
Abstract
Climate change is predicted to alter the rainfall regime in the Eastern Mediterranean Basin: total annual rainfall will decrease, while seasonal and inter-annual variation in rainfall will increase. Such changes in the rainfall regime could potentially lead to large-scale changes in aboveground net primary productivity (ANPP) in the region. We conducted a data-driven evaluation of herbaceous ANPP along an entire regional rainfall gradient, from desert (90 mm MAR [Mean Annual Rainfall]) to Mesic-Mediterranean (780 mm MAR) ecosystems, using the largest database ever collated for herbaceous ANPP in Israel, with the aim of predicting consequences of climate change for rangeland productivity. This research revealed that herbaceous ANPP increases with increasing rainfall along the gradient, but strong dependence on rainfall was only apparent within dry sites. Rain Use Efficiency peaks at mid-gradient in Mediterranean sites without woody vegetation (560 and 610 mm MAR). Inter-annual coefficients of variation in rainfall and herbaceous ANPP decrease along the rainfall gradient up to ca. 500 mm MAR. Climate change is more likely to affect herbaceous ANPP of rangelands in the arid end of the rainfall gradient, requiring adaptation of rangeland management, while ANPP of rangelands in more mesic ecosystems is less responsive to variation in rainfall. We conclude that herbaceous ANPP in most Mediterranean rangelands is less vulnerable to climate change than generally predicted.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Climatic conditions in the Mediterranean Basin have become drier in the recent decades (Kafle and Bruins 2009), and this trend is expected to continue, with a decrease of up to 20 % in total annual rainfall by the year 2050 (Golan-Angelko and Bar-Or 2008; Black 2009; Evans 2009). Changes in rainfall (rather than temperature or CO2) constitute the most important component of climate change in the Mediterranean Basin, where water is the primary limiting resource (Fay et al. 2000; Miranda et al. 2011; Shafran-Nathan et al. 2012). Intra-seasonal variation in rainfall is expected to become more extreme, with fewer, but more intense rainfall events within a season. Likewise, inter-annual variation in rainfall is also expected to increase, with very wet years alternating with drought years. Since rainfall directly affects plant growth, changes in the rainfall regime should have serious consequences for ecosystem functions and services, such as primary productivity (Fay et al. 2003; Nippert et al. 2006; Köchy et al. 2008) and vegetation diversity (Noy-Meir 1973; Weltzin et al. 2003; Köchy et al. 2008). This is particularly important for ecosystems that provide vital services to mankind, such as rangelands (Weltzin et al. 2003; Nippert et al. 2006; Swemmer et al. 2007; Tietjen and Jeltsch 2007; Köchy et al. 2008). Climate change may affect both rangeland vegetation quality and productivity, the two primary factors affecting the carrying capacity of the rangeland for livestock (Tietjen and Jeltsch 2007; Köchy et al. 2008).
Climate change effects are expected to be stronger in arid and semi-arid ecosystems, where herbaceous ANPP depends strongly on both total rainfall (Noy-Meir 1973) and its variability (Le Houérou et al. 1988). In mesic ecosystems (≥500 mm MAR), however, water is not necessarily the limiting factor for herbaceous ANPP. Yet, these systems may still be affected by changes in the rainfall regime due to its interaction with other limiting factors such as temperature, nutrient availability, and soil texture and depth (Seligman and van Keulen 1989; Zaady 2005). Moreover, species thriving in mesic ecosystems often exhibit specific mechanisms that reduce vegetation response to rainfall and rainfall variability, such as substantial soil seed banks (Harel et al. 2011) and accumulation of reserves in below-ground storage organs (Sternberg et al. 2003), both of which may buffer changes in vegetation composition.
The complex response of primary productivity to rainfall across different ecosystems complicates efforts to develop predictions regarding climate change in different regions, and their validation in the field (Huxman et al. 2004). We addressed this complexity by using recent data to examine current relationships between herbaceous ANPP and rainfall, as a basis for formulating predictions about the effects of climate change, at least in the short term, on rangeland productivity. A theoretical analysis of ANPP-rainfall relationships was conducted previously for Mediterranean rangelands (Seligman and van Keulen 1989), and found that while soil depth affected herbaceous ANPP within the range 130–530 mm MAR, the effect of annual rainfall was stronger at the low end of the range, while soil fertility was more important at the high end. To date, empirical validation of this analysis is limited to one research site in northern Israel (Henkin et al. 1998). The current paper presents the first attempt to predict effects of climate change along an entire regional rainfall gradient, encompassing a 9-fold increase in annual rainfall within a short geographical range (240 km), using a comprehensive database of empirical data. We chose ten rangeland sites along the rainfall gradient in Israel for which long-term data on herbaceous ANPP were available. We focused on the herbaceous vegetation due to its importance for livestock, and asked the following questions:
-
1.
