
Abstract
Oceanic environmental variables derived from satellites are increasingly being used to predict ecosystem states and climate impacts. Despite the concerted efforts to develop metrics and the urgency to inform policy, management plans, and actions, few metrics have been empirically tested with field data for testing their predictive ability, refinement, and eventual implementation as predictive tools. In this study, the abilities of three variations of a thermal threshold index and a multivariate stress model (MSM) were used to predict coral cover and community susceptibility to bleaching based on a compilation of field data from Indian Ocean reefs across the strong thermal anomaly of 1998. Field data included the relative abundance of coral taxa 10 years before the large-scale temperature anomaly, 2 years after (1999–2000), and during the post-bleaching recovery period (2001–2005) were tested against 1) a multivariate model based on 11 environmental variables used to predict stress or environmental exposure (MSM), 2) estimates of the time until the current mean maximum temperature becomes the mean summer condition (TtT), 3) the Cumulative Thermal Stress (CTS) for the full satellite record, and 4) the 1998 Annual Thermal Stress (1998 ATS). The MSM showed significant fit with the post-1998 cover and susceptibility of the coral community taxa (r2 = 0.50 and 0.31, respectively). Temperature threshold indices were highly variable and had relatively weak or no significant relationships with coral cover and susceptibility. The ecosystem response of coral reefs to climatic and other disturbances is more complex than predicted by models based largely on temperature anomalies and thresholds only. This implies heterogeneous environmental causes and responses to climate disturbances and warming and predictive models should consider a more comprehensive multiple parameter approach.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Climate change has progressed sufficiently to enable testing of climate change predictions with empirical field data (Rahmstorf et al. 2007). Coral reefs have been an early victim of global and local environmental changes, with coral cover losses in the Indo-Pacific at 1–2 % per year (Bruno and Selig 2007; Precht and Aronson 2006) and some regions, such as the Caribbean, experiencing coral cover losses of 90 % (Gardner et al. 2003), of which most of this change was generated from the white-band disease in the early 1980s and successive bleaching events (Schutte et al. 2010). The breakdown of the coral-algal symbiosis, or coral bleaching, is a principal source of mortality and caused by the interactions between strong oceanographic temperature anomalies and other environmental factors (Brown et al. 2002; Carilli et al. 2009; Glynn 1996; Hoegh-Guldberg 1999). Bleaching encourages diseases (Bruno and Selig 2007; Harvell et al. 2002; McClanahan et al. 2009a; Miller et al. 2009) and promotes mortality of the most susceptible taxa (McClanahan 2014). However, differences in symbiont density, taxa, and coral coloration may interact with a variety of coral life history characteristics and environmental factors to mediate a coral’s response to temperature anomalies (Coles and Brown 2003; Fitt et al. 2001; McClanahan 2014; Mieog et al. 2009; van Woesik et al. 2012). Current understanding indicates a complex relationship between bleaching, mortality, and recovery (McClanahan 2004; Wilson et al. 2012) and there are strong regional differences in responses and recovery that further illustrate this complexity (Ateweberhan et al. 2011; Baker et al. 2008; Wilson et al. 2012).
The most cited and used indicators of climate change effects on coral reefs are temperature threshold indices for coral bleaching (Liu et al. 2003; Skirving et al. 2006), which have led to the suggestion that, as seawater temperatures rise beyond a local mean maximum level of stress, bleaching events will become more intense and more frequent, leading to coral death and declining coral populations (Donner 2009; Donner et al. 2005; Hoegh-Guldberg 1999; Sheppard 2003; Van Hooidonk et al. 2013). Recognition of a temperature-threshold link to bleaching was developed based on field observations of bleaching events and post hoc analyses of seawater temperature histories (Berkelmans 2002; Glynn 1993, 1996; Glynn and D'Croz 1990; Goreau and Hayes 1994). The temperature threshold concept and the interaction with ocean acidification and human resource use have been used subsequently to predict future climate impacts on coral reefs (Hoegh-Guldberg et al. 2007). However, coral bleaching may not always lead to coral death and is distinct from recovery, and thresholds developed to predict the presence of coral bleaching may not be de facto thresholds of coral mortality and associated coral reef degradation.
The bleaching threshold concept has not been fully tested and evidence for increasing acclimation and a rising threshold are increasingly being recognized (Guest et al. 2012; Logan et al. 2014; Maynard et al. 2008a; McClanahan 2014). Intrinsic differences among sites, species, and the combinations of symbiotic relationship between algal symbionts and coral hosts have led to nuanced conceptual models suggesting potential coral response elasticity to temperature rise (Ateweberhan and McClanahan 2010; Baker et al. 2004; Hughes et al. 2003). Indeed, some empirical tests of bleaching threshold indices have suggested weaknesses in the methods, indicating the importance of other attenuating factors such as historical temperature variations and coral species composition (McClanahan et al. 2007b; van Hooidonk and Huber 2009a). Indices that incorporate multiple environmental variables and moving thresholds have been used to study bleaching responses and these efforts may improve the predictive ability of bleaching and coral mortality (Ateweberhan and McClanahan 2010; Logan et al. 2014; Maina et al. 2008; McClanahan et al. 2007b; Wooldridge 2009). In order to ascertain the functionality of coral stress metrics in informing decisions, there is a need to better understand the relationship between coral mortality and conventional temperature threshold indices, as well as whether alternative and multivariate indices could improve predictive power.
