
Abstract
Changing magnitude, frequency, and timing of precipitation can influence aquatic-system hydrological, geochemical, and biological processes, in some cases resulting in system-wide shifts to an alternate state. Since the early 1990s, the southern Prairie Pothole Region has been subjected to an extended period of increased wetness resulting in marked changes to aquatic systems defining this region. We explored numerous lines of evidence to identify: (1) how the recent wet period compared to historical variability, (2) hydrological, geochemical, and biological responses, and (3) how these responses might represent a state shift in the region’s wetland ecosystems. We analyzed long-term climate records and compared how different hydrological variables responded in this wet period compared to decades before the observed shift. Additionally, we used multi-decadal records of waterfowl population and subsurface tile drain records to explore wildlife and human responses to a shifting climate. Since 1993, a novel precipitation regime corresponded with increased pond numbers, ponded-water depths, lake levels, stream flows, groundwater heights, soil-moisture, waterfowl populations, and installation of subsurface tile drains in agricultural fields. These observed changes reflect an alteration in water storage and movement across the landscape that in turn has altered solute sources and concentrations of prairie-pothole wetlands and has increased pond permanence. Combined, these changes represent significant evidence for a state shift in the ecohydrological functioning of the region’s wetland ecosystems, a shift that may require a significant refinement of the previously developed “wetland continuum” concept.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The North American Prairie Pothole Region (PPR) encompasses a wide variety of hydrologically distinct ecosystems that support critical migratory-bird habitat, dynamic aquatic communities, extensive grasslands, and productive agricultural systems. The PPR covers approximately 770,000 km2 of the USA and Canada (Smith et al. 1964) (Fig. 1). The advance and retreat of the Laurentide ice sheet (Dyke and Prest 1987) created a landscape of low-permeability soils and millions of closed-catchment lake and wetland basins (Goldhaber et al. 2011). These depressional basins vary topographically along the landscape in relation to groundwater, which causes differences among aquatic systems in terms of permanence and salinity of ponded water (Winter 2003). Wetlands located above the local water table typically have temporarily ponded water and are able to lose solutes to groundwater, which keeps their waters fresh. Wetlands located lower on the landscape tend to have ponds that are more permanent and accumulate solutes from discharging groundwater or via surface connections to other saline wetlands. Substantial hydrogeochemical variability among wetlands has created distinct habitats that support a variety of plant, invertebrate, and vertebrate communities (Mushet et al. 2015).

Geographic location and extent of climatological, hydrological, and biological datasets used to explore an ecohydrological state shift in the southern Prairie Pothole Region (PPR). Bottom left inset shows the extent of the full PPR in light blue. Red stars represent stream gages. Purple diamond represents the Cottonwood Lake Study Area. Orange linear features represent Hydro-Climatic Data Network streams. Green polygon represents North Dakota Climate Division 5. Purple polygon represents Sheyenne River watershed. Teal polygon represents the US Fish and Wildlife Service Stratum #46. Pink rectangle highlights Devils Lake (gray)
The PPR climate is continental and highly variable both spatially and temporally (Rosenberry 2003; Mushet 2016) with year-to-year weather conditions strongly influencing hydrological status of the region’s wetlands, lakes, streams, and groundwater (Liu and Schwartz 2012; Hayashi et al. 2016; LaBaugh et al. 2016). Annual precipitation has displayed patterns of both high inter-annual variability (Hayashi et al. 2016) and decadal oscillations between dry and wet modes (Winter and Rosenberry 1998). The rapid response of surface-water levels to increasingly dynamic weather patterns can be exacerbated in this region because of the extensive network of hydrologically connected wetland basins that store water on the landscape (Stichling and Blackwell 1957; Leibowitz et al. 2016). Snowmelt is an essential hydrological input sustaining wetlands, lakes, and streams in the PPR (Shook and Pomeroy 2012). Historically, snowmelt could account for as much as 90% of the total surface runoff when soils are frozen at the time that the snow melts. (Pomeroy et al. 1998).
The southern (i.e., USA) portion of the PPR has been in an extended “wet period” since 1993 (Ballard et al. 2014), details of which will be expanded herein. More recently, these wet conditions have extended north into Canadian portions of the PPR (Hayashi et al. 2016), including increases in runoff-producing multi-day storms (Shook and Pomeroy 2012). These increasingly wet conditions may have implications in runoff generated by both snowmelt and high-intensity rainstorms (Dumanski et al. 2015). Additionally, the wet conditions influencing the PPR coincide with air temperatures that are increasing by about 0.14 °C per decade (Todhunter and Fietzek-DeVries 2016).
Prairie-pothole-wetland trophic structures are strongly influenced by both pond permanence and salinity (McLean et al. 2016a). The most immediate effects of a wetter climate on wetland ecosystems in the PPR have been an increase in pond permanence and an overall decrease in salinity (Mushet et al. 2015). However, LaBaugh et al. (2016) recently showed how sustained high water in the region’s wetlands can lead to increasing salt loads in wetland ponds. These shifts in pond permanence and salinity are affecting plant (Winter 2003), macroinvertebrate (McLean et al. 2016a), and waterfowl communities (Euliss et al. 2004). Transitioning to more permanently inundated and less-saline wetlands can increase suitable habitat for the persistence of fish populations and potentially facilitate movements among aquatic habitats and human translocations (McLean et al. 2016b). Increased fish populations introduce potential forage competition for waterfowl (Cox et al. 1998), and shifts in wetland-plant communities driven by changes in solutes and/or water levels influence available habitat for breeding (Steen et al. 2016). Future increases in precipitation and temperature are likely to continue these novel biotic interactions (Ballard et al. 2014; Johnson and Poiani 2016).
