
Abstract
Ulcerative colitis (UC) is difficult to be treated. miRNAs are a group of gene regulators. Study demonstrated that miR-200b-3p is involved in the development of UC, but the specific molecular mechanism is still unclear. A UC model was established by injecting acetic acid into rectum of rats, which were then treated with miR-200b-3p antagonists and agonists. Weight change, fecal viscosity and fecal bleeding were measured to determine disease activity index. The ratio of colon length to weight was measured. Colon lesions were detected by H&E staining. ELISA was used to detect the expression of TGF-β in colon tissues and IL-10/CRP in serum. Intestinal epithelial cells (NCM460) were treated by TNF-α to create an inflammatory environment. MRNA and protein levels of miR-200b-3p, KHDRBS1, IL-10, IL-6, IL-1β, TGF-β, Bcl-2, Bax and C-capase-3 were detected by qRT-PCR and Western blot, respectively. TargetScan database and dual-luciferase reporter assay were conducted to predict the targeting relationship between miR-200b-3p and KHDRBS1. MTT and flow cytometry were respectively performed to detect cell proliferation and apoptosis. MiR-200b-3p expression was inhibited, leading to increased disease activity index and colonic length–weight ratio, and aggravation of lesions of the UC rat model. Up-regulation of miR-200b-3p can relieve inflammation and apoptosis of immune cells in UC rats. MiR-200b-3p targeted KHDRBS1 and inhibited its expression. Moreover, KHDRBS1 reversed the effects of miR-200b-3p on apoptosis, proliferation and inflammation of intestinal epithelial cells. MiR-200b-3p alleviates UC by negatively regulating KHDRBS1.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Ulcerative colitis (UC) is an inflammatory bowel disease (IBD) caused by a combination of genetic, environmental and immune factors (Khor et al. 2011; Strober and Fuss 2011). Typical symptoms of UC are bloody diarrhea, abdominal pain, hematochezia, and weight loss (Conrad et al. 2014; Xavier and Podolsky 2007). At present, the main therapeutic drugs are salicylic acid preparations, and surgical treatment is considered for patients with severe UC (Purrmann and Strohmeyer 1989). Intestinal epithelial cells (IECs) are the main functional cells in the intestinal tract and the structural basis of the intestinal epithelial barrier (Vlantis et al. 2016; Zhang et al. 2019). Some studies proved that intestinal epithelial barrier dysfunction is the main pathogenic factor leading to UC (Stone 2012). Therefore, studying IECs could help understand the pathogenesis of UC. Current research results showed that miRNAs affect intestinal epithelial cell proliferation, apoptosis and inflammatory response, and are closely related to the occurrence and development of UC (McKenna et al. 2010; Nguyen et al. 2010; Peck et al. 2017). However, the specific molecular mechanism through which miRNA affected the UC process was unclear.
MiRNAs are a group of short (20–24 nt) non-coding RNAs that are involved in the regulation of genes (Xie et al. 2005). In recent years, miRNAs have been found to be involved in the regulation of cell apoptosis and inflammation. In particular, some miRNAs, such as miR-200 family, accelerate disease progression, showing a strong potential to serve as biomarkers (Fabbri et al. 2012). Some studies have found that miR-200 combines with Homeobox 1 and 2 (ZEB1 and ZEB2) through zinc finger E-box to form miR-200/ZEB1 or miR-200/ZEB2 axis, regulating biological functions such as cellular inflammatory response and apoptosis(Noman et al. 2017). The miR-200 family consists of miR-200a, miR-200b, miR-200c, miR-429 and miR-141. Chen et al. suggested that miR-200b could maintain intestinal epithelial integrity by inhibiting EMT and promoting proliferation of IECs (Chen et al. 2013).
KH RNA binding domain containing, signal transduction associated protein 1 (KHDRBS1) also refers to Sam68, which is a kind of protein-coding gene, participates in alternative splicing, cell cycle regulation, gene expression and regulation of human immunodeficiency virus (HIV), and other cellular processes (Lukong and Richard 2003). KHDRBS1 promotes apoptosis by destroying the protective effect of IEC through NF-κB signaling pathway (Qian et al. 2016). Both miR-200b-3p and KHDRBS1 can influence the function of IEC, and we speculated whether there was a regulatory relationship between them.
The purpose of this study was to explore the roles of miR-200b-3p and KHDRBS1 in the development of UC, and the regulatory relationship between them.
Methods and materials
Ethics statement
All animal experiments were conducted following the guidelines of the Chinese Council for Animal Care and Use. This study was approved by the Experimental Animal Committee of Huai’an TCM Hospital Affiliated to Nanjing University of Chinese Medicine. Efforts have been made to minimize pain and discomfort to the animals during the experiments. The animal experiments were conducted in Huai’an TCM Hospital Affiliated to Nanjing University of Chinese Medicine.
Model mice with ulcerative colitis
A total of 60 male Wistar rats (6 weeks) weighing 200 ± 20 g were used in this study. The animals were obtained from the Nanjing Institute of Model Zoology (China). All the rats were raised in polypropylene cages and fiven free access to food (23% protein and 4% fat) and water. The laboratory temperature was maintained at 21 °C, with humidity of 45–55% and day-night light cycle of 12 h. Domestication began one week before the experiment and the animals were fasted for 24 h before the experiment. For modeling, 50 rats were given abdominal anesthesia with 50 mg/kg ketamine and fasted overnight (they were able to drink water freely). A flexible 6F polypropylene catheter lubricated with K-Y glue was inserted into the rat colon 6–8 cm in to the anal canal. 2 ml of acetic acid (AA) (3% by volume/volume in 0.9% saline) was slowly injected into the distal colon. The animals remained in the Trendelenburg supine position for 30 s to prevent the solution from draining.