The largest restraint to model testing and refinement has been the paucity of empirical field data on important dependent measures, such as coral cover and taxonomic composition (McClanahan et al. 2007a, b, c). Empirical comparison among bleaching and stress indices is possible, due to well-documented strong temperature anomalies and regional variation in their magnitude (Goreau et al. 2000). This information along with coral bleaching responses, coral cover changes, and changes in community structure over large oceanographic regions provide a basis for testing existing stress models (Ateweberhan and McClanahan 2010; Bruno and Selig 2007; Cote et al. 2005; Graham et al. 2008; McClanahan et al. 2007a, b, c). For example, average oceanic SST’s in the Indian Ocean have increased by ~1 °C over the past 100 years (Heron et al. 2008; Ihara et al. 2008) while the intensity of the high frequency climate variability (driven by ENSO and Indian Ocean Dipole) has increased since the 1920s (Nakamura et al. 2009). The earliest reported coral bleaching in the Indian Ocean was observed from a single location in 1983 (Faure et al. 1984). However, considering the strength of the 1983 ENSO (Spencer et al. 2000), we think there were unreported strong bleaching occurrences in the western Indian Ocean. Since 1987 there have been multiple reports and observations, with numerous observations being immediately after 1998 and widespread occasional observations both before and afterwards (Oliver et al. 2009; Souter 2000).
Given the global importance of accurately predicting both the occurrence and the after effects of bleaching events, we use a recent compilation of Indian Ocean coral cover (Ateweberhan et al. 2011) and an updated evaluation of large-scale coral community composition (McClanahan et al. 2007c) to test the predictive strength of four climate stress indices. Indices include a multivariate stress model (Maina et al. 2008), a temperature threshold index, and two variations of accumulated thermal stress that represent a range of current thinking about climate change impacts on coral reefs. Our approach is based on the idea that unusual annual or cumulative inter-annual variation can have severe impacts when specific thresholds act as tipping points for changes in coral communities, coral cover, and ecological function. While these might be seen as rare stochastic events, warming of the climate system is “unequivocal” (Stocker et al. 2014), suggesting threshold-breaching anomalies will increase in the near future.
2 Methods
Community metrics
To explore the skill of multiple thermal stress indices, we examined the response of two community-level metrics, coral cover (as a percentage of hard coral among other types of substrate cover) and site-susceptibility to bleaching. Coral cover data were compiled from multiple published and unpublished sources, resulting in a database of ~2000 site-time combinations gathered between 1958 and 2005 (Ateweberhan et al. 2011). The main data sources were: CORDIO status reports (Souter 2000); ReefBase-Reef Check data (Tupper et al. 2011), CRCP/WCS Database (McClanahan 2014); (Jennings and Polunin 1996; Sheppard 1999; Wilkinson 2004); (Hardman et al. 2004); (Mohammed et al. 2000) and (Graham et al. 2006). For each study, hard coral cover data were extracted, and the date of the field survey and global position of the study sites entered into the database. Cover data in 1998 were collected immediately after the bleaching event, which occurred at the end of 1997 to early 1998 depending on the latitude of the site. Coral cover data were pooled into three disturbance phases, pre- (up to 1997), immediately after - (1998–2000), and post- (2001–2005) bleaching for analyses with stress metrics.
Coral bleaching site susceptibility (a coral community measure of each site’s susceptibility to bleaching), was determined from a series of targeted coral community surveys undertaken during bleaching events after the strong 1998temperature anomaly (McClanahan et al. 2007a). The relative abundance of each coral genus was multiplied by a scaled taxa-specific bleaching response, based on a large number of bleaching observations (>45,000 individual corals), and summing this weighted metric across all taxa. The community-level site-susceptibility index was therefore a measure of each site’s susceptibility to bleaching and was influenced by the history of disturbance at each location (for full details see (McClanahan et al. 2007b, c). This was deemed important as bleaching (and other disturbances) often eliminate the most susceptible taxa, thus coral bleaching site-susceptibility index accounts for the history of environmental stress (Darling et al. 2013). A declining susceptibility index reflects the increasing relative dominance of bleaching resistant taxa. In comparison to coral cover, site susceptibility data were available from a limited number of locations collected only between 2005 and 2008. Hence, the two response variables were analysed separately, using available corresponding stress indices.