Agroecosystems, which are widespread and economically important in the PPR (Gascoigne et al. 2013), also are affected by a wetter climate. Increased surface-water on the landscape can necessitate installation of subsurface tile drains (Blann et al. 2009), which have been used extensively to remove surface-waters in other Midwestern US states, including Iowa, Illinois, Indiana, and Ohio, since the late 1800s. The 1978 Census of Agriculture found that over 30 million acres (120,000 km2) had been altered via subsurface tile drains (Pavelis 1987).
We focused our research on the southern PPR, the portion that has experienced this “wet period” for the longest time and to the greatest extent. We pursued three research objectives to determine the extent that increased precipitation has changed wetland ecosystems in the southern PPR: (1) determine if increases in wetness in the southern PPR are beyond recent historical variability; (2) quantify hydrological, chemical, and biological (including human) responses; and (3) explore the possibility that identified hydrological, chemical, and biological responses represent an ecosystem-level state shift in the wetland systems of this ecologically and economically important region. To address these objectives, we first analyzed long-term climate records. We examined a number of long-term surface-water, groundwater, and soil-moisture records from North Dakota in the southern PPR to compare hydrological functioning during the recent wet period to hydrologic functioning during decades before 1993. We also used long-term records of regional waterfowl production and subsurface tile drain permits to assess the ecosystem and human responses to different climate regimes. We then evaluated the possibility that observed changes occurring in the southern PPR have resulted in a shift in the ecohydrological state of the region’s wetland ecosystems not yet accounted for by the widely used “wetland continuum” conceptual framework (Euliss et al. 2004).
2 Methods and datasets
2.1 Study area
The southern PPR is located south of Manitoba and Saskatchewan, Canada, and north and east of the Missouri River in the USA (Fig. 1). Soils in the southern PPR are variable but can be up to 20 m deep, and high clay content can cause very low permeability in some parts of the region (Goldhaber et al. 2014). November through March is cold and dry; average temperature is below 0 °C, and average precipitation prior to the recent wet period was less than 70 mm. Snowfall accumulated during this period is crucial for supplying freshwater to the region’s aquatic systems (Renton et al. 2015). The warmest and wettest part of the year is usually June through August with average temperatures around 20 °C and rainfall (before 1993) totaling nearly 220 mm. Summer rainfall is typically generated by high-intensity thunderstorms and accounts for approximately half of annual precipitation (Rosenberry 2003; Vecchia 2008).
2.2 Climate shift: meteorological datasets
We chose meteorological datasets that represented both precipitation and evapotranspiration, and extended over more than a century. We used meteorological datasets from North Dakota Climate Division 5 (Fig. 1). We used the precipitation dataset (1895–2015) from the Global Historical Climatology Network-Monthly version 2 (GHCN-M) (Menne et al. 2009). We also analyzed changes in the monthly Palmer Hydrological Drought Index (PHDI) (Palmer 1965; Karl et al. 1987). PHDI is one of the most widely used drought indices with values above zero indicating wet conditions and values below zero indicating dry conditions (Vicente-Serrano et al. 2012). Temperature and precipitation are used in PHDI for calculating water balance components such as runoff, evapotranspiration, and soil recharge (Karl 1983). We chose PHDI over PDSI because PHDI is considered more appropriate for tracking long-term, surface-water dynamics (Dai 2011; Vicente-Serrano et al. 2012). Detailed methods for calculating PHDI can be found in Karl (1983). We obtained monthly PHDI data for 1895–2015 from the NOAA National Climate Data Center (http://www1.ncdc.noaa.gov/pub/data/cirs/). We also calculated potential evapotranspiration using the temperature-based Hamon equation (Hamon 1961) and compared annual and monthly means for the periods 1895–1992 and 1993–2015 (Fig. S1).
2.3 Hydrological and biological response datasets
We used various multi-decadal hydrological datasets from across the southern PPR. We analyzed pond-elevation (Mushet et al. 2015) and groundwater-depth (Winter 2003) data from wetlands of the Cottonwood Lake Study Area (CLSA), which is located on the eastern edge of the Missouri Coteau in Stutsman County, North Dakota (Winter 2003). The CLSA data we used came from eight “temporarily” ponded wetlands (T2–T9) that historically dried seasonally, and eight “semi-permanently” ponded wetlands (P1–P4, P6–P8, P11) that retain water year round except during periods of severe drought (Mushet et al. 2016). We also used annual average water-table heights (1978–2015) from 16 CLSA groundwater wells (wells 2–16 and 54). We analyzed USGS water-elevation data for Devils Lake (1932–2016; https://nd.water.usgs.gov/devilslake/data/) (Todhunter 2016). To measure changes in the number of wetland ponds available to breeding waterfowl, we used the US Fish and Wildlife (USFWS) Spring Pond Count data (1955–2015) from stratum #46 (Fig. 1) located in southeastern North Dakota (USFWS 2016).