Both the agonist and antagonist of miR-200b-3p were purchased from RIBOBIO Biotech (China). The rats were randomly divided into 6 groups (10 rats/ group) and treated as follows: Group 1 (control group): untreated rats; Group 2 (Model group): UC rats; Group 3 (Model+agomiR-NC group): UC rats were injected with 5 nmol miR-200b-3p agomiR negative control for 3 days; Group 4 (Model+agomiR group): UC rats were injected with 5 nmol miR-200b-3p agomiR for 3 days; Group 5 (Model+antagomiR-NC group): UC rats were injected with 10 nmol miR-200b-3p antagomiR negative control for 3 days; Group 6 (Model+antagomiR-NC group): UC rats were injected with 10 nmol miR-200b-3p antagomiR for 3 days. The rats were cultured for another 24 h under laboratory conditions and then sacrificed by cervical dislocation.
Disease activity index (DAI) and colon weight/ length ratio
The DAI could reflect the severity of the disease. The main indicators are scores of weight loss percentage, fecal viscosity and fecal bleeding specifically as follows: weight loss percentage (0, none; 1, 1–5%; 2, 5–10%; 3, 10–15%; 4, > 15%), defecate viscosity (0, nomal; 2, loose stool; 4, diarrhea), and defecate haemorrhage (0, normal; 2, slight bleeding; 4, overt bleeding). DAI value is equal to the mean of the three scores. After sacrifice, the rat colons were removed, and the weight and length of the colon were measured.
H&E staining
The colon of each animal was removed from the anal margin to the colonic cecum junction, then fixed with 10% neutral buffered formalin, and paraffin-embedded. The colon tissue was cut into 5 μm sections. The slices were rinsed in xylene for 10 min twice, and then in 95% alcohol for 3 min twice, followed by rinsing in 80% alcohol for 1 min. Next, the slices were then washed by distilled water for 1 min, stained with sappanwood semen (C0105, Beyotime Biotechnology, China) for 10 min, rinsed in running water for 3 s (s), then treated by 1% hydrochloric acid glycol for 3 s, washed with running water for 10 s, and then rinsed in running water for 10 min. Subsequently, the slices were stained with Eosin (0.5%) (C0105, Beyotime Biotechnology, China) for 2 min, and rinsed in distilled water for 2 s, then in 80% glycol for 3 s. The slices were added with 95% alcohol for 3 min twice, and treated by anhydrous ethanol for 5 min twice, then transferred to xylene for 5 min twice. Finally, the slices were sealed by neutral gum. The cytoplasm and nucleus were red and blue-purple, respectively.
ELISA assay
ELISA assay kits (TGF-β: ab119558, abcam, USA; IL-10: ab214566, abcam, USA; CRP: ab256398, abcam, USA) were used to detect the content of TGF-β in colon tissues of rats and the content of IL-10 and CRP in blood of rats. Colon tissues of rats were rinsed with pre-cooled saline, dried by filter paper and weighed. 0.86% pre-cooled normal saline was added to the tissues, which were then fragmented by a tissue homogenizer into 10% tissue fluid at 10,000 rpm/min. The tissue fluid or serum was then transferred to a centrifuge tube for later use. The experiment was performed at room temperature (20–25 °C) for 30 min. Six concentrations were used in corresponding groups, and blank group and sample group for testing were also set. Briefly, 100 μL of standard solution was added into blank micropores, while 100 μL of distilled water was added to the blank control holes. The remaining micropores were added with 100 μL samples for testsing. Then 50 μL of enzyme-labeled solution was added to each hole of the standard group and the sample group. Next, the label plate was sealed with seal paper and incubated in a wet box at 37 °C for 1 h. After incubation, each well was filled with diluted washing solution, then stood for 15–30 s, the enzyme label plate was fully cleaned for 5 times, and the water absorbent paper was thoroughly patted dry. 50 μL of chromogenic solution A and 50 μL of chromogenic solution B was added to each hole for reaction at 25–37 °C in the dark for 10–15 min. 50 μL termination liquid was added to stop the reaction. Finally, OD value of each hole at a wavelength of 450 nm was measured with a microplate reader (Molecular Devices, Shanghai, China).
TargetScan database
TargetScan 7.2 (http://www.targetscan.org/vert_72/) was used to predict targeted binding genes of miR-200b-3p to select an appropriate candidate gene.
Dual-luciferase reporter gene experiment
The 3′-UTR sequences of rat whole-genome transcripts was obtained from National Center for Biotechnology Information (https://www.ncbi.nlm.nih.gov/). The 3′-UTR of KHDRBS1 with highly conserved binding sites of miR-200b-3p including the sequence of binding sites was 5′-GGGCCCTAATAATTAAATTTTCAGTTATTTCTCGAC-3′. The 3'UTR sequence was inserted into the pcDNA 3.1 vector [P0157, Miaolingbio, China (http://www.miaolingbio.com/)] by ApaIand XhoIdouble digestion to construct the recombinant double-luciferase reporter vector pcDNA 3.1. Meanwhile, the core sequence of miR-200b-3p binding sites was mutated through the DNA synthesis to generate plasmid pcDNA 3.1-Mut containing mutant KHDRBS1 3′UTR (5′-GGGCCCTAATAATTAAATTTTAGUCGCGTCTCGAC-3′). MiR-200b-3p sequence (5′-UAAUACUGCCUGGUAAUGAUGA-3′) was constructed into pCMV-miR vector to form miR-200b-3p overexpressed plasmid. NCM460 cells (ATCC, Shanghai, China) were inoculated into a 12-well plate at a density of 50% and cultured in an incubator with 5% CO2 at 37 °C for 16 h. When cell fusion degree reached about 70%, the luciferase reporter plasmid was transfected. Six co-transfection groups were divided as follows: KHDRBS1-3′UTR-NC+miR-200b-3p group; KHDRBS1-3′UTR-NC+mirRNA-NC group; KHDRBS1-3′UTR-WT+miR-200-b-3p group; KHDRBS1-3′UTR-WT+mirRNA-NC group; KHDRBS1-3′UTR-MU+miR-200-b-3p group; KHDRBS1-3′UTR-MU+mirRNA-NC group. 1 μg of 3′UTR dual-luciferase plasmid and 0.4 μg of miRNA plasmid were transfected into the paired cells, respectively. After transfection for 6 h, the culture medium was changed and then the cells were continued to be incubated with 5% CO2 at 37 °C. Luciferase activity was measured by Dual Luciferase ® Reporter Assay system (Molecular Devices) for 48 h after transfection.