Stress indices
The first index, the multivariate stress model (MSM), utilized SST data from NOAA at 4-km resolution and HADISST (1 × 1o resolution), photosynthetically active radiation (PAR) and chlorophyll-a from SEAWIFS, ultraviolet (UV) irradiance estimates from NASA’s TOMS, and surface currents from the OSCAR model (fuller details provided in (Maina et al. 2008)). The following 11 variables were used to develop the model: mean maximum temperature, temperature variability (coefficient of variation, CV), coefficient of temperature rate of rise (based on linear model), degree heating weeks (DHW), photosynthetic radiation (PAR), ultraviolet light, water column chlorophyll-a concentrations, wind speed, and zonal and meridional current speeds. These 11 variables were synthesized using fuzzy logic algorithms, where functions that represent the effects these variables on coral bleaching response were estimated from field studies, applied as best-fit mathematical functions, and weighted based on the strength of the environmental-bleaching relationships (Maina et al. 2008). This process made use of climate variables, coral bleaching experiments and monitoring data, and statistical analysis of ReefBase bleaching data for 1983–2005 but did not use these data to test its predictive ability. Two multivariate spatial models estimating the stress on coral reefs in the Western Indian Ocean were developed, but the differences in their predictive ability were small and predictions for stress model 1 (SM1; described by (Maina et al. 2008) were used. The model was tested against a smaller sample of field data and showed a moderate predictive ability of regional change in coral cover at a gross level immediately after 1998 (Maina et al. 2008).
A time-to-threshold index (TtT) was calculated based on a rate of SST rise established by the longer Hadley SST data and the mean temperatures from the more spatially resolved NOAA Climate Data Center data. Hadley SST global monthly mean time series data for 1950–2008 were downloaded from the United Kingdom Meteorological Office archives (http://hadobs.metoffice.com/hadsst2/data/download.html) and annual averages calculated and the SST rate of rise over the 59 years determined. Monthly-averaged SST data for 1985–2008, at 4-km resolution product of AVHRR Pathfinder Version 5 SST Project were downloaded from the NOAA Climate Data Center website (ftp://data.nodc.noaa.gov/pub/data.nodc/pathfinder). Mean maximum and mean summer SST were calculated for each pixel, the former being the average of the annual maximum for all years and the latter being the average of the warmest 3 months of the year. Hadley SST trends were re-sampled to 4-km resolution and the predicted time for summer mean temperatures to reach the present warmest month’s average was calculated by:
2.1 TtT = (mean maximum SST – mean summer SST)/ SST rate of rise
This index balanced the existing summer temperature variability of each location with regional trends; for example, locations with a larger range of temperatures during summer have more capacity to cope with a warming trend than sites with a lesser range of summer temperatures. Because the mean summer SST is based on the past 59 years it may apply well to sites reaching this threshold in the next 50 years but unlikely to sites with low rate of SST rise.
NOAA Coral Reef Watch metrics are frequently used to identify regions of thermal stress around the globe in near real-time. We examined the utility of three thermal indices derived from the Reef Watch for describing longer-term ecosystem responses to stressful events. These metrics are derived from satellite temperature measurements that have been used to monitor thermal stress on global coral reefs since late-2000 (Liu et al. 2003; Skirving et al. 2006). The widely-used Degree Heating Weeks (DHW) metric is a measure of thermal stress accumulated through time, calculated by integrating half-weekly HotSpot values that are at or above the bleaching threshold (i.e., mean monthly maximum) of 1 °C during the preceding 12-week period. The index has been widely associated with coral bleaching events.
The HotSpot metric provides a location-specific warm temperature anomaly based on the summer maximum climatology. The HotSpot analysis was extended to before 2000 using a retrospective HotSpot dataset from 1985 to 2006 that emulates current methodology (Eakin et al. 2009). Two thermal indices were developed from the extended HotSpot dataset. First, HotSpots were integrated for each year to give the total Annual Thermal Stress (ATS) and ATS 1998 was used as it represented the highest stress. This index is identical to the maximum DHW value except at locations where thermal events last longer than 12 weeks, such as the equatorial Indian Ocean and the Gulf of Aden. We used the sum of annual “high stress” values (DHW >8 °C-week; (Liu et al. 2003) as an index of Cumulative Thermal Stress (CTS), indicating the chronic exposure for each site through the 22-year dataset.
Data analyses
The MSM, CTS, 1998 ATS and TtT index estimates for locations were mapped and corresponding community susceptibilities were extracted for analysis for coral reef locations where coral cover and coral community susceptibility values were available (Figs. 1 and 2). The four indices were compared by pair-wise correlation to determine the level of interrelationship. Models for coral susceptibility and cover at three time periods (pre 1998 ENSO: up to1997, immediate post 1998: 1998–2000, and late post 1998: 2001–2005) were fit using generalized additive mixed models (GAM), using the GAM function in R version 3.2.2 (R Core Team 2014). GAM was adopted rather than linear regression to allow for possible non-linear effects of predictors on the response variable, without the necessity to define the functional form. Smooth terms were fit using a cubic spline basis with shrinkage and limiting the smoothing parameter to a maximum value of 5 to avoid over-fitting and ensure monotonic relationships (Wood 2006). Coral cover data were square-root transformed and assumptions of the analysis evaluated using residual plots - data met the requirements. Coral community susceptibility data obtained between 2005 and 2008 were used In addition, models were compared for predictability using Bayesian Information Criterion (BIC), which is best suited for comparing models based on different sample sizes (Wood 2006).