We analyzed trends in discharge for the major streams and rivers included in the USGS Hydro-Climatic Data Network 2009 (HCDN-2009) from 1938 to 2015. Streams used were the five HCDN-2009 streams in the southern PPR, i.e., Sheyenne River (USGS 05057000), Baldhill Creek (USGS 05057200), Wintering River (USGS 05120500), Willow Creek (USGS 05123400), and James River (USGS 06468170) (Fig. 1). All stream gage data was obtained from https://waterdata.usgs.gov. We also conducted a separate, more detailed analysis that focused on flow (i.e., river discharge) responses of the Sheyenne River to precipitation before and since 1993. This precipitation to discharge relationship is controlled by soil-moisture status in the watershed (Fig. 1) (Todhunter 2016). We used modeled average annual soil-moisture data (1932–2015) from the Climate Prediction Center (CPC) (http://www.cpc.ncep.noaa.gov/). Modeled soil-moisture at the end of each month came from a one-layer water balance model that was driven by air temperature, monthly precipitation, and calibrated runoff data. The model generates an average for a soil column of 1.6 m with a common porosity of 0.47 that gives a maximum holding capacity of 760 mm (Huang et al. 1996; van den Dool et al. 2003).
We chose biological response variables that represented both waterfowl production and changes in agricultural practices. We used data from the Waterfowl Breeding Population and Habitat Survey (USFWS 2016) to measure changes in breeding-waterfowl populations in southeastern North Dakota stratum #46 from 1955 to 2015. Stratum #46 encompasses 78,000 km2 and is representative of wetland distribution found throughout the southern PPR (Fig. 1). The USFWS conducted these surveys each May. To analyze the human response to excess water in agricultural fields, we used counts of subsurface tile drain permits (1993–2016) issued by the North Dakota State Water Commission (Finocchiaro 2016). North Dakota drain permitting has been recorded since 1975 (Sando 2015).
2.4 Wetland ecosystem state shift
In addition to analyzing pond depths across wetlands at CLSA, we also conducted a detailed analysis of changes in salinity in CLSA wetland P1. We used total dissolved solids (TDS) (kg) in the P1 pond updated from LaBaugh et al. (2016) as a proxy for P1 salinity. TDS was estimated from an empirical relationship between specific conductance and TDS (R 2 = 0.97). Calibration techniques are detailed in LaBaugh et al. (2016). We also qualitatively evaluated how observed changes in salinity and pond depth can move a wetland ecosystem in novel directions along different axes of the wetland continuum conceptual model (Euliss et al. 2004).
2.5 Statistical analysis
Means and coefficients of variation were determined for two periods, years before 1993 and 1993–2015. We evaluated changes in means of each of our climatological, hydrological, and biological variables using Student’s t tests and reported all t statistics generated from these analyses. Statistical significance was indicated as p values at α = 0.05. We also used segmented regression analysis and Student’s t test to identify best-fit slopes and compare cumulative discharge to cumulative precipitation in the Sheyenne River before and after 1993. Segmented regression analysis was used to estimate any breakpoints in the Sheyenne River analysis using the “segmented” package in R (Muggeo 2008). We conducted all analyses and generated all figures using R version 3.0.2 (R Core Team 2016).
3 Results
3.1 Climate shift
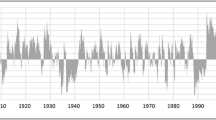
In addition to significant warming over the last century (Todhunter and Fietzek-DeVries 2016), we found there was an increase in overall wetness (Fig. 2). Annual precipitation during 1993–2016 increased by 55 mm, or 12% when compared to 1895–1992 (t = 1.99, p = 0.02) (Fig. 2b). We found that precipitation increases occurred mostly in the spring, summer, and early fall. Monthly precipitation increased by 18 mm in May (t = 2.22, p = 0.03), 16 mm in July (t = 2.48, p = 0.02), and 16 mm in October (t = 2.73, p = 0.007) (Fig. S2). While precipitation significantly increased and despite gradual temperature increases (Todhunter and Fietzek-DeVries 2016), annual potential evapotranspiration has not significantly increased since 1993 (Fig. S1). The average monthly PHDI also significantly increased (t = 11.3, p < 0.0001) from the 1895–1992 period (\( \overline{x} \) = 0.60) to the 1993–2016 period (\( \overline{x} \) = 2.66) (Fig. 2a). A value of 0.60 is considered “near normal,” whereas a value of 2.66 would be considered “unusually moist” (Palmer 1965). The years 1993–2016 included the highest monthly PHDI values recorded since 1895 and a significant increase in the proportion of months with PHDI values greater than zero (Table S1).