To what extent can inter-annual variation in herbaceous ANPP be explained by a) total annual rainfall and b) inter-annual variation in rainfall? and
-
2.
How will climate change affect herbaceous ANPP (i.e. forage production) in desert and Mediterranean rangelands?
2 Methods
2.1 Description of research sites
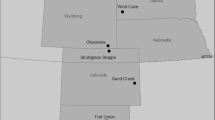
We used data from ten research sites along the rainfall gradient in Israel, from Sede Boqer (Site 1) in the south (ca. 90 mm MAR) to En Ya’aqov (Site 10) in the north (ca. 780 mm MAR). Figure 1 presents a map of the study sites; their characteristics are summarized in Online Resource 1. Herbaceous vegetation is an important component at each site, although its cover relative to that of woody vegetation decreases from south to north. At Karei Deshe (Site 8) there is no perennial woody vegetation, rather up to 40 % of the vegetation consists of hemicryptophytes (Sternberg et al. 2003), including Hordeum bulbosum L., a perennial grass which is the dominant palatable plant species at the site. Most sites have a long grazing history, by cattle in the north and by sheep and goats in the south. Nowadays grazing is conducted at some of the research sites, either for raising livestock or as a management tool for fire prevention (Online Resource 1). Throughout this paper, we refer to Sites 1–4 as ‘dry sites’ and Sites 5–10 as ‘mesic sites’.

Location of the ten research sites on the rainfall map of Israel and the Israeli-occupied territories according to U.N. designations, with typical landscape photos. The map presents long-term average annual rainfall for the period 1981–2010 (kindly provided by the Israeli Meteorological Service)
2.2 Data collection
We chose sites for which long-term (9–23 years) data from monitoring of the aboveground herbaceous vegetation were available. At each site biomass sampling was conducted within an experimental framework that examined the response of the herbaceous biomass to various factors, such as grazing, removal of woody vegetation, mechanical disturbance to the soil, fertilization, seeding, addition or reduction of rainfall. Each experiment comprised several treatments with 2–8 replications per treatment, ranging in size from several square metres to tens of hectares in area. To represent the spatial heterogeneity of the vegetation, several replicate samples were harvested in each plot at each harvest date. The data used in this study were collected by different people at the different sites, since each experiment was conducted over a time period specific to each site.
In order to enable comparisons across all experimental sites, we used data from control plots only. At Sites 8 and 10 we deviated slightly from this policy and included data from plots with limited exposure to grazing, assuming that since grazing at both sites occurred after seed dispersal, its effect on biomass production in the following season was minimal. At the low-to-moderate grazing intensities used, trampling may actually promote biomass production by damaging woody vegetation (Site 10 only) and creating microsites for germination (Eckert et al. 1986; Winkel and Roundy 1991), while compaction effects may be short-lived (Weigel et al. 1990). At all sites we used biomass data collected in the spring (March–May, depending on specific site conditions) at the peak of the growing season (when standing biomass reaches its maximum) which is a good measure of Aboveground Net Primary Productivity (ANPP) in herbaceous systems. In total, the database on which this paper is based is the largest ever amassed in Israel, including 31,831 quadrats and spanning the period 1988–2011.
2.3 Vegetation sampling method
Herbaceous biomass was harvested in quadrats of 20 × 20 cm at some sites, and quadrats of 25 × 25 cm at other sites, with replication according to the experimental design at each site. Quadrats were thrown randomly within the sampling areas of the experimental plots. At sites with presence of shrubs, sampling areas excluded distances within 40 cm of a shrub, in order to avoid direct shrub effects on the herbaceous vegetation (Arnon et al. 2007). All biomass samples were dried before weighing, and biomass was expressed in grams of dry matter per square metre.
The sites differed in the harvesting method: at some (Sites 3, 6, 7, 8 and 10), vegetation was harvested 2–3 cm above ground level using secateurs, while at others (Sites 1, 2, 4, 5 and 9), plants were pulled carefully from the ground (with or without first cutting the roots 1 cm below the soil surface) and roots were removed before weighing.
