction
Observers indicated on one occasion that a female released pups as the animal was being landed. Pups may be aborted by late term females following hooking and preceding landing, and as such the physical presence of pups may not be adequate for accurate determination of fecundity. Cliff et al. (1988) noted similar occurrences with sandbar sharks, Carcharhinus plumbeus, caught in protective gill-nets. Four pups were released from a term female upon extraction from the net. Placental scars were evident from the four released pups. Examination of placental scarring may provide a more accurate estimate of fecundity for late term females that may have released pups due to the stress of being hooked. This method was not employed within the present study. Although placental scaring was not used within this study, it is unlikely that our estimates of litter size were biased given the complete range of embryo lengths obtained within the study.
Springer (1940) recorded 5 litters in the month of January and March from Englewood, Florida. The average total length of the January and March litters was 80.5 cm and 60.0 cm, respectively. The former pups would have been pupped the following spring while the latter pups would have been pupped 12–14 months later.
Clark and von Schmidt (1965) found a similar phenomenon of two distinct groups of pups. One group of pups examined from August–March ranged from 21.6–69.9 cm FL. These pups would have been pupped in a year’s time. The second group of pups examined in December and January ranged from 85.5–96.5 cm FL. This group of pups would have been pupped the following spring. These litters averaged 7.7 pups and the sex ratio was approximately 1:1.
Elasticity and sensitivity analysis
The contribution of the juvenile stage to population growth is directly related to the length of the juvenile stage (Heppell et al. 2000a). Summing elasticities across age classes for each stage to estimate the importance of stages within the life history of species is biased when stages of disproportionate length are compared. Elasticity analysis within our models ranked age-0 and age-20 as being equally important to population growth. Sensitivity analysis appears to be a more appropriate method of estimating the importance of stages or age classes of long-lived species especially when a fished population that regularly experiences perturbations is involved. It follows that sensitivity analysis should be used for establishing management decisions when specific size ranges or age classes are in need of protection. Under fishing scenarios where multiple age classes were impacted, population growth was most sensitive to age-0 survival or fecundity.
Analyses of elasticity ratios illustrated possible compensatory responses for dusky sharks. The population could sustain the fishing mortality schedule set in scenario two if mean fecundity increased to 9 pups per litter. This is possible given the fecundity values previously reported and those values determined in the present study. Under the conditions in scenario three the population may not be able to compensate for fishing mortality unless management measures are taken to reduce juvenile mortality.
Conservation measures
Dusky sharks have slow growth and mature later than most other sharks (Natanson et al. 1995). In addition, they have a longer reproductive cycle than most other sharks. These characteristics result in a very low intrinsic rate of population increase and extreme vulnerability to over-fishing and population collapse. Demographic models suggest that even under modest levels of F, the dusky shark population may be on the edge of collapse. Better estimates of fishing mortality are needed to obtain a more accurate estimate of the effect of the commercial shark fishery on the population. Simpfendorfer (1999a) indicated that intrinsic rates of population increase would remain above zero if fishing mortality was restricted to the youngest year class. Although the greatest hooking mortality occurs in the youngest age classes, all year classes experience hooking mortality at some level.
Hooking mortality estimates used did not include possible post-release mortality of sharks classified as alive at time of landing. Some level of post-release mortality does occur and as such our estimates are likely to be below true levels and present a conservative estimate of population growth under the assumptions made.
The target for fishing mortality on the large coastal complex is F = 0.05–0.08 (NMFS 2003). If juvenile survival increases, the population may increase at very low levels of fishing mortality if fishing mortality is only experienced as hooking mortality. A directed fishery across all age classes is not sustainable even at the lowest levels of fishing mortality. Dusky sharks require greater protections from hooking mortality for the population to recover from intense fishing pressure.