Cell culture and treatment
Intestinal epithelial cell line NCM460 was cultured in DMEM medium containing 90% DMEM (high glucose) (12,491-015, Gibco, USA, 10%FBS 10,099-141, Gibco, USA), 100 IU/ml streptomycin and 100 IU/ml penicillin (15070063, Gibco, USA) in an incubator with 5% CO2 at 37 °C. The cells were divided into 7 groups for treatment as follows: blank group (normal NCM469 cells), TNF-α group (treatment with 0.5 ng/ mL TNF-α (HY-P1860, MCE, China) at 37 °C), TNF-α+MC group (treatment with 4 μg of miR-200b-3p mimic control plasmid at 37 °C on the basis of the TNF-α group), TNF-α+M group (treatment with 4 μg of miR-200b-3p mimic plasmid at 37 °C on the basis of the TNF-α group), TNF-α+NC group (treatment with 4 μg of KHDRBS1 negative control plasmid at 37 °C on the basis of the TNF-α group), TNF-α + KHDRBS1 group (treatment with 4 μg of KHDRBS1 plasmid at 37 °C on the basis of the TNF-α group), and TNF-α+M+KHDRBS1 group (treatment with 4 μg of miR-200b-3p mimic plasmid and 4 μg of KHDRBS1 plasmid at 37 °C on the basis of the TNF-α group).
Cell transfection
The sequences of miR-200b-3p mimics were 5′-TAATACTGCCTGGTAATGATGAC-3′ and 5′-UCAUCACCACCAGGCAGUAUUA-3′ (antisence). KHDRBS1 plasmid was constructed as above. Lipofectamine 2000 (lipo2000) (11,668-027, Invitrogen, USA) was used to transfect the plasmid. Follow the instructions for transfection procedures, simply, NCM460 cells were planted in a 6-well plate with 2 × 105 cells/well and cultured in a low-serum medium. Transfection began the next day when the cell fusion rate reached above 90%. 50 μL Opti-MEM medium (31,985-070, Gibco, USA) was added to A and B centrifuge tube. 10 μL lipo2000 was added to tube A and 4 μg plasmid (miR-200b-3p mimic plasmid and KHDRBS1 plasmid) was added to tube B, and then let stand for 5 min. Subsequently, tube A and tube B was mixed and let stand for 20 min. Add into the corresponding cell hole for further culture. After 6 h, the culture medium was replaced with normal medium and continued in the incubator with 5% CO2 at 37 °C.
qRT-PCR
Colons of rats or IECs were collected. Total RNA of tissues/cells was extracted with TRIzol (15,596-018, Invitrogen, USA), and miRNA of tissues/cells was extracted with MiRcute miRNA isolation kit (DP501, Tiangen, China). All RNA and miRNA were quantified by enzyme marker (Molecular Devices). Then 1 μg of RNA was reverse-transcribed using the first cDNA synthesis kit (RR047AA, TaKaRa, Japan). The 20 μmol/L system for qRT-PCR consisted of 2 μL cDNA, 10 μL SYBR Premix Ex TaqII™ (2 ×) (RR036A, TaKaRa, Japan), and 0.2 mol/L for each primer. Finally, the system was filled with sterile distilled water. ABI 7500 sequencing detection system (Biosystems, Foster City, USA) was used for qRT-PCR amplification of cDNA. The reaction conditions were: 95 °C 30 s; followed 40 cycles at 95 °C 30 s, at 60 °C 3 s. Primers of Bax, Bcl-2, KHDRBS1, TGF-β, IL-10, IL-1, IL-6, miR-200b-3p, U6 and GAPDH in rat or human were shown in Table 1. The mRNA expressions of Bax, Bcl-2, KHDRBS1, TGF-β, IL-10, IL-1 and IL-6 were normalized to that of GAPDH, while miR-200b-3p mRNA expression was normalized to that of U6.
Western blot
The RIPA (P0013B, Beyotime Biotechnology, China) lysis buffer containing protease inhibitor (P1030, Beyotime Biotechnology, China) and phosphatase inhibitor (P1081, Beyotime Biotechnology, China) was added to the cells or tissues, and the whole cell/ tissues extract was obtained after lysis at 4 °C for 30 min. BCA protein quantitative kit (P0012, Beyotime Biotechnology, China) was used to quantify the protein concentration of each sample. The protein was purified by sodium dodecyl sulfate/polyacrylamide gel electrophoresis (P0012A, Beyotime Biotechnology, China), and then transferred to polyvinylidene fluoride membrane (PVFD) (ISEQ00010/IPVH00010, MILLIPORE, USA). The maker (PR1910/PR1920, Solarbio, China) was used to mark the molecular weight of the protein. The membrane was blocked by 5% skimmed milk (232,100, BD, USA) for 1 h, cleaned by TBST for 5 min, and then reacted with primary antibodies (Bcl-2: ab59348, 1:800, abcam; Bax: ab32503, 1:5000, abcam; C-caspase-3: ab49822, 1:500, abcam; KHDRBS1: ab76471, 1:20,000, abcam; IL-6: ab9770, 1:500, abcam; IL-10: ab33471, 1:1500, abcam; TGF-β: ab179695, 1:1000, abcam; IL-1β: ab205924, 1:1000, abcam; GAPDH: ab181602, 1:10,000, abcam) and incubated at 4 °C overnight. On the second day, the membrane was taken out and primary antibodies were recovered. Then TBST was cleaned for 4 times 5 min each time. The cleaned membrane was then incubated in secondary antibodies (Goat Anti-Rabbit IgG H&L: ab6721, 1:10,000, abcam; Goat Anti-Rat IgG H&L: ab97057, 1:5000, abcam) for 1 h. At the end of incubation, TBST was used for washing for 6 times 5 min for each time. Finally, the enhanced chemiluminescence Advanced System (Bio-Rad, CA, USA) was used to visualize the membrane by adding the enhanced chemiluminescence solution (WBKLS0500, MILLIPORE, USA). Development results were analyzed by Image J (1.8.0, National Institutes of Health, Germany).