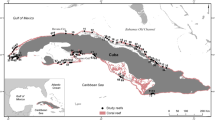
A map of the study area showing locations of (a) coral cover sites, and (b) community susceptibility to bleaching sites

Maps of the distribution of the (a) multivariate stress index map (SM1), (b) time-to-threshold index, (c) 1998 Annual Thermal Stress (ATS), and (d) Cumulative Thermal Stress (CTS, 1985-2008)
3 Results
The four indices were significantly correlated with each other (Table 1). The strongest association was between CTS and the 1998 ATS (r = 0.77); the latter term explained more than half of the variance in the 22-year time-series, indicating the significance of the 1998 stress event within the record.
Coral cover prior to 1998 was highly variable with a mean bottom cover of ~45 % (Fig. 3a). The northern Indian Ocean including Seychelles, Kenya, Maldives, and Chagos Islands had the greatest losses of coral cover after 1998 (Fig. 3b). The southern Indian Ocean, including Mozambique, South Africa, Madagascar, Reunion, and Mauritius, was less affected. Tanzania had the highest within country coral cover variation. Prior to 1998, coral cover showed a weak positive association with CTS (r2 = 0.03, Table 2a). Analysis of coral cover 2 years immediately after the 1998-bleaching event found coral cover was highly negatively correlated (r2 = 0.50) with the multivariate stress model (MSM) and to a lesser extent with the 1998 ATS (r2 = 0.03); CTS 1998–2000 (r2 = 0.05); and CTS 1985–2006 (r2 = 0.03). The strength of the multivariate stress model-coral cover relationship declined after 2000 due to the patchy recovery of coral cover in some countries, notably the Maldives and the Chagos Archipelago (r2 = 0.24 Fig. 3). Coral community susceptibility when fitted as a function of MSM and threshold models, showed a significant negative response pattern with the multivariate stress model (r2 = 0.31), and with two of the three temperature threshold models (i.e., 1998 ATS (r2 = 0.13); CTS 1985–2006 (r2 = 0.11); Fig. 3, Table 2b, A1). Comparisons of the two significant predictors of coral community susceptibility depict multivariate stress model as a stronger predictor.

Plots of multivariate stress index for the three coral cover time periods, (a) pre 1998, b) immediate after 1998 (1998–2000), (c) post bleaching (2001–2005), and (d) the coral community structure susceptibility. Lines and shaded regions represents non-linear model fits with + − 95 % confidence intervals
4 Discussion
Despite moderate associations among the indices, there were considerable differences in their predictive ability. The multivariate stress model (MSM) had substantially greater association with patterns of coral cover and community susceptibility after the 1998 disturbance while the temperature threshold indices had little predictive power. The acute thermal stress (ATS) index is very similar to the DHW metric, which had moderate to good success at predicting coral bleaching in various regional basins (McClanahan et al. 2007a) and the abundance of some bleaching-sensitive taxa (McClanahan et al. 2007b, c). There are, however, large differences among regions and taxa in their response to cumulative thermal stress measures (McClanahan et al. 2007b, c; van Hooidonk and Huber 2009a; Yee et al. 2008), which can be influenced by other temperature, light, and water flow variables (Donner et al. 2005; Maina et al. 2008; McClanahan et al. 2005). In addition, a considerable number of ecological processes that occur after bleaching, including acclimatization, mortality, and community change can influence any crude ecological measure of reef condition, such as coral cover (Maynard et al. 2008b).
The 1998 anomaly caused large-scale mortality across the Indian Ocean and these patterns were roughly associated with high thermal stress (Ateweberhan and McClanahan 2010). Despite the clear large-scale association between bleaching and ATS, this study shows that neither acute nor chronic thermal stress (CTS) metrics explained coral cover and the state of the coral community. Responses are probably more complicated, possibly dependent on local environmental histories, acclimatization, and the susceptibility of the coral community, particularly after there has been time for coral communities to respond to disturbance (McClanahan 2014). Despite well recorded long-term sea surface temperature variability in the tropics, the frequency of strong temperature variability events has increased in recent years (Gergis and Fowler 2009; Ihara et al. 2008; Nakamura et al. 2009). As background SSTs warm further, the likelihood of these ENSO and IOD events to raise temperatures above critical threshold has become higher and predictions indicate that repeated extreme events will exceed post disturbance recovery time (Donner et al. 2005; Sheppard 2003; Van Hooidonk and Huber 2012).
While metrics of annual accumulated thermal stress (e.g., DHW, ATS) may be useful for predicting bleaching (the design purpose of these indices) - particularly when the region, historical temperature, and taxonomic composition of the reefs are known or uniform- the CTS index did not predict the immediate post-1998 cover or community structure. The 1998 event was the most influential factor accounting for the negative relationships between ATS and MSM and coral cover. The MSM and community susceptibility relationships could only be tested for the post 1998 period but various lines of evidences suggest directional shifts in dominance towards bleaching tolerant taxa, depending on the intensity of the thermal anomaly (McClanahan 2014; McClanahan et al. 2007c).