a Average monthly Palmer Hydrological Drought Index from 1895 to 2015 for North Dakota Climate Division 5 (NOAA 2016). Positive values indicate magnitude of wet conditions whereas negative values indicate severity of drought. b Total annual precipitation from 1895 to 2015 for North Dakota Climate Division 5 (NOAA 2016). Horizontal dashed line represents long-term mean annual precipitation. Vertical dashed line in both panels highlights 1993. Gray rectangles behind plots identify the pre-1993 and post-1993 range between maximum and minimum values
3.2 Hydrological and biological response datasets
Each of the ecohydrological variables we examined exhibited quantitatively significant changes in functioning since 1993 and correlated with the change to a wetter climate (Fig. 3). Increased precipitation corresponded with significant increases in monthly average soil-moisture (mm) for all months across North Dakota Climate Region 5 (Fig. S2). Mean annual soil-moisture storage (mm) in the southern PPR significantly increased (t = 1.98, p < 0.0001) by 16% (40 mm) from 1932–1992 to 1993–2015 (Fig. 3a). Inter-annual soil-moisture coefficient of variation has also decreased by 20% since 1993. This decrease in soil-moisture variability corresponded with a 57-mm increase in the minimum annual soil-moisture from 171 mm before 1993 to 228 mm since 1993. The largest monthly soil-moisture increase occurred in October with a significant increase of 49 mm (t = 4.61, p < 0.0001) (Fig. S2). Groundwater heights also showed a significant increase of ~ 2 m on average (t = 6.9, p < 0.0001) since 1993 (Fig. 3b). Groundwater heights increased in each of the 16 wells, with the greatest increases of ~ 4 m (1978–2015) occurring in two of the 16 wells (Fig. 3b).

a Modeled mean annual average soil-moisture 1932–2015 for North Dakota Climate Division 5. b Mean annual elevations of 16 groundwater wells located across a topographic gradient in the Cottonwood Lake Study Area, 1978–2015. c Mean annual depth of ponded water for eight semi-permanently ponded prairie-pothole wetlands (gray squares) and the eight seasonally ponded prairie-pothole wetlands (black squares) in the Cottonwood Lake Study Area 1978–2015. d Mean annual elevation of Devils Lake water surface from 1932 to 2015. e Mean annual daily discharge for eastern North Dakota streams included in the Hydro-Climatic Data Network (HCDN) 1938–2015. f The US Fish and Wildlife Service annual pond counts for stratum 46 in the southern Prairie Pothole Region. g The US Fish and Wildlife Service annual waterfowl counts for stratum 46 in the southern Prairie Pothole Region. h Number of annual subsurface tile drain permits issued in North Dakota 1975–2015. The vertical dashed line in all graphs demarks the beginning of a period of increased precipitation in 1993. Error bars represent standard error
Surface-water in the southern PPR has also responded much differently in the last 23 years compared to years prior to 1993. Average wetland pond water depths significantly increased (t = 5.9, p < 0.0001) by ~ 1 m at the CLSA (Fig. 3c). The semi-permanently ponded (P) wetlands have become much deeper, lake-like, systems. Before 1993, the average pond depth of P wetlands at CLSA was 0.45 m, and the deepest pond was just over 1 m. Since 1993, the average annual depth for P wetland ponds has increased by four times to 1.83 m, and the deepest of these ponds was near 4-m deep. The temporarily ponded (T) wetlands have become much more consistently ponded since 1993. Since 1993, all of the T wetlands contained ponded surface-water at some point during the year. Similar trends existed on a larger spatial scale in Devils Lake. The average depth of Devils Lake has increased significantly (t = 15.7, p < 0.0001) by ~ 10 m, with the majority of this increase occurring from 1993 to 1999; the lake level has remained above 440 MASL since then (Fig. 3d), despite pumping efforts that were initiated by the state of North Dakota in 2007 (Vecchia 2008). The mean number of wetlands with ponded water present during the spring Waterfowl Breeding Population and Habitat Survey has also significantly increased since 1993 (t = 7.4, p < 0.0001) (US Fish and Wildlife USFWS 2016). The mean number of ponds has more than doubled in the southern PPR from 132,748 in 1955–1992 to 297,545 in 1993–2015 (Fig. 3f).
The collective increase in surface-water in the southern PPR also correlated with biological, including human, responses since 1993. The mean annual breeding-waterfowl population has significantly increased (t = 9.8, p < 0.0001) by almost threefold since 1993, from 0.76 million birds (1955–1992) to 2.0 million birds (1993–2015) (Fig. 3g). This increase in annual breeding waterfowl population was strongly and linearly correlated with the increased annual number of ponded wetlands (R 2 = 0.72, p = 0.001). Subsurface tile drain installation also correlated with increased landscape surface moisture. No tile drain permits were issued in North Dakota prior to 1993. The installation of tile drains exponentially increased from 2002 to 2010 (Fig. 3h). The most tile drain permits issued in 1 year was 206 in 2013. This spike in the issuance of subsurface tile drain permits comes after the three highest recorded years of soil-moisture for the southern PPR (Fig. 3a) and a resultant need to remove surface and near-surface water to sustain agricultural productivity.
Mean daily discharge of our five study streams also significantly increased (t = 5.4, p < 0.0001) from 0.5 m3/s (1945–1992) to 3.8 m3/s (1993–2015) (Fig. 3e). Discharge in the Sheyenne River became 2.7 times more sensitive to precipitation from 1993 to 2015 as compared to 1945–1992. A double mass-curve analysis performed by Todhunter and Rundquist (2004), and extended here, revealed a linear relationship between precipitation input and runoff response (i.e., cumulative discharge) for the Sheyenne River, with a distinct change in slope from 0.06 to 0.16 in 1993 (Fig. 4). We verified this change in slope via segmented regression analysis that identified 1993 as the year with the lowest mean squared error and the only breakpoint.