Since such differences did not allow us to use the raw data in analyses involving different sites, we developed a method to calibrate the measurements from the different sites. To this end, we conducted paired vegetation sampling at all sites during the 2010/11 season, at peak biomass (March–May). At each site we chose ca. 20 homogeneous vegetation patches representing the range of herbaceous productivity at the particular site. In each patch we set down two quadrats of the size used at that site. In one quadrat the vegetation was sampled according to the site-specific method, while in the other the vegetation was cut at a height of 1 cm above the soil surface. Based on these sample pairs, we conducted orthogonal regressions—designed for situations in which experimental error is present in both of the variables being analyzed (Fuller 1987)—and normalized the local data (for all years) to obtain results that could be compared among sites. The correlation coefficients for the orthogonal regressions ranged between 0.82 and 0.96 among the sampled sites, with the exception of Site 5, where it was lower, but still highly significant (r = 0.66; p < 0.001).
2.4 Rainfall data
We collected rainfall data from the experimental sites or nearest meteorological station. For most sites, we were able to obtain daily rainfall data, from which we calculated the annual totals used here based on a hydrological year starting 1 September.
2.5 Rain use efficiency
For each site we calculated Rain Use Efficiency (RUE) as the ratio between ANPP and MAR (Le Houérou et al. 1988), expressed in g m−2 mm−1, in order to assess the efficiency of biomass production along the rainfall gradient. Theoretically, RUE should increase with MAR, as the proportion of effective rain (for plant growth) increases with decreasing aridity (Le Houérou 1984), until other environmental factors limit ANPP. However, RUE is also highly dependent on soil and vegetation characteristics and conditions, and therefore may not necessarily respond linearly with rainfall.
2.6 Variability indicators
We calculated coefficients of variation (CV) for both annual rainfall and herbaceous ANPP in each site, as indicators of the variability of the rainfall regime and of herbaceous vegetation productivity. The Production to Rain Variability Ratio (PRVR) for each site was then calculated as the ratio between the inter-annual CV of ANPP and that of rainfall. PRVR is usually greater than unity in dry ecosystems, due to the high dependence of ANPP on MAR, while in mesic ecosystems PRVR tends to be closer to, or even slightly below, unity, because ANPP is less influenced by changes in total rainfall (Le Houérou et al. 1988) and is buffered by other biotic and abiotic factors.
2.7 Statistical analyses
We conducted linear regression of herbaceous ANPP on MAR for all sites combined, separate linear regressions of herbaceous ANPP on annual rainfall for each site, and linear regression of CVANPP on CVrainfall. To test whether the PRVR values of the different sites were significantly different from unity we first tested the significance of the difference between the linear regression of CVANPP on CVrainfall and the 1:1 line (intercept = 0; slope = 1). As this difference was significant, we then tested the significance of the largest residual (Site 1) from the 1:1 line against the t-distribution. This residual was significantly different from zero, therefore Site 1 was deemed to have a PRVR value different from unity. In order to determine whether PRVR of additional sites was different from unity, we removed Site 1 and repeated the above process until the regression line was no longer significantly different from the 1:1 line. Regression analyses were conducted in JMP 7.0.2 (SAS Institute Inc., Cary, USA); significance of residuals was determined using the TDIST function in Microsoft Excel, using n − 2° of freedom.
3 Results
MAR increases along the rainfall gradient from south to north (Fig. 2). Trends in ANPP along the rainfall gradient are less clear (Fig. 2), due to large differences in ANPP between sites receiving similar amounts of rain (e.g. Site 5 versus Site 8). ANPP increases significantly with increasing MAR, from the arid and semi-arid sites in the south to the wetter sites in the north (Fig. 2; r 2 = 0.45; p = 0.034), but with increasing scatter in the MAR–ANPP relationship towards the wet end of the gradient (from Site 5 northwards). Indeed, when yearly data for each site are examined separately (Fig. 3), a significant and positive relationship between ANPP and MAR emerges for the southern, dry Sites 1–4, while in the mesic sites, this relationship is absent, except in Site 8.