The CTS index was weakly positively correlated with coral cover prior to 1998, indicating some benefits from occasional warm water. Temperature is frequently positively associated with coral reef growth and diversity up to a certain thermal limit of about 29–30 °C (Lough and Barnes 2000; Rosen 1971). The concern is, however, that a detrimental temperature threshold for many coral reef states and processes lies just beyond the beneficial point. The threshold for coral bleaching, derived from physiological experiments and field observations, indicates rapid declines in physiological processes at temperatures around 30–31 °C (Berkelmans and Willis 1999). Experiments where many potentially stressful variables are manipulated simultaneously show both negative and positive effects of multiple variables on coral condition (Anthony et al. 2007; Coles and Jokiel 1978). Consequently, while temperature is a critical factor, there are other relevant environmental variables that might interact and attenuate or induce losses in coral cover and community composition.
The time-to-threshold index (TtT) is an extension of the thermal stress metric that considers the seasonal temperature ranges and the moderate-term rising trends in SST. The index estimates the time until a locally significant warming occurs. Rates of SST rise and TtT are highly variable in the Indian Ocean region, with the TtT ranging from 25 to 500 years (Fig. 2b). Because of the current rapid changes in climate, the method is likely to have limited long term predictability. It is further complicated by the weak and non-linear relationships between accumulated thermal stress, SST rise, and possibly other environmental and ecological variables. This index did not show significant relationship with change in coral cover across the 1998-bleaching event, despite being a core element in models used to predict the future of coral reefs (Hoegh-Guldberg 1999; Sheppard 2003; Van Hooidonk et al. 2013).
Despite the concern that the SST rate of rise will lead to the demise of coral reefs, the relationship with bleaching has been equivocal and sometimes negative when the coolest regions were not in the analyses (Kleypas et al. 2008; Maina et al. 2008; McClanahan et al. 2007a; van Hooidonk and Huber 2009b). For example, DHWs in 1998 were neither linear or positively related to the rate of SST rise along the East African coastline with the highest DHW regions having a moderate SST rise (McClanahan et al. 2007a). Some Tanzanian and north-western Madagascan reefs located in high SST rise areas had low mortality and rapid recovery of corals (Maina et al. 2008; McClanahan et al. 2007c, 2009b). North-eastern Madagascar had both a low rate of rise and low DHWs in 1998; but this is not true for northern Kenya, which had low SST rise but very high DHWs in 1998 (McClanahan et al. 2007a; 2009d). SST rise is one of the variables in the MSM but was weakly negatively related to bleaching intensity and not strongly weighted in the model (Maina et al. 2008). However, this relationship could change as SST increases become more dominant in the future and determine DHW values. Climate disturbance models with rolling acclimations have been developed and this may help to improve their predictability of coral bleaching (Logan et al. 2014).
The TtT index is conceptually appealing and has therefore received considerable attention for predicting the future of coral reefs (Donner et al. 2005; Hoegh-Guldberg 1999; Sheppard 2003). Our analyses show that the index is not, so far, empirically supported in the western Indian Ocean region and, perhaps, cannot be tested until a future time when summer temperatures consistently exceed the maximum tolerance threshold. Nevertheless, warming is sufficiently underway that patterns, albeit weak, should have emerged. There is a need for more empirical research to evaluate relationships, even in the early stages of change (Rahmstorf et al. 2007). To be more predictive, we suggest that climate change indices and models should include more variables known to have both direct and indirect relationships with coral bleaching and mortality. The multivariate stress model weighed historical temperature variability highly and negatively and may be one reason for the better fits with coral cover after 1998.
Inclusion of numerous relevant environmental variables that could influence the susceptibility, resistance, tolerance of corals to thermal stress is likely to explain the success of the MSM. Each variable contributes only a small portion of the total weight of the model (Maina et al. 2008), making it difficult to attribute clear cause and effect to a particular variable. Consequently, this complexity may make the MSM index less appealing as causation is difficult to assign but, based on its predictive power, it currently appears to be the most realistic model. Nevertheless, even with 11 variables the MSM had only moderate predictive ability. Consequently, the considerable amount of unexplained variance may be uncovered when the influence of these and other variables are resolved and better understood. Certainly, local environment, habitat, and fisheries management factors contribute to a considerable amount of the total variation (McClanahan 2014).
The relationships suggest that predictions of future impacts of climate warming by the temperature threshold and MSM scenarios are considerably different. The threshold indices project a widespread and global-scale loss of reefs based on SST rate of rise as conditions exceed their limits (Hoegh-Guldberg et al. 2007; Sheppard 2003; van Hooidonk et al. 2014). In contrast, the MSM projects multiple and spatially heterogeneous responses dependent on the spatial heterogeneity of these and other variables and their interactions (Fig. 2a). Where as slowing and stopping anthropogenic climate change is the single long-term management solution for reef persistence, the MSM predicts considerable spatial and temporal variability in the destructive effects of the warming. The MSM model can, therefore, be useful in identifying reef areas based on the heterogeneous vulnerability to extreme anomalous events (McClanahan et al. 2009c). The model can help prioritize conservation and management actions to reduce local stress effects while global efforts to reduce warming are underway.