MTT assay
Cell proliferation was measured by MTT (ST316, Beyotime Biotechnology, China). In short, IECs were first seeded into 6-well plates at 5000 cells/well for 24 h. Then, TNF-α, and plasmid were separately added into each group of cells as previously described. The cells were then added with MTT (20 μL 5 mg/ ml) and incubated at 37 °C for 4 h. Next, the supernatant was removed and DMSO (100 μL/well) was added to the cells. Finally, absorbance at 570 nm was measured with a microplate reader (Molecular Devices, Shanghai, China).
Flow cytometry
Cell apoptosis was detected by flow cytometry and the cells were processed using the Annexin V-FITC Apoptosis Kit (556,547, Becton,Dickinson and Company, USA), according to the manufacturer’s instruction. Briefly, the cells were collected in a centrifuge tube and added with 500 μL of pre-cooled 1 × PBS (4 °C). The cells were resuspended twice and centrifuged at 2000 rpm/ min for 5–10 min. After discarding the supernatant, the cells were resuspended in 100 μL 1 × Binding Buffer and then transferred to a test tube. Subsequently, Annexin V-FITC (5 μL) and PI (5 μL) were added to the tube at room temperature and in the dark for 30 min. After the cells were added with 400 μL of 1 × Binding Buffer, and another resuspension, the cells apoptosis was finally detected with a CytoFLEX LX Flow cytometer (Beckman Coulter, CA, USA).
Data analysis
The results were expressed as mean ± SD. Differences among multiple groups were compared using one-way ANOVA, and then Tukey's test was performed. Kruskal–wallis nonparametric test and Nemenyi test were used to analyze data with skewed distribution or variance heterogeneity. P < 0.05 was considered as statistically significant. SPSS.19.0 (IBM, NY, USA) was used for data analysis.
Results
Effects of miR-200b-3p on lesions and inflammation in UC rats
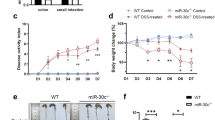
In order to explore the expression of miR-200b-3p in UC, we established a model of UC by injecting AA into rectum of rats (Saber et al. 2019). As shown in Fig. 1A, the expression of miR-200b-3p in the UC model group was significantly lower than that in the control group (P < 0.001). In addition, we injected the antagonists and agonist of miR-200b-3p in vivo through systematic administration. The agonist up-regulated the expression of miR-200b-3p in colon tissues of rats with UC, while antagonists downregulated the expression of miR-200b-3p (Fig. 1A).

The expression of miR-200b-3p was down-regulated in UC rats. A The expression of miR-200b-3p in each group was detected by qRT-PCR. (n = 10, * vs. Control; # vs. Model+agomiR-NC; ^ vs. Model+antagomiR-NC. ^P < 0.05; ***P < 0.001; ###P < 0.001). B Effect of miR-200b-3p on disease activity index (DAI) in each group rats. (n = 10, * vs. Control; # vs. Model+agomiR-NC; ^ vs. Model+antagomiR-NC. ^^^P < 0.001; ***P < 0.001; ###P < 0.001). C Colon weight/length ratio in each group rats. (n = 10, * vs. Control; # vs. Model+agomiR-NC; ^ vs. Model+antagomiR-NC. ^^P < 0.01; ***P < 0.001; ##P < 0.01). D H&E staining identified the effect of miR-200b-3p on the pathological status of colon in each group rats. E and F ELISA was used to detect the effect of miR-200b-3p on the expressions of related inflammatory factors in each group rats. (n = 10, * vs. Control; # vs. Model+agomiR-NC; ^ vs. Model+antagomiR-NC. ^^P < 0.01; ^^^P < 0.001; ***P < 0.001; ##P < 0.01; ###P < 0.001). Group: Control group; Model group; Model+agomiR-NC group; Model+agomiR group; Model+antagomiR-NC group; Model+antagomiR group. qRT-PCR quantitative reverse transcription polymerase chain reaction, H&E hematoxylin and eosin staing, ELISA enzyme-linked immuno sorbent assay, TGF-β transforming growth factor beta, IL-10 interleukin 10, CRP, C-reactive protein
DAI score was used to assess the effect of miR-200b-3p on the disease activity index of rats. The disease activity index of UC rat model was significantly increased, as compared with the control group. miR-200b-3p agonist could effectively reduce the severity of UC (P < 0.001), as compared with the UC group. However, miR-200b-3p antagonists significantly increased the severity of UC (P < 0.001) (Fig. 1B).
As can be seen from Fig. 1C, AA in the UC group led to a significant increase in colon weight/length ratio compared with normal animals (P < 0.001). MiR-200b-3p agonist significantly offset the increase of colon weight/length ratio (P < 0.01), as compared with the UC group. However, miR-200b-3p antagonists further promoted the increase of colon length–weight ratio (P < 0.01), as compared with the UC group.
The results of H&E staining showed that in addition to bleeding, edema and severe neutrophil infiltration (deep to muscle layer), the rats in the UC group also showed the characteristics of hemorrhagic ulcerative enteritis, accompanied by extensive intestinal crypt necrosis. The rats with treatment of miR-200b-3p agonist showed a marked reduction of lesions. As expected, miR-200b-3p antagonists significantly increased lesions in UC rats (Fig. 1D).
TGF-β, IL-10, and CRP are all involved in inflammatory response. The previous experiment demonstrated that TGF-β and CRP expression is increased and IL-10 expression is decreased under inflammation (Motoyama et al. 2009). TGF-β in rat tissues and IL-10 and CRP in serum were detected by ELISA, and the results showed that TGF-β and CRP levels were increased and IL-10 level was decreased in UC rats, suggesting the occurrence of inflammation. In addition, up-regulation of miR-200b-3p reduced TGF-β expression and increased CRP and IL-10 levels. The down-regulation of miR-200b-3p showed opposite results (Fig. 1E–G).