References
Branstetter S, Burgess GH (1996) Commercial Shark Fishery Observer Program. Characterization and comparisons of the directed commercial shark fishery in the eastern Gulf of Mexico and off North Carolina through as observer program. MARFIN Award NA47ff0008, March 1996
Cailliet GM (1992) Demography of the central California population of leopard shark (Triakis semifasciata). Aust J Mar Freshwater Res 43:183–193 doi:10.1071/MF9920183
Cailliet GM, Mollet HF, Pittinger GG, Bedford D, Natanson LJ (1992) Growth and demography of the Pacific angel shark (Squatina californica), based upon tag returns off California. Aust J Mar Freshwater Res 43:1313–1330 doi:10.1071/MF9921313
Carlson JK, Baremore IE (2002) Changes in biological parameters of Atlantic sharpnose shark, Rhizoprionodon terraenovae, in the Gulf of Mexico: evidence for density-dependent regulation. Sci Counc Res Doc NAFO 2:8
Cassof RM, Campana SE, Myklevoll S (2007) Changes in the baseline growth and maturation parameters of the Northwest Atlantic porbeagle, Lamna nasus, following heavy exploitation. Can J Fish Aquat Sci 64:19–29
Castro JI (1993) The shark nursery of Bulls Bay, South Carolina, with a review of the shark nurseries of the southeastern coast of the United States. Environ Biol Fishes 38:37–48 doi:10.1007/BF00842902
Castro JI (1997) The limitations of shark fisheries and the shark fishery of the Southeastern United States. Collective volume of scientific papers. ICCAT 46:399–401
Caswell H (2001) Matrix population models:construction, analysis, and interpretation. Sinauer, Sunderland, Massachusetts, p 722
Chen SB, Watanabe S (1989) Age dependence of natural mortality coefficient in fish population dynamics. Nippon Suisan Gakkai Shi 55:205–208
Clapham P, Barlow J, Bessinger M, Cole T, Mattila D, Pace R, Palka D, Robbins J, Seton R (2003) Abundance and demographic parameters of humpback whales from the Gulf of Maine, and stock definition relative to the Scotian Shelf. J Cetacean Res Manag 5:13–22
Clark E, von Schmidt K (1965) Sharks of the Central Gulf coast of Florida. Bull Mar Sci 15:13–83
Cliff G, Dudley SFJ, Davis B (1988) Sharks caught in the protective gill nets off Natal, South Africa. 1. The sandbar shark Carcharhinus plumbeus (Nardo). S Afr J Mar Sci 7:255–256
Compagno LJV (1984) FAO species catalogues, vol 4. Sharks of the World. An annotated and illustrated catalogue of shark species known to date. FAO Fisheries Synopsis No. 125
Cortés E (1998) Demographic analysis as an aid in shark stock assessment and management. Fish Res 92:199–208 doi:10.1016/S0165-7836(98)00183-0
Cortés E (2002) Incorporating uncertainty into demographic modeling: application to shark populations and their conservation. Conserv Biol 16:1048–1062 doi:10.1046/j.1523-1739.2002.00423.x
Gedamke T, Hoenig JM, Musick JA, DuPaul WD, Gruber SH (2007) Using demographic models to determine intrinsic rate of increase and sustainable fishing for elasmobrnachs: pitfalls, advances, and appplications. N Am J Fish Manage 27:605–618 doi:10.1577/M05-157.1
Goldman KJ (2002) Aspects of age, growth, demographics and thermal biology of two lamniform shark species. Dissertation, The College of William & Mary, School of Marine Science, Virginia Institute of Marine Science
Gruber SH, de Marignac JRC, Hoenig JM (2001) Survival of juvenile lemon sharks at Bimini, Bahamas, estimated by mark-depletion experiments. Trans Am Fish Soc 130:376–384 doi:10.1577/1548-8659(2001)130<0376:SOJLSA>2.0.CO;2
Heppell SS, Crowder LB, Menzel TR (1999) Life table analysis of long-lived marine species with implications for conservation and management. In: Musick JA (ed) Life in the slow lane: ecology and conservation of long-lived marine animals. Symposium 23. American Fisheries Society, Bethesda, pp 137–148
Heppell S, Pfister C, de Kroon H (2000a) Elasticity analysis in population biology: methods and applications. Ecol 81:605–606
Heppell SS, Caswell H, Crowder LB (2000b) Life histories and elasticity patterns: perturbation analysis for species with minimal demographic data. Ecol 81:654–665
Heupel MR, Simpfendorfer CA (2002) Estimation of mortality of juvenile blacktip sharks, Carcharhinus limbatus, within a nursery area using telemetry data. Can J Fish Aquat Sci 59:624–632 doi:10.1139/f02-036
Hoenig JM (1983) Empirical use of longevity data to estimate mortality rates. Fish Bull (Wash D C) 82:898–903
Hoenig JM, Gruber SH (1990) Life-history patterns in the elasmobranchs: Implications for fisheries management In: Pratt JHL, Gruber SH, Taniuchi T (ed) Elasmobranchs as living resources: Advances in the biology, ecology, systematics, and the status of the fisheries, NOAA Tech. Rep. NMFS 90
Jensen AL (1996) Beverton and Holt life history invariants result from optimal trade-off of reproduction and survival. Can J Fish Aquat Sci 53:820–822 doi:10.1139/cjfas-53-4-820
Kohler NE, Casey JG, Turner PA (1998) NMFS Cooperative Shark Tagging Program, 1962–93: an atlas of shark tag and recapture data. Mar Fish Rev 60:1–87
McAuley RB, Simpfendorfer CA, Hall NG (2007) A method for evaluating the impacts of fishing mortality and stochastic influences on the demography of two-long-lived shark stocks. ICES J Mar Sci 64:1710–1722 doi:10.1093/icesjms/fsm146