References
Anthony KRN, Connolly SR, Hoegh-Guldberg O (2007) Bleaching, energetics, and coral mortality risk: effects of temperature, light, and sediment regime. Limnol Oceanogr 52:716–726
Ateweberhan M, McClanahan TR (2010) Relationship between historical sea-surface temperature variability predicts climate change-induced coral mortality in the western Indian Ocean Mar. Pollut Bull 60:964–970
Ateweberhan M, McClanahan TR, Graham NAJ, Sheppard CRC (2011) Episodic heterogeneous decline and recovery of coral cover in the Indian Ocean. Coral Reefs 30:739–752
Baker AC, Starger CJ, McClanahan TR, Glynn PW (2004) Coral reefs: corals’ adaptive response to climate change. Nature 430:741
Baker AC, Glynn PW, Riegl B (2008) Climate change and coral reef bleaching: an ecological assessment of the long-term impacts, recovery trends and future outlooks. Estuar Coast Shelf Sci 80:435–471
Berkelmans R (2002) Time-integrated thermal bleaching thresholds of reefs and their variation on the Great Barrier Reef. Mar Ecol-Prog Ser 229:73–82
Berkelmans R, Willis BL (1999) Seasonal and local spatial patterns in the upper thermal limits of corals on the inshore Central Great Barrier Reef. Coral Reefs 18:219–228
Brown BE, Dunne RP, Goodson MS, Douglas AE (2002) Experience shapes the susceptibility of a reef coral to bleaching. Coral Reefs 21:119–126
Bruno JF, Selig ER (2007) Regional decline of coral cover in the Indo-Pacific: timing, extent, and subregional comparisons. PLoS One 2:e711
Carilli JE, Norris RD, Black BA, Walsh SM, McField M (2009) Local stressors reduce coral resilience to bleaching. PLoS One 4:e6324
Coles SL, Brown BE (2003) Coral bleaching - capacity for acclimatization and adaptation. Adv Mar Biol 46:183–223
Coles SL, Jokiel PL (1978) Synergistic effects of temperature, salinity and light on the hermatypic coral Montipora verrucosa. Mar Biol 49:187–195
Cote IM, Gill JA, Gardner TA, Watkinson AR (2005) Measuring coral reef decline through meta-analyses. Phil Trans R Soc 360:385–395
Darling ES, McClanahan TR, Cote IM (2013) Life histories predict coral community disassembly under multiple stressors. Glob Change Biol 19:1930–1940
Donner SD (2009) Coping with commitment: projected thermal stress on coral reefs under different future scenarios. PLoS One 4:e5712
Donner SD, Skirving WJ, Little CM, Oppenheimer M, Hoegh-Guldberg O (2005) Global assessment of coral bleaching and required rates of adaptation under climate change. Glob Change Biol 11:2251–2265
Eakin C, Lough J, Heron S (2009) Climate variability and change: monitoring data and evidence for increased coral bleaching stress. In: Coral Bleaching. Springer, pp 41–67
Faure G, Guillaume M, Payri C, Thomassin BA, Vanpraet M, Vasseur P (1984) Massive bleaching and death of corals in the Mayotte Reef ecosystem (SW Indian-Ocean) Comptes Rendus De L Academie Des Sciences Serie III -Sciences De La Vie-Life Sciences 299:637–642
Fitt WK, Brown BE, Warner ME, Dunne RP (2001) Coral bleaching: interpretation of thermal tolerance limits and thermal thresholds in tropical corals. Coral Reefs 20:51–65
Gardner TA, Cote IM, Gill JA, Grant A, Watkinson AR (2003) Coral reef decline in the Caribbean - response. Science 302:392–393
Gergis JL, Fowler AM (2009) A history of ENSO events since A.D. 1525: implications for future climate change. Climate Change 92:343–387
Glynn PW (1993) Coral reef bleaching: ecological perspectives. Coral Reefs 12:1–17
Glynn PW (1996) Coral reef bleaching: facts, hypotheses and implications. Glob Change Biol 2:495–509
Glynn PW, D'Croz L (1990) Experimental evidence for high temperature stress as the cause of El Nino-coincident coral mortality. Coral Reefs 8:181–191
Goreau TJ, Hayes RL (1994) Coral bleaching and ocean “hot spots. Ambio 23:176–180
Goreau T, McClanahan T, Hayes R, Strong A (2000) Conservation of coral reefs after the 1998 global bleaching event. Cons Biol 14:5–15
Graham NAJ, Wilson SK, Jennings S, Polunin NVC, Bijoux JP, Robinson J (2006) Dynamic fragility of oceanic coral reef ecosystems. Proc Natl Acad Sci U S A 103:8425–8429
Graham NAJ, McClanahan TR, MacNeil MA(2008) Climate warming and the ocean-scale integrity of coral reef ecosystems. PLoS One 3:e30309
Guest JR, Baird AH, Maynard JA (2012) Contrasting patterns of coral bleaching susceptibility in 2010 suggest an adaptive response to thermal stress. PLoS One 7:e33353
Hardman ER, Meunier MS, Turner JR, Lynch TL, Taylor M, Klaus R (2004) The extent of coral bleaching in Rodrigues, 2002. J Nat Hist 38:3077–3089
Harvell CD, Mitchell CE, Ward JR, Altizer S, Dobson AP, Ostfeld RS, Samuel MD (2002) Ecology - Climate warming and disease risks for terrestrial and marine biota. Science 296:2158–2162