MiR-200b-3p affected expressions of apoptotic markers in UC rats
Bcl-2 and Bax are two of the most important regulatory genes with opposite functions during apoptosis (Yang et al. 1995). C-caspase-3 is the most critical apoptotic executive protease in the process of apoptosis (Qiu et al. 2000). We detected the protein expression levels of Bcl-2, Bax and C-Caspase-3, and found that the expressions of Bcl-2 and Bax proteins were consistent with their mRNA expressions. The protein expression level of C-caspase-3 was significantly decreased in UC rats as compared to the control group. Thus, miR-200b-3p agonist could reduce the expression of C-caspase-3, and that miR-200b-3p antagonists could further increase the expression of C-caspase-3 (Fig. 2A–D). We also detected the mRNA expression levels of Bcl-2 and Bax in the colon tissues of UC rats. As shown in Fig. 2E and F, the mRNA expression level of Bcl-2 was significantly decreased in UC group, as compared with the control group. MiR-200b-3p agonist could increase the expression of Bcl-2 and decrease the expression of Bax, but the effect of miR-200b-3p antagonist was the opposite.

MiR-200b-3p promoted the expressions of apoptosis-related proteins in UC rats. A Western blot analysis of C-Caspase-3, Bcl-2 and Bax expressions in each group rats. B–D The signal intensities of C-Caspase, Bcl-2 and Bax were determined. (n = 10, * vs. Control; # vs. Model+agomiR-NC; ^ vs. Model+antagomiR-NC. ^P < 0.05; ^^P < 0.01; ***P < 0.001; ##P < 0.01). E and F qRT-PCR analysis of Bcl-2 and Bax expression in each group rats. (n = 10, * vs. Control; # vs. Model+agomiR-NC; ^ vs. Model+antagomiR-NC. ^P < 0.05; ^^^P < 0.001; ***P < 0.001; ##P < 0.01; ###P < 0.001). Group: Control group; Model group; Model+agomiR-NC group; Model+agomiR group; Model+antagomiR-NC group; Model+antagomiR group. Bcl-2 B cell lymphoma/lewkmia-2, Bax Bcl-2 associated X protein
MiR-200b-3p negatively regulated KHDRBS1
Through TargetScan Human 7.2 (http://www.targetscan.org/vert_72/), we predicted the binding target of miR-200b-3p (Fig. 3A). KHDRBS1 was selected as the candidate molecule in combination with the score. Dual-luciferase reporter assay was used to further verify the targeted binding of miR-200b-3p to KHDRBS1, and the fluorescence intensity of KHDRBS1-MUT was significantly higher than that of KHDRBS1-WT (P < 0.001) (Fig. 3B). By further detecting the mRNA and protein expression of KHDRBS1 after up-regulation or down-regulation of miR-200b-3p, we detected a regulatory relationship between miR-200b-3p and KHDRBS1. The mRNA and protein expression levels of KHDRBS1 were increased after the down-regulation of miR-200b-3p, as shown by qRT-PCR and Western blot. When miR-200b-3p was up-regulated, the results were reversed (Fig. 3C–E). Figure 4A and B showed that TNF-α inhibited the expression of miR-200b-3p and promoted the expression of KHDRBS1 (P < 0.001). In addition, over-expressing both miR-200b-3p and KHDRBS1 significantly decreased KHDRBS1 expression, as compared with over-expression of KHDRBS1 (P < 0.001). Further detection of the protein expression of KHDRBS1 in each group showed consistent result with the mRNA expression, suggesting a negative regulatory effect of miR-200b-3p on KHDRBS1 (Fig. 4C, D). The above results indicated that miR-200b-3p had a negative regulatory effect on KHDRBS1.

KHDRBS1 is a targeted binding molecule of miR-200b-3p. A Targetscan predicts the binding sites of miR-200b-3p and KHDRBS1. B Dual-luciferase reporter assay verified the targeted binding of miR-200b-3p and KHDRBS1. (* vs. MC; ***P < 0.001). C qRT-PCR analysis of KHDRBS1 expression in each group rats. (n = 10, # vs. Control; ^ vs. Model+agomiR-NC; § vs. Model+antagomir-NC. §§§P < 0.001; ***P < 0.001; ###P < 0.001). D Western blot analysis of KHDRBS1 expression in each group rats. E The signal intensities of KHDRBS1 were quantified. (n = 10, # vs. Control; ^ vs. Model+agomiR-NC; § vs. Model+antagomiR-NC. §§§P < 0.001; ***P < 0.001; ###P < 0.001). Group: Control group; Model group; Model+agomiR-NC group; Model+agomiR group; Model+antagomiR-NC group; Model+antagomiR group. qRT-PCR quantitative reverse transcription polymerase chain reaction, KHDRBS1 KH RNA binding domain containing, signal transduction associated 1

MiR-200b-3p alleviated TNF-α-induced apoptosis and TNF-α-inhibited proliferation through KHDRBS1. A The expression of miR-200b-3p in each group was detected by qRT-PCR. (n = 3, * vs. Blank; # vs. TNF-α+MC; ‡ vs. TNF-α+KHDRBS1. ***P < 0.001; ###P < 0.001; ‡‡‡P < 0.001). B qRT-PCR analysis of KHDRBS1 expression in each group (n = 3, * vs. Blank; # vs. TNF-α+MC; ^ vs. TNF-α+M; § vs. TNF-α+NC; ‡ vs. TNF-α+KHDRBS1. ***P < 0.001; ###P < 0.001; ‡‡‡P < 0.001; ^^^P < 0.001; §§§P < 0.001). C Western blot analysis of KHDRBS1 expression in each group. D The signal intensities of KHDRBS1 were quantified. (n = 3, * vs. Blank; # vs. TNF-α+MC; ^ vs. TNF-α+M; § vs. TNF-α+NC; ‡ vs. TNF-α+KHDRBS1. ***P < 0.001; ###P < 0.001; ‡‡‡P < 0.001; ^^^P < 0.001; §§§P < 0.001). E Cells viability was measured by MTT in each group (n = 3, * vs. Blank; # vs. TNF-α+MC; ^ vs. TNF-α+M; § vs. TNF-α+NC; ‡ vs. TNF-α+KHDRBS1. ***P < 0.001; ###P < 0.001; ‡‡‡P < 0.001; ^^^P < 0.001; §§§P < 0.001). F Apoptotic progression was determined by Flow cytometric analysis. G The results were expressed as percentages of positive mean values ± SD (n = 3, * vs. Blank; # vs. TNF-α+MC; ^ vs. TNF-α+M; § vs. TNF-α+NC; ‡ vs. TNF-α+KHDRBS1. ***P < 0.001; ##P < 0.01; ‡‡‡P < 0.001; ^^P < 0.01; §§§P < 0.001). Group: Blank group; TNF-α group; TNF-α+MC group; TNF-α+M group; TNF-α+NC group; TNF-α+KHDRBS1 group; TNF-α+M+KHDRBS1 group. qRT-PCR quantitative reverse transcription polymerase chain reaction, KHDRBS1 KH RNA binding domain containing, signal transduction associated 1, MTT 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, TNF-α tumor necrosis factor-α
MiR-200b-3p alleviated TNF-α-induced apoptosis and TNF-α-inhibited proliferation through KHDRBS1
MTT results showed that TNF-α decreased cell proliferation, as compared with the control group. The proliferation ability of TNF-α+M group was significantly increased, as compared with the TNF-α group. However, KHDRBS1 over-expression reversed the effect of miR-200b-3p over-expression on the proliferation ability. Over-expressed plasmids of miR-200b-3p and KHDRBS1 reversed the effect of KHDRBS1 on proliferation (Fig. 4E).