Sheyenne River cumulative discharge versus cumulative annual precipitation 1945–2015. Black dashed line represents the beginning of the recent wet period in 1993. Gray line represents the best-fit model 1945–1992 and the black, solid line represents the best-fit model 1993–2015. The data from 2007 to 2015 include subtraction of discharge values from Devil’s Lake pumping. Figure expanded and adapted from Todhunter and Rundquist (2004)
3.3 Wetland ecosystem state shift
Along with increased water in prairie-pothole wetlands, there has also been an increase in salt loads in semi-permanent ponds (Fig. 5a). Mean TDS mass in CLSA wetland P1 before 1993 was 14,600 kg; after 1993, mean TDS was 131,800 kg. In addition, since 1993, mean TDS in wetland P1 has increased by 8900 kg per year (Fig. 5a). These results also indicate that wetlands may move in novel directions along the wetland continuum (Fig. 5b). Traditionally, wetlands that supported salt-tolerant vegetation communities existed during “drought” periods, as shown on the y-axis of Fig. 5b. Figure 5a shows a wetland that is rich in salts but operating during a “deluge” period.

a Hydrologic and chemical conditions in wetland P1 at the Cottonwood Lake Study Area (1980–2015). Water volume is represented by black circles and mass of total dissolved solids (TDS) is represented by gray circles (updated from LaBaugh et al. 2016). b The wetland continuum framework (Euliss et al. 2004) modified to show an example state shift (lettered arrows) in a wetland’s hydrologic relation to groundwater. A wetland at position “A” that is changing in response to oscillations between wet and dry conditions can shift along “B” to a new state “C” if altered hydrologic conditions favor the buildup of salt loads during an extended wet period. The wetland will likely remain at this new position, with its higher salt loads and more salt tolerant biotic communities, until hydrologic conditions revert “?” to a condition favoring the export of accumulated salt
4 Discussion
The observed shift in climate has continued to persist even as inter-annual and decadal patterns such as the El Niño–Southern Oscillation (ENSO) or the Pacific Decadal Oscillation (PDO) have changed from wet to dry modes (Huang et al. 2015). Ballard et al. (2014) found that both ENSO and PDO were not strong drivers of PPR climate and surface-water levels. Not only were there higher PHDI values during this wet period than at any other time in the past century, but there was also a conspicuous absence of extremely low (i.e., dry) PHDI values (Table S1).
Todhunter (2016) and others have postulated that the current wet period began in 1980 and is part of a normal shift between long-term dry and wet modes. Our evidence indicates this shift occurred in 1993, is novel, and is distinctly different from previously measured oscillations. Water levels reflective of current conditions have likely not occurred in the southern PPR during the past 500 years (Winter and Rosenberry 1998) and certainly not during the last 100 years. With an absence of the normal cyclical dry-down periods, there has been a prolonged recharging of soil-moisture, surface-water, and groundwater. The enhanced response of discharge in areas such as the Sheyenne River in the last 23 years (Fig. 4) illustrates how upland soils that are close to saturation can more efficiently transform both snowmelt and precipitation into runoff.
Another example of an ecohydrological response is from the Devils Lake Basin, where runoff into the lake annually has increased by ~ 400% over the last 30 years (Todhunter 2016). It is fair to assume that a similar mechanism is driving the large increases in pond-water depth (Fig. 3c) and pond number (Fig. 3f) across the southern PPR. Other studies of the wetland habitats across the PPR have found similar responses (Mushet et al. 2015; Cressey et al. 2016; Hayashi et al. 2016). It is likely that increases in soil-moisture (Fig. 3a) are also causing enhanced snowmelt runoff during spring runoff events (Shook and Pomeroy 2012). Even though precipitation during winter months has not increased (Fig. S2), more soil-moisture would cause less snowpack infiltration and more effective runoff. Additionally, increased soil-moisture during fall would contribute to frost seals in the soil that enhance spring runoff events. Precipitation increases during summer and fall have a compounded effect on runoff when intense rainfall from thunderstorms occurs when soils are saturated (Fig. 3b) (Rosenberry 2003).