Heron S, Skirving W, Eakin M (2008) Global climate change and coral reefs: reef temperature perspectives covering the last century. In: Wilkinson C (ed) Status of coral reefs of the world: 2008. Global coral reef monitoring network and reef and rainforest. Research Centre, Townsville, p 296
Hoegh-Guldberg O (1999) Climate change, coral bleaching and the future of the world’s coral reefs. Mar Freshwater Res 50:839–866
Hoegh-Guldberg O et al (2007) Coral reefs under rapid climate change and ocean acidification. Science 318:1737–1742
Hughes TP, Baird AH, Bellwood DR (2003) Climate change, human impacts, and the resilience of coral reefs. Science 301:929–933
Ihara C, Kushnir Y, Cane MA (2008) Warming trend of the Indian Ocean SST and Indian Ocean Dipole from 1880 to 2004. J Clim 21:2035–2046
Jennings S, Polunin NV (1996) Impacts of fishing on tropical reef ecosystems. Ambio:44–49
Kleypas JA, Danabasoglu G, Lough JM (2008) Potential role of the ocean thermostat in determining regional differences in coral bleaching events. Geophysical Res Lett 35:L03613
Liu G, Hogrefe C, Rao ST (2003) Evaluating the performance of regional-scale meteorological models: effect of clouds simulation on temperature prediction. Atmos Environ 37:1425–1433
Logan CA, Dunne JP, Eakin CM, Donner SD (2014) Incorporating adaptive responses into future projections of coral bleaching. Glob Change Biol 20:125–139
Lough JM, Barnes DJ (2000) Environmental controls on growth of the massive coral Porites. J Exp Mar Biol Ecol 245:225–243
Maina J, Venus V, McClanahan TR, Ateweberhan M (2008) Modelling susceptibility of coral reefs to environmental stress using remote sensing data and GIS models in the western Indian Ocean. Ecol Model 212:180–199
Maynard JA, Anthony KRN, Marshall PA, Masiri I (2008a) Major bleaching events can lead to increased thermal tolerance in corals. Mar Biol 155:173–182
Maynard JA, Baird AH, Pratchett MS (2008b) Revisiting the Cassandra syndrome; the changing climate of coral reef researcher. Coral Reefs 27:745–749
McClanahan TR (2004) The relationship between bleaching and mortality of common corals. Mar Biol 144:1239–1245
McClanahan TR (2014) Decadal coral community reassembly on an African fringing reef. Coral Reefs 33:939–950
McClanahan T, Maina J, Moothien-Pillay R, Baker A (2005) Effects of geography, taxa, water flow, and temperature variation on coral bleaching intensity in Mauritius. Mar Ecol Prog Ser 298:131–142
McClanahan TR, Ateweberhan M, Muhando CA, Maina J, Mohammed MS (2007a) Effects of climate and seawater temperature variation on coral bleaching and mortality. Ecol Monogr 77:503–525
McClanahan TR, Ateweberhan M, Ruiz Sebastian C, Graham NAJ, Wilson SK, Bruggemann JH, Guillaume MMM (2007b) Predictability of coral bleaching from synoptic satellite and in situ temperature observations. Coral Reefs 26:695–701
McClanahan TR, Ateweberhan M, Sebastian CR, Graham NAJ, Wilson SK, Guillaume MMM, Bruggemann JH (2007c) Western Indian Ocean coral communities: bleaching responses and susceptibility to extinction. Mar Ecol Prog Ser 337:1–13
McClanahan TR, Weil E, Maina J (2009a) Strong relationship between coral bleaching and growth anomalies in massive Porites. Glob Change Biol 15:1804–1816
McClanahan TR, Muthiga NA, Maina J, Kamukuru AT, Yahya S (2009b) Changes in northern Tanzania coral reefs over a period of increased fisheries management and climatic disturbance. Aquat Conserv 19:758–771
McClanahan TR, Cinner JE, Graham NAJ, Daw TM, Maina J, Stead SM, Wamukota A, Brown K, Venus V, Polunin NVC (2009c) Identifying reefs of hope and hopeful actions: Contextualizing environmental, ecological, and social parameters to respond effectively to climate change. Cons Biol 23:662–671
McClanahan TR, Ateweberhan M, Omukoto J, Pearson L (2009d) Recent seawater temperature histories, status, and predictions for Madagascar’s coral reefs. Mar Ecol Prog Ser 380:117–128
Mieog JC, Olsen JL, Berkelmans R, Bleuler-Martinez SA, Willis BL, van Oppen MJH (2009) The roles and interactions of symbiont, host and environment in defining coral fitness. PLoS One 4:e6364
Miller J, Muller E, Rogers C (2009) Coral disease following massive bleaching in 2005 causes 60 % decline in coral cover on reefs in the US Virgin Islands. Coral Reefs 28:925–937
Mohammed MS, Muhando C, Machano H (2000) Assessment of coral reef degradation in Tanzania: results of coral reef monitoring 1999. In: Souter D, Obura D, Linden O (eds) Coral reef degradation in the Indian Ocean. CORDIO, Vasteras, Sweden, pp 35–42