Similarly, TNF-α induced apoptosis, as compared with the control group. The apoptosis ability of TNF-α+M group was significantly lower than TNF-α group. KHDRBS1 over-expression could reverse the effect of miR-200b-3p on apoptosis. Addition both of miR-200b-3p and KHDRBS1 over-expressed plasmids reversed the effect of KHDRBS1 on apoptosis (Fig. 4F). To further explore the effects of miR-200b-3p and KHDRBS1 on apoptosis, the expressions of Bcl-2, Bax and C-Caspase-3 were also detected, and we found that Bcl-2 expression was down-regulated after TNF-α stimulation. Over-expression of miR-200b-3p could up-regulate the expression of Bcl-2. However, over-expression of KHDRBS1 further inhibited the expression of Bcl-2. Moreover, addition both of miR-200b-3p and KHDRBS1 over-expressed plasmids reversed the effect of KHDRBS1 on the expression of Bcl-2. The expression of Bax and C-Caspase-3 were different from that of Bcl-2 (Fig. 5A–F).

The effects of miR-200b-3p and KHDRBS1 on apoptosis-related proteins in vitro. A Western blot analysis of C-Caspase-3, Bcl-2 and Bax expression in each group. B–D The signal intensities of C-Caspase, Bcl-2 and Bax were quantified (n = 3, * vs. Blank; # vs. TNF-α+MC; ^ vs. TNF-α+M; § vs. TNF-α+NC; ‡ vs. TNF-α+KHDRBS1. **P < 0.01; ***P < 0.001; ###P < 0.001; ‡‡P < 0.01; ‡‡‡P < 0.001; ^^^P < 0.001; §§P < 0.01). E and F qRT-PCR analysis of Bcl-2 and Bax expression in each group rats (n = 3, * vs. Blank; # vs. TNF-α+MC; ^ vs. TNF-α+M; § vs. TNF-α+NC; ‡ vs. TNF-α+KHDRBS1. ***P < 0.01; ##P < 0.01; ###P < 0.001; ‡‡P < 0.01; ‡‡‡P < 0.001; ^^P < 0.01). Group: Blank group; TNF-α group; TNF-α+MC group; TNF-α+M group; TNF-α+NC group; TNF-α+KHDRBS1 group; TNF-α+M+KHDRBS1 group. qRT-PCR quantitative reverse transcription polymerase chain reaction, KHDRBS1 KH RNA binding domain containing, signal transduction associated 1, TNF-α tumor necrosis factor-α, Bcl-2 B cell lymphoma/lewkmia-2, Bax Bcl-2 associated X protein
These results suggested that miR-200b-3p alleviated TNF-α-inhibited proliferation and TNF-α-induced apoptosis thorugh negatively regulating KHDRBS1.
MiR-200b-3p alleviated TNF-α-induced inflammatory response through KHDRBS1
TGF-β, IL-10, IL-6, and IL-1β are important markers of inflammation. We performed Western blot and qRT-PCR to detect their protein and mRNA expressions, respectively. TGF-β, IL-6, and IL-1β proteins expressions were significantly increased after TNF-α pre-treatment (P < 0.01, P < 0.001). MiR-200b-3p over-expression reversed the effect of TNF-α (P < 0.01, P < 0.001), and over-expression of KHDRBS1 further promoted the effect of TNF-α (P < 0.01, P < 0.001). However, both over-expression of miR-200b-3p and KHDRBS1 only reversed the effect of KHDRBS1 over-expression (P < 0.01, P < 0.001) (Fig. 5A, B, D, E). IL-10 protein expression was the opposite (Fig. 5C). The mRNA expressions of TGF-β, IL-10, IL-6, and IL-1β were consistent with protein expressions (Fig. 5F–I). These results indicated that miR-200b-3p alleviated TNF-α-induced inflammatory response by negatively regulating KHDRBS1 (Fig. 6).