Although the evidence we reviewed clearly suggests climate has been the dominant driver of hydrologic change in the southern PPR, others have postulated that agricultural land-use change and/or wetland drainage have been the primary drivers of increased water inputs to wetlands in the region (Mccauley et al. 2015). We offer two lines of reasoning to support the climate-shift hypothesis. First, all of the changes we observed started in 1993, concurrent with the beginning of the extreme wet period (Fig. 3). Recent analysis of land-use conversion indicates there was no significant change in either land use or wetland drainage initiated in 1993 that can explain the observed hydrologic changes (Drummond et al. 2012; Wright and Wimberly 2013). Drummond et al. (2012) found the largest significant land-conversion rates occurred from 1992 to 2000, however the driver of this change was a conversion of wetlands to open-water. Wright and Wimberly (2013) indicated that major shifts to increased corn and soy production in the region did not occur until 2006, well after the hydrologic shifts in 1993. Furthermore, increases in groundwater and wetland water levels at the CLSA (Fig. 3b, c) could not have resulted from land-use changes because the site where those data were collected has not been altered from its natural state (Mushet et al. 2015). Second, the number of wetlands containing ponded water has increased dramatically since 1993. Consolidation drainage proposed by Anteau (2012) as the primary driver leading to increased pond levels in wetlands of the southern PPR would result in a decrease in number and area of wetland ponds, the goal of this land management practice, as numerous small ponds are drained (i.e., consolidation) into a smaller number of larger, but deeper, ponds. Instead, starting in 1993, the number of wetland ponds (Fig. 3f) and total surface area of water present on the landscape has greatly increased (Liu and Schwartz 2012; Ballard et al. 2014). Rather than being the cause of the state shift in the southern PPR, it is instead likely that land-use change is exacerbating effects of the climate shift.
One of the most unexpected results of the recent hydrological changes has been an increase in salt mass in prairie-pothole-wetland ponds coinciding with increased water levels (Fig. 5a). We propose that this change is a key indicator of a state shift in the functioning of the region’s wetland ecosystems. The wetland continuum concept was proposed by Euliss et al. (2004) as a framework for better understanding biotic responses of wetlands that vary along continuous groundwater and atmospheric water gradients (Fig. 5b). The underlying presumption of the concept is that the biotic communities of a wetland are the result of abiotic conditions dependent on that wetland’s position in space and time along these two gradients. Additionally, it is presumed that the position along the vertical, dry to wet axis changes with time, whereas the position along the horizontal relationship to groundwater axis does not change (Euliss et al. 2004). However, the evidence that we present here brings into question this underlying assumption of the wetland continuum concept.
LaBaugh et al. (2016) showed that if high surface and groundwater levels persist, inflows to the wetland increase salt concentrations in that wetland to levels characteristic of drier, even drought-like conditions. These salinity increases result in shifts in the composition of biotic communities (McLean et al. 2016a). Thus, the biotic communities at set positions along the horizontal axis identified in the wetland continuum can shift to communities that are characteristic of other positions along that same axis while not varying along the vertical axis (Fig. 5b). If drought conditions return, it is possible that the wetlands will increase in salinity even more due to evaporative concentration. If this is the case, however, the semi-permanent ponded wetlands will not return to the previous position in the continuum but will have shifted to a new set of abiotic and biotic conditions. Therefore, in terms of determining biotic communities, the two-dimensional framework provided by Euliss et al. (2004) needs modification to describe biotic community response to novel climate conditions that result in sustained high-water levels. This suggestion presents additional evidence of a state shift affecting these wetlands, a shift beyond the conditions envisioned in the original wetland continuum.
In summary, we have presented abundant evidence that the recent wet climatic conditions sustained in the southern PPR since 1993 have been greater in magnitude and persisted longer than during any period in the last century, and as suggested by Winter and Rosenberry (1998), likely the last 500 years (Table S1; Fig. 2). This shift has distinctly altered the aquatic and terrestrial systems of the southern PPR (Fig. 3). The capacity for prairie-pothole-wetland biota in particular to return to their original state may be diminished due to increased salt loads (Fig. 5a). Our findings illuminate a need for the development of a generalized coupled hydro-geochemical model for management applications of prairie-pothole wetlands that can incorporate the functional changes associated with different ecohydrological states in order to more fully understand the ultimate influences of climate and ecosystem shifts.
References
Anteau MJ (2012) Do interactions of land use and climate affect productivity of waterbirds and prairie-pothole wetlands? Wetlands 32:1–9
Ballard T, Seager R, Smerdon JE, Cook BI, Ray AJ, Rajagopalan B, Kushnir Y, Nakamura J, Henderson N (2014) Hydroclimate variability and change in the Prairie Pothole Region, the “duck factory” of North America. Earth Interact 18:14.01–14.28