Herbaceous ANPP (mean ± se) as a function of MAR (mean ± se) in ten research sites along the rainfall gradient in Israel. The slope of the regression between herbaceous ANPP and MAR and the significance of the slope (* = p < 0.05) as well as the value of r 2 appear next to the regression line. Sites are ordered according to geographical location from south to north: 1 = Sede Boqer; 2 = Shaked Park; 3 = Lehavim; 4 = Lahav; 5 = Matta; 6 = Ramat Hanadiv; 7 = Ofer; 8 = Karei Deshe; 9 = Yechiam; 10 = En Ya’aqov

Herbaceous ANPP as a function of annual rainfall for each of the ten research sites along the rainfall gradient in Israel. The slopes of the regressions between herbaceous ANPP and rainfall, the significance of the slope (* = p < 0.05; ** = p < 0.01; *** = p < 0.001; ns = not significant) and the coefficient of determination appear below the name of each site
Rain Use Efficiency (RUE) averaged 0.391 ± 0.067 g m−2 mm−1 across all sites. If we examine the general relationship between RUE and rainfall from a broad perspective, we see that RUE for 6 out of 10 sites was fairly constant, falling within the range 0.262–0.480 g m−2 mm−1 (Fig. 4). RUE in the remaining four sites (1, 6, 8 and 10) fell outside the 95 % confidence interval (0.240–0.542): whereas Sites 6 and 8 had very high RUE (0.677 and 0.803 g m−2 mm−1, respectively), RUE at Site 10 was relatively low (0.209 g m−2 mm−1), and the lowest value was recorded at Site 1 (0.100 g m−2 mm−1).

Rain Use Efficiency (RUE; mean ± se) as a function of mean annual rainfall (MAR; mean ± se) in ten research sites along the rainfall gradient in Israel. Sites are ordered according to geographical location from south to north: 1 = Sede Boqer; 2 = Shaked Park; 3 = Lehavim; 4 = Lahav; 5 = Matta; 6 = Ramat Hanadiv; 7 = Ofer; 8 = Karei Deshe; 9 = Yechiam; 10 = En Ya’aqov
At the dry end of the gradient, the inter-annual CVs of both rainfall and herbaceous ANPP were relatively high, and decreased markedly from Site 1 northwards to the middle of the gradient (Fig. 5a), with a much wider range of values, and steeper decline along the rainfall gradient for herbaceous ANPP than for rainfall. At this point, both CVs are in the range 20–25 %, and remain at this level up to the mesic end of the gradient. The relationship between the two CVs (Fig. 5b) was positive and highly significant (r 2 = 0.85; p = 0.0001), crossing the 1:1 line at the mesic end of the gradient. Consequently, the Production to Rain Variability Ratio (PRVR – Online Resource 2) was greater than 1 at the dry end of the gradient, decreasing towards 1 mid-gradient. In order to fit the CVANPP versus CVrainfall regression to the 1:1 line we needed to omit Sites 1, 2 and 3, where PRVR was significantly different from 1 (Table 1; PRVR = 2.43, 1.96 and 1.57 for Sites 1, 2 and 3, respectively).

Inter-annual coefficient of variation (CV) of rainfall and herbaceous ANPP in ten research sites along the rainfall gradient in Israel. a CV of rainfall (blue) and herbaceous ANPP (red) as a function of mean annual rainfall (MAR). b Linear regression of CV of herbaceous ANPP on CV of rainfall, including the 1:1 line for interpretive purposes. The slope of the regression, the significance of the slope (*** = p < 0.001) and the coefficient of determination appear next to the regression line
4 Discussion
4.1 Trends in herbaceous ANPP
Our analysis indicates that MAR is the limiting factor for herbaceous ANPP in Israel’s rangelands, at the geographical scale of the rainfall gradient. Indeed, if we remove the highly productive Sites 6 and 8 from the linear regression in Fig. 2, the relationship becomes quite strong, with an r 2 of 0.81. The pattern of Rain Use Efficiency (RUE) at the different research sites adds a further level of complexity to the ANPP–MAR relationship along the gradient. RUE for 6 out of 10 sites was within the range 0.262–0.480 g m−2 mm−1, typical levels for rangeland productivity in the scientific literature (Le Houérou et al. 1988; Paruelo et al. 1999; Huxman et al. 2004), however differences in RUE along the gradient were quite marked.
Sites 6 and 8 exhibited 2- to 3-fold higher RUE than the other sites, with values characteristic of highly productive grasslands with good range condition (Le Houérou et al. 1988). Sites 6 and 8 are different in that they comprise herbaceous vegetation only, whereas vegetation in other mesic sites (5, 7, 9, 10) includes a perennial shrub component, which competes with the herbaceous vegetation for light and soil resources, and limits its growth (Online Resource 1; Holzapfel et al. 2006). Thus it can be argued that more water and solar radiation are available for herbaceous plant growth at Sites 6 and 8 since there is no competition with woody plants for these resources. In addition, herbaceous perennial species (hemicryptophytes, geophytes, grasses) which are more productive than annual species, have greater cover at Sites 6 and 8 compared to the other mesic sites. These two factors increase the potential for higher herbaceous ANPP at Sites 6 and 8, hence high RUE.