Mills LS, Doak DF, Wisdom MJ (1999) Reliability of conservation actions based on elasticity analysis of matrix models. Conserv Biol 13:815–829 doi:10.1046/j.1523-1739.1999.98232.x
Mollet HF, Cailliet GM (2002) Comparative population demography of elasmobranchs using life history tables, Leslie matrices and stage-based matrix models. Mar Freshw Res 53:503–516 doi:10.1071/MF01083
Musick JA (1995) Critically endangered large coastal sharks, a case study: the sandbar shark, Carcharhinus plumbeus (Nardo, 1827). Shark News 5, October 1995: 6–7, illustr
Musick JA, Colvocoresses JA (1986) Seasonal recruitment of subtropical sharks in the Chesapeake Bight, USA. In: Yanez-Aracibia A, Pauley D (ed) Workshop on recruitment in tropical coastal demersal communities. FAO/UNESCO, Campeche Mexico. 21–25 April 1986. I.O.C. Workshop Report No. 44
Musick JA, Branstetter S, Colvocoresses JA (1993) Trends in shark abundance from 1974 to 1991 for the Chesapeake Bight region of the U.S. Mid-Atlantic coast NOAA. Tech Rep NMFS 115:1–18 pp. 115
Natanson LJ, Casey JG, Kohler NE (1995) Age and growth estimates of the dusky shark, Carcharhinus obscurus, in the western North Atlantic Ocean. Fish Bull (Wash D C) 93:116–126
NMFS (1993) Fishery management plan for sharks of the Atlantic Ocean. NOAA/NMFS, U.S. Department of Commerce Department of Commerce, Feb. 25, 1993
NMFS (2000) Fisheries of the United States. NOAA/NMFS, U.S. Department of Commerce, October 2000
NMFS (2003) Stock Assessment and Fishery Evaluation Report for Atlantic Highly Migratory Species. NOAA/NMFS, U.S. Department of Commerce. January 2003
Olesiuk PF, Bigg MA, Ellis GM (1990) Life history and population dynamics of resident killer whales (Orcinus orca) in the coastal water of British Columbia and Washington State In: Hammond PS, Mizroch SA, Donovan GP (ed) Individual recognition of cetaceans: Use of photo-identification and other techniques to estimate population parameters. Incorporating the proceedings of the symposium and workshop on individual recognition and the estimation of cetacean population parameters., Rep Int Whaling Comm (Spec. Issue), pp 209–243
Pauly D (1980) On the interrelationships between natural mortality, growth parameters, and mean environmental temperature in 175 fish stocks. J Conseil Int Explor Mer 39:175–192
Peterson I, Wroblewski JS (1984) Mortality rate of fishes in the pelagic ecosystem. Can J Fish Aquat Sci 41:1117–1120 doi:10.1139/f84-131
Pratt HL Jr (1993) The storage of spermatozoa in the oviducal glands of western North Atlantic sharks. Env Bio Fish 38:139-149
Saether B-E, Bakke O (2000) Avian life history variation and contribution of demographic traits to the population growth rate. Ecol 8:642–653
Scott GP (1997) Recent trends in catch rates of some Atlantic sharks. Collective Volume of Scientific Papers. ICCAT 46:409–414
Simpfendorfer C (1999a) Demographic analysis of the dusky shark in southwestern Australia. In: Musick JA (ed) Life in the slow lane: Ecology and conservation of long-lived marine animals. American Fisheries Society, Bethesda, pp 149–160
Simpfendorfer C (1999b) Mortality estimates and demogrpahic analysis for the Australian sharpnose shark, Rhizoprionodon taylori, from northern Australia. Fish Bull (Wash D C) 97:978–986
Simpfendorfer CA (2005) Demographic models: life tables, matrix models and rebound potential. In: Musick JA, Bonfil R (ed) Elasmobranch Fisheries Management Techniques, FAO Fisheries Technical Paper, Rome, pp 187–204
Simpfendorfer CA, McAuley RB, Chidlow J, Unsworth P (2002) Validated age and growth of the dusky shark, Carcharhinus obscurus, from Western Australian waters. Mar Freshw Res 53:567–573 doi:10.1071/MF01131
Sminkey TR (1996) Demographic analyses of natural and exploited populations of three large coastal sharks. Document SB-III-8 of the 1996 Report of the Shark Evaluation Workshop. June 1996. Southeast Fisheries Science Center, Miami, FL, p 5
Sminkey TR, Musick JA (1995) Age and growth of the sandbar shark, Carcharhinus plumbeus, before and after population depletion. Copeia 4:871–883 doi:10.2307/1447035
Smith SE, Au DW, Show C (1998) Intrinsic rebound potentials of 26 species of Pacific sharks. Mar Freshw Res 49:663–678 doi:10.1071/MF97135
Springer S (1940) The sex ratio and distribution of some Florida sharks. Copeia 3:188–194 doi:10.2307/1437982
Vaughan DS, Saila SB (1976) A method for determining mortality rates using the Leslie matrix. Trans Am Fish Soc 105:380–383 doi:10.1577/1548-8659(1976)105<380:AMFDMR>2.0.CO;2
Weimerskirch H, Stahl J, Jouventin P (1992) The breeding biology and population dynamics of king penguins Aptenodytes patagonica on the Crozet Islands. Ibis 134:107–117 doi:10.1111/j.1474-919X.1992.tb08387.x
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Romine, J.G., Musick, J.A. & Burgess, G.H. Demographic analyses of the dusky shark, Carcharhinus obscurus, in the Northwest Atlantic incorporating hooking mortality estimates and revised reproductive parameters. Environ Biol Fish 84, 277–289 (2009). https://doi.org/10.1007/s10641-008-9435-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-008-9435-6