Blann KL, Anderson JL, Sands GR, Vondracek B (2009) Effects of agricultural drainage on aquatic ecosystems: a review. Crit Rev Environ Sci Technol 39:909–1001
Cox RR, Hanson MA, Roy CC, Euliss NH Jr, Johnson DH, Butler MG (1998) Mallard duckling growth and survival in relation to aquatic invertebrates. J Wildl Manag 62:124–133
Cressey RL, Austin JE, Stafford JD (2016) Three responses of wetland conditions to climatic extremes in the Prairie Pothole Region. Wetlands 36:357–370
Dai AG (2011) Characteristics and trends in various forms of the Palmer Drought Severity Index during 1900–2008. J Geophys Res-Atmos 116:1–26
Drummond MA, Auch RF, Karstensen KA, Sayler KL, Taylor JL, Loveland TR (2012) Land change variability and human–environment dynamics in the United States Great Plains. Land Use Policy 29:710–723
Dumanski S, Pomeroy JW, Westbrook CJ (2015) Hydrological regime changes in a Canadian Prairie basin. Hydrol Process 29:3893–3904
Dyke A, Prest V (1987) Late Wisconsinan and Holocene history of the Laurentide ice sheet. Géog Phys Quatern 41:237–264
Euliss NH, Labaugh JW, Fredrickson LH, Mushet DM, Laubhan MRK, Swanson GA, Winter TC, Rosenberry DO, Nelson RD (2004) The wetland continuum: a conceptual framework for interpreting biological studies. Wetlands 24:448–458
Finocchiaro RG (2016) Agricultural subsurface drainage tile locations by permits in North Dakota: U.S. Geological Survey data release. https://doi.org/10.5066/F7QF8QZW
Gascoigne WR, Hoag DL, Johnson RR, Koontz LM (Eds) (2013) Dynamics of land-use change and conservation in the Prairie Pothole Region of the United States—Environmental and economic implications with linkages to rural community well-being. U.S. Geological Survey Professional Paper 1800. Retrieved from http://pubs.usgs.gov/pp/1800/
Goldhaber MB, Mills C, Stricker CA, Morrison JM (2011) The role of critical zone processes in the evolution of the Prairie Pothole Region wetlands. Appl Geochem 26:S32–S35
Goldhaber MB, Mills CT, Morrison JM, Stricker CA, Mushet DM, LaBaugh JW (2014) Hydrogeochemistry of prairie pothole region wetlands: role of long-term critical zone processes. Chem Geol 387:170–183
Hamon WR (1961) Estimating potential evapotranspiration. J Hydraulics Div 87:107–120
Hayashi M, van der Kamp G, Rosenberry DO (2016) Hydrology of prairie wetlands: understanding the integrated surface-water and groundwater processes. Wetlands 36:237–254
Huang J, van den Dool HM, Georgakakos KP (1996) Analysis of model-calculated soil moisture over the United States (1931–1993) and applications to long-range temperature forecasts. J Clim 9:1350–1362
Huang BY, Banzon VF, Freeman E, Lawrimore J, Liu W, Peterson TC, Smith TM, Thorne PW, Woodruff SD, Zhang HM (2015) Extended reconstructed sea surface temperature version 4 (ERSST.v4). Part I: upgrades and intercomparisons. J Clim 28:911–930
Johnson WC, Poiani KA (2016) Climate change effects on prairie pothole wetlands: findings from a twenty-five year numerical modeling project. Wetlands 36:273–285
Karl TR (1983) Some spatial characteristics of drought duration in the United States. J Clim Appl Meteor 22:1356–1366
Karl T, Quinlan F, Ezell DS (1987) Drought termination and amelioration: its climatological probability. J Clim Appl Meteor 26:1198–1209
LaBaugh JW, Mushet DM, Rosenberry DO, Euliss NH, Goldhaber MB, Mills CT, Nelson RD (2016) Changes in pond water levels and surface extent due to climate variability alter solute sources to closed-basin prairie-pothole wetland ponds, 1979 to 2012. Wetlands 36:343–355
Leibowitz SG, Mushet DM, Newton WE (2016) Intermittent surface water connectivity: fill and spill vs. fill and merge dynamics. Wetlands 36:S323–S342
Liu GM, Schwartz FW (2012) Climate-driven variability in lake and wetland distribution across the Prairie Pothole Region: from modern observations to long-term reconstructions with space-for-time substitution. Water Resour Res 48:W08526
Mccauley LA, Anteau MJ, van der Burg MP, Wiltermuth MT (2015) Land use and wetland drainage affect water levels and dynamics of remaining wetlands. Ecosphere 6:1–22
McLean KI, Mushet DM, Renton DA, Stockwell CA (2016a) Aquatic-macroinvertebrate communities of prairie-pothole wetlands and lakes under a changed climate. Wetlands 36:423–435
McLean KI, Mushet DM, Stockwell CA (2016b) From “duck factory” to “fish factory”: climate induced changes in vertebrate communities of prairie pothole wetlands and small lakes. Wetlands 36:407–421
Menne MJ, Williams CN Jr, Vose RS (2009) The US Historical Climatology Network monthly temperature data, version 2. Am Meteorol Soc B 90:993–1007
Muggeo VMR (2008) Segmented: an R package to fit regression models with broken-line relationships. R News 8:20–25
Mushet DM (2016) Midcontinent prairie-pothole wetlands and climate change: an introduction to the supplemental issue. Wetlands 36:223–228
Mushet DM, Goldhaber MB, Mills CT, McLean KI, Aparicio VM, McCleskey RB, Holloway JM, Stockwell CA (2015) Chemical and biotic characteristics of prairie lakes and large wetlands in south-central North Dakota—effects of a changing climate. Scientific Investigations Report 2015–5126, Reston. Retrieved from https://pubs.er.usgs.gov/publication/sir20155126
Mushet DM, Rosenberry DO, Euliss Jr. NH, Solensky MJ (2016) Cottonwood Lake study area—water surface elevations. U.S. Geological Survey Data Release, Reston. https://doi.org/10.5066/F7707ZJ6
NOAA (2016) National Centers for Environmental information, Climate at a Glance: U.S. Time Series. Retrieved on December 19, 2016 from http://www.ncdc.noaa.gov/cag/
Palmer WC (1965) Meteorological drought. Research paper No. 45. U.S. Department of Commerce Weather Bureau, Washington
Pavelis GA (1987) Economic survey of farm drainage. In: Pavelis GA (ed) Farm drainage in the United States: history, status, and prospects. U.S. Gov. Print Office, Washington, D.C.