Apart from the effect of vegetation composition (Paruelo et al. 1999; Bai et al. 2008), RUE has also been shown to be strongly affected by rangeland condition and soil characteristics (Noy-Meir 1973; Le Houérou 1984), such that relatively high ANPP may be recorded in arid ecosystems on sandy soils with good vegetation condition, or conversely, low ANPP could be obtained for an ecosystem in a high rainfall zone if the vegetation is in poor condition (Le Houérou 1984; Le Houérou 1996). The large increase in RUE from Site 1 to Site 2 could be related to the deep loessial soil at Site 2 which significantly increases the amount of water available for plant growth. Conversely, it is possible that the relatively low RUE at Site 10 (0.209 g m−2 mm−1), despite high MAR, results from intensive summer grazing for fire prevention. The exceptionally low RUE in Site 1 (0.10) is associated with low plant density, low production potential, and high evaporative demand (Noy-Meir 1973; Paruelo et al. 1999).
Paruelo et al. (1999) evaluated world-wide RUE patterns in grasslands and found a recurring pattern of vegetational constraints at low MAR and biogeochemical constraints at high MAR, while at intermediate MAR (ca. 475 mm), both vegetational and biogeochemical constraints were relatively low resulting in higher RUE compared to wetter or drier conditions. The results of the current study do not fit this trend, probably due to the fact that most of our research sites are not herbaceous grasslands senso stricto, rather they include a woody component which is actually the dominant vegetation component in most of our mesic sites.
The strong relationship between herbaceous ANPP and MAR within each of the dry sites, where herbaceous ANPP is very low, is in agreement with previous studies in other arid and semi-arid regions around the world (Le Houérou et al. 1988; Huxman et al. 2004; Bai et al. 2008). Therefore we can unequivocally conclude that rainfall is the primary factor limiting growth in the dry sites (Noy-Meir 1973). In contrast, the lack of relationship between herbaceous ANPP and MAR in five of the six mesic sites indicates that other factors besides the total amount of rainfall limit herbaceous ANPP in these sites. Five of the six mesic sites (Sites 5–7, 9–10) are characterized by shallow Terra Rossa soils that developed over Cenomanian-Turonian chalk and limestone bedrock, which is low in phosphorus (P) (Singer 2007; Online Resource 3), an important macronutrient limiting plant growth in the region (Henkin et al. 2010). Previous studies at Site 10 (Henkin et al. 1998; Henkin et al. 2010) emphasized the P-limitation to vegetative growth at this site. This P-limitation probably restricts ANPP in wet years, thus limiting the response of herbaceous ANPP to rainfall. In contrast, Site 8 is characterized by relatively deep basaltic soil that developed over basaltic rock which is rich in P (Online Resource 3). Therefore, in this site, P probably does not limit growth, thus enabling higher mean ANPP and a positive ANPP–MAR relationship.
Previous studies have examined the relationships between variation in rainfall and in herbaceous ANPP, assuming that spatial and temporal variation in rainfall has a much higher potential effect than average conditions (Le Houérou et al. 1988; Swemmer et al. 2007; Tietjen and Jeltsch 2007). Here we used coefficients of variation (CV) of both rainfall and herbaceous ANPP to describe their relative variation among years. The decrease in the CVs of both herbaceous ANPP and rainfall to 20–25 % as rainfall increases to 500–600 mm MAR reflects the decrease in climatic variability at the mesic end of the gradient. The higher Production to Rain Variability Ratio (PRVR) in the dry sites (Fig. 5b), as observed in other arid regions around the world (Wiegand et al. 2004; Nippert et al. 2006), reflects the strong effect of small changes in rainfall between years on herbaceous ANPP in dry regions. In our study sites, low ANPP and the predominance of annual species with low seed dormancy (Harel et al. 2011) lead to the high dependence of herbaceous ANPP on rainfall. In contrast, above 500 mm rainfall, CV of both herbaceous ANPP and rainfall are relatively stable, and PRVR is close to 1. The higher cover of perennial herbaceous species in the mesic sites may contribute to reducing relative inter-annual variation in herbaceous ANPP at these sites. These species can store reserves in underground storage organs (e.g. bulbs, corms) in above-average rainfall years for growth during dry years (Sternberg et al. 2003; Shafran-Nathan et al. 2012). In addition, Harel et al. (2011) showed that seed dormancy of herbaceous annuals is higher at the mesic end of our studied rainfall gradient (specifically, in Site 9), and the development of a long-term seed bank contributes to buffering of herbaceous ANPP variation between years.