The effects of miR-200b-3p and KHDRBS1 on inflammatory factors in vitro. A Western blot analysis of TGF-β, IL-10, IL-6 and IL-1β expressions in each group. B–E The signal intensities of TGF-β, IL-10, IL-6 and IL-1β were quantified. (n = 3, * vs. Blank; # vs. TNF-α+MC; ^ vs. TNF-α+M; § vs. TNF-α+NC; ‡ vs. TNF-α+KHDRBS1. **P < 0.01; ***P < 0.001; ##P < 0.01; ###P < 0.001; ‡‡P < 0.01; ‡‡‡P < 0.001; ^^^P < 0.05; ^^^P < 0.01; ^^^P < 0.001; §§P < 0.01; §§§P < 0.001). F-I qRT-PCR analysis of TGF-β, IL-10, IL-6 and IL-1β expression in each group rats (n = 3, * vs. Blank; # vs. TNF-α+MC; ^ vs. TNF-α+M; § vs. TNF-α+NC; ‡ vs. TNF-α+KHDRBS1. **P < 0.01; ***P < 0.01; ###P < 0.001; ‡‡‡P < 0.001; ^^P < 0.01; P < 0.001). Group: Blank group; TNF-α group; TNF-α+MC group; TNF-α+M group; TNF-α+NC group; TNF-α+KHDRBS1 group; TNF-α+M+KHDRBS1 group. qRT-PCR quantitative reverse transcription polymerase chain reaction, KHDRBS1 KH RNA binding domain containing, signal transduction associated 1, TNF-α tumor necrosis factor-α, Bcl-2 B cell lymphoma/lewkmia-2, Bax Bcl-2 associated X protein, TGF-β transforming growth factor beta, IL-10 interleukin 10, IL-6 interleukin 6, IL-1β interleukin 1β
Discussion
UC is a chronic intestinal inflammatory disease with unclear etiology, and is one of the most difficult diseases to be treated in the world (Warren and Sommers 1949). IECs, which are the main functional cells in the intestinal tract and the structural basis of the intestinal epithelial barrier, regulate intestinal epithelial permeability through the transcellular pathway (Benjamin et al. 2000). High intestinal permeability is a main cause of UC (Benjamin et al. 2000). Therefore, studying IEC could help understand UC. Studies have shown that miR-200b could protect the intestinal epithelial barrier through transepithelial membrane pathway (Chen et al. 2013). Our study found that the expression of miR-200b-3p in colon tissues of UC rats was significantly decreased, which was also consistent with previous studies. Further experiments showed that the down-regulation of miR-200b-3p could increase the disease activity index of rats and promote the colon lesions of rats. In addition, we also detected the expression levels of inflammatory markers in UC rats, and the results showed that miR-200b-3p could inhibit the occurrence of inflammatory response.
It has been found that inflammatory responses caused apoptosis (Peng et al. 2007). During the inflammatory response, neutrophils and other holdup cells are recognized and cleared by macrophages through apoptotic mechanisms, while the membrane remains intact to avoid a secondary response (Platt et al. 2000). Regulating death is an important mechanism through which the body regulates the development of inflammatory response and prevents the injury and scar formation of tissues after inflammation. We hypothesized that miR-200b-3p could reduce apoptosis. Bax and Bcl-2 belong to the Bcl-2 gene family (Oh and Lim 2007). Bcl-2 is an apoptosis suppressor gene. Bax antagonizes Bcl-2, and promotes cell apoptosis (Roucou et al. 2002). In addition, C-Caspase-3 is the ultimate executor of apoptosis (Qiu et al. 2000). By detecting the expression levels of these genes, we found that miR-200b-3p could inhibit apoptosis.
The target binding molecules of miR-200b-3p were predicted by Targetscan database, and the candidate molecule KHDRBS1 was determined and further confirmed by double-luciferase reporter gene assay. KHDRBS1 is involved in cell inflammation and apoptosis in several studies (Sun et al. 2018). By detecting the expression of KHDRBS1, we found that miR-200b-3p had a negative regulatory effect on KHDRBS1. Tumor necrosis factor–α (TNF-α) is often used to create an inflammatory environment (Graham et al. 2006; Shen et al. 2006). In this study, IEC is also treated with TNF-α to create inflammatory environment. Expression of miR-200b-3p in IEC (treated with TNF-α) was down-regulated. MiR-200b-3p could alleviate TNF-α-induced apoptosis and inflammatory response, and TNF-α-inhibited proliferation by negatively regulating KHDRBS1.
In summary, our study firstly proved miR-200b-3p attenuated UC lesions and reduced disease activity index. Further investigation revealed that miR-200b-3p alleviated TNF-α-induced apoptosis and inflammatory response through negative regulation of KHDRBS1. Based on the above studies, we basically proved that the regulatory relationship between miR-200b-3p and KHDRBS1 plays an important role in the development of UC, which may provide a new idea for the treatment of UC.
References
Benjamin MA, McKay DM, Yang PC, Cameron H, Perdue MH (2000) Glucagon-like peptide-2 enhances intestinal epithelial barrier function of both transcellular and paracellular pathways in the mouse. Gut 47:112–119. https://doi.org/10.1136/gut.47.1.112
Chen Y et al (2013) miR-200b inhibits TGF-β1-induced epithelial-mesenchymal transition and promotes growth of intestinal epithelial cells. Cell Death Dis 4:e541. https://doi.org/10.1038/cddis.2013.22
Conrad K, Roggenbuck D, Laass MW (2014) Diagnosis and classification of ulcerative colitis. Autoimmun Rev 13:463–466. https://doi.org/10.1016/j.autrev.2014.01.028
Fabbri M et al (2012) MicroRNAs bind to toll-like receptors to induce prometastatic inflammatory response. Proc Natl Acad Sci USA 109:E2110-2116. https://doi.org/10.1073/pnas.1209414109
Graham WV et al (2006) Tumor necrosis factor-induced long myosin light chain kinase transcription is regulated by differentiation-dependent signaling events. Characterization of the human long myosin light chain kinase promoter. J Biol Chem 281:26205–26215. https://doi.org/10.1074/jbc.M602164200