Pomeroy JW, Gray DM, Shook KR, Toth B, Essery RLH, Pietroniro A, Hedstrom N (1998) An evaluation of snow accumulation and ablation processes for land surface modelling. Hydrol Process 12:2339–2367
R Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Renton DA, Mushet DM, DeKeyser ES (2015) Climate change and prairie pothole wetlands: mitigating water-level and hydroperiod effects through upland management: U.S. Geological Survey Scientific Investigations Report 2015–5004, Reston. Retrieved from https://pubs.usgs.gov/sir/2015/5004/
Rosenberry DO (2003) Climate of the Cottonwood Lake area. In: Winter TC (ed) Hydrological, chemical, and biological characteristics of a prairie pothole wetland complex under highly variable climate conditions: the Cottonwood Lake area, east-central North Dakota: U.S. Geological Survey Professional Paper 1675, Reston. Retrieved from https://pubs.er.usgs.gov/publication/pp1675
Sando T (2015) Trends in North Dakota field water management. The Oxbow, pp 1–18. North Dakota State Water Commission, Bismark. Retrieved from: http://www.swc.nd.gov/info_edu/reports_and_publications/oxbow_articles.html
Shook K, Pomeroy J (2012) Changes in the hydrological character of rainfall on the Canadian prairies. Hydrol Process 26:1752–1766
Smith A, Stoudt J, Gallop J (1964) Prairie pothole wetlands and marshes. In: Linduska J (ed) Waterfowl tomorrow. U.S. Government Printing Office, Washington, DC, pp 39–50
Steen VA, Skagen SK, Melcher CP (2016) Implications of climate change for wetland-dependent birds in the Prairie Pothole Region. Wetlands 36:445–459
Stichling W, Blackwell SR (1957) Drainage area as a hydrologic factor on the glaciated Canadian prairies. IUGG Proceedings 111:365–376
Todhunter PE (2016) Mean hydroclimatic and hydrological conditions during two climatic modes in the Devils Lake Basin, North Dakota (USA). Lakes Reserv Res Manag 21:338–350
Todhunter PE, Fietzek-DeVries R (2016) Natural hydroclimatic forcing of historical lake volume fluctuations at Devils Lake, North Dakota (USA). Nat Hazards 81:1515–1532
Todhunter PE, Rundquist BC (2004) Terminal lake flooding and wetland expansion in Nelson County, North Dakota. Phys Geogr 25:68–85
USFWS (2016) Waterfowl population status, 2016. U.S. Department of the Interior, Washington, D.C.
van den Dool H, Huang J, Fan Y (2003) Performance and analysis of the constructed analogue method applied to U.S. soil moisture over 1981–2001. J Geophys Res Atmosp 108:1–16
Vecchia AV (2008) Climate simulation and flood risk analysis for 2008-40 for Devils Lake, North Dakota. US Geological Survey Scientific Investigations Report 2008-5011, Reston. Retrieved from: https://pubs.usgs.gov/sir/2008/5011/
Vicente-Serrano SM, Beguería S, Lorenzo-Lacruz J, Camarero JJ, López-Moreno JI, Azorin-Molina C, Revuelto J, Morán-Tejeda E, Sanchez-Lorenzo A (2012) Performance of drought indices for ecological, agricultural, and hydrological applications. Earth Interact 16:1–27
Winter TC (2003) Hydrological, chemical, and biological characteristics of a prairie pothole wetland complex under highly variable climate conditions : the Cottonwood Lake area, east-central North Dakota. US Geological Survey Professional Paper 2003-1675, Reston. Retrieved from: https://pubs.er.usgs.gov/publication/pp1675
Winter TC, Rosenberry DO (1998) Hydrology of prairie pothole wetlands during drought and deluge: a 17-year study of the cottonwood lake wetland complex in North Dakota in the perspective of longer term measured and proxy hydrological records. Clim Chang 40:189–209
Wright CK, Wimberly MC (2013) Recent land use change in the Western Corn Belt threatens grasslands and wetlands. Proc Natl Acad Sci USA 110:4134–4139
Acknowledgements
We thank Kyle McLean for feedback on figures and analysis, Natalie McKenna for copyediting, Matt Solensky for collecting and maintaining data from CLSA, and Christopher Mills and three anonymous individuals for providing their constructive reviews of our manuscript. Data from Cottonwood Lake Study Area wetlands are publically available through the Missouri Coteau Wetland Ecosystem Observatory (https://www.sciencebase.gov/catalog/item/52f0ffd9e4b0f941aa181fc6) and maintained through funding received from the U.S. Geological Survey’s Climate Research and Development Program. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US government.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(DOCX 293 kb)
Rights and permissions
About this article

Cite this article
McKenna, O.P., Mushet, D.M., Rosenberry, D.O. et al. Evidence for a climate-induced ecohydrological state shift in wetland ecosystems of the southern Prairie Pothole Region. Climatic Change 145, 273–287 (2017). https://doi.org/10.1007/s10584-017-2097-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10584-017-2097-7