4.2 Effects of climate change
Climate change models predict a decrease of up to 20 % in annual rainfall and an increase in its inter-annual variation in the eastern Mediterranean region (Golan-Angelko and Bar-Or 2008; Black 2009; Evans 2009). We can estimate the potential decrease in herbaceous ANPP in response to reduced rainfall, using the regressions in Fig. 3, assuming that the relationships between herbaceous ANPP and rainfall hold within the range of exploration. A 20 % reduction in rainfall will therefore lead to reductions in herbaceous ANPP of 40 % for the arid site (Site 1), 34 %, 31 % and 16 % for the semiarid sites (Sites 2–4) and 12 % for the Mediterranean site (Site 8). These values are within the range of the present variation in herbaceous ANPP, however they show that lower rainfall alone could have a significant effect on herbaceous ANPP.
Both reduced rainfall and higher inter-annual variability in rainfall have the potential to detrimentally affect herbaceous ANPP in arid and semi-arid sites, with significant reductions in ANPP as well as marked increases in ANPP variability (Figs. 3 and 5). However, any analysis examining possible effects of changes in the rainfall regime in these dry sites (Sites 1–4) must differentiate between short-term effects and long-term cumulative effects. Assuming a decreasing trend in annual rainfall, prevailing inter-annual variability in rainfall may, for some years, moderate trends of declining growth and reduced productivity due to the occurrence of above-average rainfall years, which allow replenishment of the limiting soil seed bank (Chesson 2000). However, ‘recovery’ of the vegetation in an above-average rainfall year does not guarantee high ANPP in the following year, due to the low presence of perennial herbaceous species, and restricted seed accumulation in the soil due to low seed dormancy (Harel et al. 2011). Previous models developed for dry grasslands in Israel showed that herbaceous ANPP in these grasslands is fairly resilient to reduced and more variable rainfall (Köchy et al. 2008; Shafran-Nathan et al. 2012). However, Köchy et al. (2008) emphasized the important role of grazing in such ecosystems, and the possible negative feedback cycle that could develop in the longer term, leading to potential collapse of the economic viability of dry rangeland livestock production (see also Ellis and Swift 1988).
Since we did not find a relationship between ANPP and MAR in most of the mesic sites, and accounting for the relatively low variability in both ANPP and MAR across all mesic sites, we would expect that productivity in these sites should not deviate in the short term from the current range in productivity in response to decreased annual rainfall. Herbaceous ANPP at Site 10 was previously shown to be limited rather by the interaction between rainfall and temperature (Henkin et al. 2010) and by biogeochemical constraints such as soil P (Henkin et al. 1998). Furthermore, variation in ANPP in mesic sites has been shown to be buffered by reserves in underground storage organs of perennial herbaceous species (Sternberg et al. 2003—Site 8) and by a more persistent soil seed bank (Harel et al. 2011—Site 9). If we assume a continuation of the existing random combination of above- and below-average rainfall years we would not expect any significant changes in average ANPP, due to the positive contribution of above-average rainfall years which allow replenishment of the soil seed bank of the annual species and recovery of herbaceous perennials (Sternberg et al. 2003; Shafran-Nathan et al. 2012). Thus the plant traits of the vegetation impart resilience to climate change in these mesic sites.
4.3 Limitations of the study
Two potentially important aspects of ANPP–MAR relationships were not addressed in our analysis: the effects of within-season rainfall variation and of consecutive drought years. Within-season rainfall variation comprises several factors, including frequency, intensity and duration of rain events, as well as the timing and length of the rainy season, and may. Using a model that manipulated within-season rainfall distribution, Shafran-Nathan et al. (2012) found that a delay in the commencement of the rainy season, or an early end to the rainy season may have serious consequences for vegetative growth. Similarly, in a manipulation experiment, Fay et al. (2003) found that changes in the temporal distribution of rainfall along the season had a greater impact on herbaceous ANPP than did a 30 % reduction in MAR. Conversely, other rainfall manipulation experiments found remarkable resilience of semi-arid ecosystems to changes in temporal distribution of rainfall along the season (Bates et al. 2006; Miranda et al. 2011). Indeed, 10 years of rainfall manipulations in two of the sites used in the current study (Sites 4, 5) did not lead to any response in productivity of the herbaceous vegetation (Kigel et al. unpublished data). The interaction between within-season rainfall distribution and temperature may also play a role in explaining ANPP–MAR relationships, especially within mesic sites. In a previous study at Site 10, Henkin et al. (1998) showed that temperature was a limiting factor for vegetative growth early in the growing season. We are currently examining patterns of within-season rainfall variation and their relationships with herbaceous ANPP within all 10 sites along the gradient.