Nakamura N, Kayanne H, Iijima H, McClanahan TR, Behera SK, Yamagata T (2009) Mode shift in the Indian Ocean climate under global warming stress. Geophysical Res Lett 36:23. doi:10.1029/2009GL040590
Oliver JK, Berkelmans R, Eakin CM (2009) Coral bleaching in space and time. In: van Oppen M, Lough JM (eds) Coral bleaching: patterns, processes, causes and consequences. Springer Ecological Studies, Berlin, pp 21–39
Precht WF, Aronson RB (2006) Death and resurrection of Caribbean coral reefs: a palaeoecological perspective. In: Cote IM, Reynolds JD (eds) Coral reef conservation. Cambridge University Press, Cambridge, pp 40–77
Rahmstorf S, Cazenave A, Church JA, Hansen JE, Keeling RF, Parker DE, Somerville RCJ (2007) Recent climate observations compared to projections. Science 316:709
Rosen BR (1971) The distribution of reef coral genera in the Indian Ocean. Symp Zool Soc Lond 28:263–299
Schutte V, Selig E, Bruno J (2010) Regional spatio-temporal trends in Caribbean coral reef benthic communities. Mar Ecol Prog Ser 402:115–122
Sheppard CR (1999) Coral decline and weather patterns over 20 years in the Chagos Archipelago, Central Indian Ocean Ambio:472–478
Sheppard CRC (2003) Predicted recurrences of mass coral mortality in the Indian Ocean. Nature 425:294–297
Skirving W, Liu G, Strong AE, Liu C, Sapper J, Arzayus F (2006) Extreme events and perturbations of coastal ecosystems. In: remote sensing of aquatic coastal ecosystem processes. Springer, Dordrecht, pp 11–25
Souter D (2000) Coral reef degradation in the Indian Ocean. Status reports and project presentation 2000. CORDIO & SAREC, Stockholm University, Sweden
Spencer T, Teleki KA, Bradshaw C, Spalding MD (2000) Coral bleaching in the southern Seychelles during the 1997–1998 Indian Ocean warm event. Mar Pollut Bull 40:569–586
Stocker TF, Qin D, Plattner GK (2014) Climate change 2013: the physical science basis intergovernmental panel on climate change, working group I contribution to the IPCC fifth assessment report (AR5). Cambridge Univ Press, New York
Team RC (2014) R Foundation for Statistical Computing. Vienna, Austria. 2012 R: A language and environment for statistical computing
Tupper M, Tan M, Tan S, Radius M, Abdullah S (2011) ReefBase: a global information system on coral reefs
van Hooidonk R, Huber M (2009a) Quantifying uncertainties in predictions of coral bleaching. Coral Reefs 28:579–587
van Hooidonk R, Huber M (2009b) Equivocal evidence for a thermostat and unusually low levels of coral bleaching in the Western Pacific Warm Pool. Geophysical Res Lett 36:6
Van Hooidonk R, Huber M (2012) Effects of modeled tropical sea surface temperature variability on coral reef bleaching predictions. Coral Reefs 31:121–131
Van Hooidonk R, Maynard J, Planes S (2013) Temporary refugia for coral reefs in a warming world. Nat Clim Change 3:508–511
van Hooidonk R, Maynard JA, Manzello D, Planes S (2014) Opposite latitudinal gradients in projected ocean acidification and bleaching impacts on coral reefs. Glob Change Biol 20:103–112
van Woesik R, Franklin EC, O'Leary JK, McClanahan TR, Klaus JS, Budd AF (2012) Hosts of the Plio-Pleistocene past reflect modern-day coral vulnerability. P Roy Soc B-Biol Sci 279:2448–2456
Wilkinson CR (2004) Status of coral reefs of the world 2004: summary. Australian Institute of Marine Science, Townsville
Wilson SK, Graham NAJ, Fisher R (2012) Effect of macroalgal expansion and marine protected areas on coral recovery following a climatic disturbance. Cons Biol 26:995–1004
Wood S (2006) Generalized additive models: an introduction with R. CRC press
Wooldridge S (2009) Water quality and coral bleaching thresholds: formalising the linkage for the inshore reefs of the Great Barrier Reef, Australia. Mar Pollut Bull 58:745–751
Yee SH, Santavy DL, Barron MG (2008) Camparing environmental influences on coral bleaching across and within species using clustered binomial regression. Ecol Model 218:162–174
Acknowledgments
Research was supported by the Wildlife Conservation Society through grants from the Western Indian Ocean Marine Science Association (WIOMSA) Marine Science for Management (MASMA) program, the World Bank Targeted Research Group on Coral Bleaching, and John D. and Catherine T. MacArthur Foundation. S. Heron provided NOAA data and A. MacNeil made useful comments to an earlier draft of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
McClanahan, T.R., Maina, J. & Ateweberhan, M. Regional coral responses to climate disturbances and warming is predicted by multivariate stress model and not temperature threshold metrics. Climatic Change 131, 607–620 (2015). https://doi.org/10.1007/s10584-015-1399-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10584-015-1399-x