Khor B, Gardet A, Xavier RJ (2011) Genetics and pathogenesis of inflammatory bowel disease. Nature 474:307–317. https://doi.org/10.1038/nature10209
Lukong KE, Richard S (2003) Sam68, the KH domain-containing superSTAR. Biochim Biophys Acta 1653:73–86. https://doi.org/10.1016/j.bbcan.2003.09.001
McKenna LB, Schug J, Vourekas A, McKenna JB, Bramswig NC, Friedman JR, Kaestner KH (2010) MicroRNAs control intestinal epithelial differentiation, architecture, and barrier function. Gastroenterology 139:1654–1664. https://doi.org/10.1053/j.gastro.2010.07.040
Motoyama S et al (2009) CRP genetic polymorphism is associated with lymph node metastasis in thoracic esophageal squamous cell cancer. Ann Surg Oncol 16:2479–2485. https://doi.org/10.1245/s10434-009-0525-2
Nguyen HT et al (2010) MicroRNA-7 modulates CD98 expression during intestinal epithelial cell differentiation. J Biol Chem 285:1479–1489. https://doi.org/10.1074/jbc.M109.057141
Noman MZ et al (2017) The immune checkpoint ligand PD-L1 is upregulated in EMT-activated human breast cancer cells by a mechanism involving ZEB-1 and miR-200. Oncoimmunology 6:e1263412. https://doi.org/10.1080/2162402x.2016.1263412
Oh PS, Lim KT (2007) HeLa cells treated with phytoglycoprotein (150 kDa) were killed by activation of caspase 3 via inhibitory activities of NF-kappaB and AP-1. J Biomed Sci 14:223–232. https://doi.org/10.1007/s11373-006-9140-4
Peck BC, Mah AT, Pitman WA, Ding S, Lund PK, Sethupathy P (2017) Functional transcriptomics in diverse intestinal epithelial cell types reveals robust microRNA Sensitivity in intestinal stem cells to microbial status. J Biol Chem 292:2586–2600. https://doi.org/10.1074/jbc.M116.770099
Peng Y, Martin DA, Kenkel J, Zhang K, Ogden CA, Elkon KB (2007) Innate and adaptive immune response to apoptotic cells. J Autoimmun 29:303–309. https://doi.org/10.1016/j.jaut.2007.07.017
Platt N, Suzuki H, Kodama T, Gordon S (2000) Apoptotic thymocyte clearance in scavenger receptor class A-deficient mice is apparently normal. J Immunol 164:4861–4867. https://doi.org/10.4049/jimmunol.164.9.4861
Purrmann J, Strohmeyer G (1989) Pathogenesis and management of ulcerative colitis. Hepatogastroenterology 36:209–212
Qian J, Zhao W, Miao X, Li L, Zhang D (2016) Sam68 modulates apoptosis of intestinal epithelial cells via mediating NF-κB activation in ulcerative colitis. Mol Immunol 75:48–59. https://doi.org/10.1016/j.molimm.2016.05.011
Qiu JH, Asai A, Chi S, Saito N, Hamada H, Kirino T (2000) Proteasome inhibitors induce cytochrome c-caspase-3-like protease-mediated apoptosis in cultured cortical neurons. J Neurosci 20:259–265. https://doi.org/10.1523/jneurosci.20-01-00259.2000
Roucou X, Rostovtseva T, Montessuit S, Martinou JC, Antonsson B (2002) Bid induces cytochrome c-impermeable bax channels in liposomes. Biochem J 363:547–552. https://doi.org/10.1042/0264-6021:3630547
Saber S, Khalil RM, Abdo WS, Nassif D, El-Ahwany E (2019) Olmesartan ameliorates chemically-induced ulcerative colitis in rats via modulating NFκB and Nrf-2/HO-1 signaling crosstalk. Toxicol Appl Pharmacol 364:120–132. https://doi.org/10.1016/j.taap.2018.12.020
Shen L, Black ED, Witkowski ED, Lencer WI, Guerriero V, Schneeberger EE, Turner JR (2006) Myosin light chain phosphorylation regulates barrier function by remodeling tight junction structure. J Cell Sci 119:2095–2106. https://doi.org/10.1242/jcs.02915
Stone CD (2012) The economic burden of inflammatory bowel disease: clear problem, unclear solution. Dig Dis Sci 57:3042–3044. https://doi.org/10.1007/s10620-012-2417-8
Strober W, Fuss IJ (2011) Proinflammatory cytokines in the pathogenesis of inflammatory bowel diseases. Gastroenterology 140:1756–1767. https://doi.org/10.1053/j.gastro.2011.02.016
Sun W et al (2018) Sam68 promotes invasion, migration, and proliferation of fibroblast-like synoviocytes by enhancing the NF-κB/P65 pathway in rheumatoid arthritis. Inflammation 41:1661–1670. https://doi.org/10.1007/s10753-018-0809-4
Vlantis K et al (2016) NEMO Prevents RIP kinase 1-mediated epithelial cell death and chronic intestinal inflammation by NF-κB-dependent and -independent functions. Immunity 44:553–567. https://doi.org/10.1016/j.immuni.2016.02.020
Warren S, Sommers SC (1949) Pathogenesis of ulcerative colitis. Am J Pathol 25:657–679
Xavier RJ, Podolsky DK (2007) Unravelling the pathogenesis of inflammatory bowel disease. Nature 448:427–434. https://doi.org/10.1038/nature06005
Xie Z, Allen E, Fahlgren N, Calamar A, Givan SA, Carrington JC (2005) Expression of arabidopsis MIRNA genes. Plant Physiol 138:2145–2154. https://doi.org/10.1104/pp.105.062943
Yang E, Zha J, Jockel J, Boise LH, Thompson CB, Korsmeyer SJ (1995) Bad, a heterodimeric partner for Bcl-XL and Bcl-2, displaces bax and promotes cell death. Cell 80:285–291. https://doi.org/10.1016/0092-8674(95)90411-5
Zhang SQ, Ni WK, Xiao MB, Jiang F, Lu CH, Wang RH, Ni RZ (2019) Actin related protein 3 (ARP3) promotes apoptosis of intestinal epithelial cells in ulcerative colitis. Pathol Res Pract 215:235–242. https://doi.org/10.1016/j.prp.2018.10.011
Funding
This work was supported by the Traditional Chinese Medicine Technology Development Plan of Jiangsu Province [YB2020075].
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Zheng, C., Lu, T. & Fan, Z. miR-200b-3p alleviates TNF-α-induced apoptosis and inflammation of intestinal epithelial cells and ulcerative colitis progression in rats via negatively regulating KHDRBS1. Cytotechnology 73, 727–743 (2021). https://doi.org/10.1007/s10616-021-00490-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10616-021-00490-3