In this study we did not consider extreme conditions of consecutive drought years, simply because they were too rare to enable meaningful analysis. However, it is reasonable to expect that such conditions could occur with greater frequency as climate change progresses. Consecutive drought years could lead to mortality of herbaceous perennials and a depleted soil seed bank of the annual species, with a consequent reduction in productivity of the herbaceous vegetation, even in mesic sites.
4.4 Conclusions
In conclusion, MAR is a strong driver of herbaceous ANPP along the desert–mesic Mediterranean rainfall gradient. However, while ANPP increases strongly with MAR at the dry end of the gradient, herbaceous ANPP is limited at the mesic end, possibly by the interaction of rainfall and temperature, and/or biogeochemical constraints. There was no clear trend in RUE along the gradient—this index was affected more by vegetation and soil conditions than by rainfall per se. Within sites, herbaceous ANPP is tightly coupled to annual rainfall in the dry sites, while in mesic sites, species traits such as underground storage organs and a persistent seed bank probably buffer variation in herbaceous ANPP between years. The interaction of reduced rainfall and increased rainfall variability has the potential to negatively affect herbaceous ANPP, especially in the dry sites, requiring adaptation of rangeland management. However, models and empirical studies have shown that the semi-arid and mesic rangelands along the studied rainfall gradient exhibit resilience to climate change. We conclude that Mediterranean rangelands are less vulnerable to climate change than generally predicted.
References
Arnon A, Ungar ED, Svoray T, Shachak M, Blankman J, Perevolotsky A (2007) The application of remote sensing to study shrub-herbaceous relations at a high spatial resolution. Israel J Plant Sci 55:73–82
Bai Y, Wu J, Xing Q, Pan Q, Huang J, Yang D, Han X (2008) Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau. Ecology 89:2140–2153
Bates JD, Svejcar T, Miller RF, Angell RF (2006) The effects of precipitation timing on sagebrush steppe vegetation. J Arid Environ 64:670–697
Black E (2009) The impact of climate change on daily precipitation statistics in Jordan and Israel. Atmos Sci Lett 10:192–200
Chesson P (2000) Mechanisms of maintenance of species diversity. Annu Rev Ecol Syst 31:343–366
Eckert RE Jr, Peterson FF, Meurisse MS, Stephens JL (1986) Effects of soil-surface morphology on emergence and survival of seedlings in Big Sagebrush communities. J Range Manage 39:414–420
Ellis JE, Swift DM (1988) Stability of African pastoral systems: alternate paradigms and implications for development. J Range Manage 41:450–459
Evans JP (2009) 21st century climate change in the Middle East. Clim Chang 92:417–432
Fay PA, Carlisle JD, Knapp AK, Blair JM, Collins SL (2000) Altering rainfall timing and quantity in a mesic grassland ecosystem: design and performance of rainfall manipulation shelters. Ecosystems 3:308–319
Fay PA, Carlisle JD, Knapp AK, Blair JM, Collins SL (2003) Productivity responses to altered rainfall patterns in a C4-dominated grassland. Oecologia 137:245–251
Fuller WA (1987) Measurement error models. John Wiley, USA
Golan-Angelko A, Bar-Or Y (2008) Israeli preparation for global climate change. Ministry for Environmental Protection, Office of the Chief Scientist [Hebrew], Jerusalem
Harel D, Holzapfel C, Sternberg M (2011) Seed mass and dormancy of annual plant populations and communities decreases with aridity and rainfall predictability. Basic Appl Ecol 12:674–684
Henkin Z, Seligman NG, Kafkafi U, Noy-Meir I (1998) ‘Effective growing days’: a simple predictive model of the response of herbaceous plant growth in a Mediterranean ecosystem to variation in rainfall and phosphorus availability. J Ecol 86:137–148
Henkin Z, Seligman NG, Noy-Meir I (2010) Long-term productivity of Mediterranean herbaceous vegetation after a single phosphorus application. J Veg Sci 21:979–991
Holzapfel C, Tielbörger K, Parag HA, Kigel J, Sternberg M (2006) Annual plant-shrub interactions along an aridity gradient. Basic Appl Ecol 7:268–279
Huxman TE, Smith MD, Fay PA, Knapp AK, Shaw MR, Loik ME, Smith SD, Tissue DT, Zak JC, Weltzin JF, Pockman WT, Sala OE, Haddad BM, Harte J, Koch GW, Schwinning S, Small EE, Williams DG (2004) Convergence across biomes to a common rain-use efficiency. Nature 429:651–654
Kafle HK, Bruins HJ (2009) Climatic trends in Israel 1970–2002: warmer and increasing aridity inland. Clim Chang 96:63–77
Köchy M, Mathaj M, Jeltsch F, Malkinson D (2008) Resilience of stocking capacity to changing climate in arid to Mediterranean landscapes. Reg Environ Chang 8:73–87
Le Houérou HN (1984) Rain use efficiency: a unifying concept in arid-land ecology. J Arid Environ 7:213–247
Le Houérou HN (1996) Climate change, drought and desertification. J Arid Environ 34:133–185
Le Houérou HN, Bingham RL, Skerbek W (1988) Relationship between the variability of primary production and the variability of annual precipitation in world arid lands. J Arid Environ 15:1–18
Miranda JD, Armas C, Padilla FM, Pugnaire FI (2011) Climatic change and rainfall patterns: effects on semi-arid plant communities of the Iberian southeast. J Arid Environ 75:1302–1309
Nippert JB, Knapp AK, Briggs JM (2006) Intra-annual rainfall variability and grassland productivity: can the past predict the future? Plant Ecol 184:65–74
Noy-Meir I (1973) Desert ecosystems: environment and producers. Ann Rev Ecol Evol Syst 4:25–51
Paruelo JM, Lauenroth WK, Burke IC, Sala OE (1999) Grassland precipitation-use efficiency varies across a resource gradient. Ecosystems 2:64–68
Seligman NG, van Keulen H (1989) Herbage production of a Mediterranean grassland in relation to soil depth, rainfall and nitrogen nutrition: a simulation study. Ecol Model 47:303–311
Shafran-Nathan R, Svoray T, Perevolotsky A (2012) The resilience of annual vegetation primary production subjected to different climate change scenarios. Clim Chang doi:10.1007/s10584-012-0614-2
Singer A (2007) The soils of Israel. Springer, Berlin
Sternberg M, Gutman M, Perevolotsky A, Kigel J (2003) Effects of grazing on soil seed bank dynamics: an approach with functional groups. J Veg Sci 14:375–386
Swemmer AM, Knapp AK, Snyman HA (2007) Intra-seasonal precipitation patterns and above-ground productivity in three perennial grasslands. J Ecol 95:780–788
Tietjen B, Jeltsch F (2007) Semi-arid grazing systems and climate change: a survey of present modeling potential and future needs. J Appl Ecol 44:425–434
Weigel JR, Britton CM, McPherson GR (1990) Trampling effects from short-duration grazing on tobosa-grass range. J Range Manage 43:92–95
Weltzin JF, Loik ME, Schwinning S, Williams DG, Fay PA, Haddad BM, Harte J, Huxman TE, Knapp AK, Lin G, Pockman WT, Shaw MR, Small EE, Smith MD, Smith SD, Tissue DT, Zak JC (2003) Assessing the response of terrestrial ecosystems to potential changes in precipitation. BioScience 53:941–952
Wiegand T, Snyman HA, Kellner K, Paruelo JM (2004) Do grasslands have a memory: modeling phytomass production of a semiarid South African grassland. Ecosystems 7:243–258
Winkel VK, Roundy BA (1991) Effects of cattle trampling and mechanical seedbed preparation on grass seedling emergence. J Range Manage 44:176–180
Zaady E (2005) Seasonal change and nitrogen cycling in a patchy Negev desert: a review. Arid Land Res Manag 19:111–124
Acknowledgments
To Irit Konsens (Sede Boqer, Lahav, Matta, Yechiam), Hagit Baram (Lehavim, Ofer), Itzhak Bezalel (Ofer), Eli Zaady, Daniel Barkai and Tanya Gendler (Shaked Park) and Liat Hadar and Mario Gutman (Ramat Hanadiv) for biomass data, and to Amir Arnon, Noam Ben Ari, Shilo Navon, Ezra Ben Moshe and Rafi Yonatan for assistance in the field and laboratory. To Mark Perel, Moty Peres, Alona Arie and David Bonfil for climatic data. To Hillary Voet for statistical advice and analysis. To Avner Furshpan and Noam Halfon from the Israeli Meteorological Service for help with preparing Fig. 1. To the GLOWA Jordan River project for supporting research at Sede Boqer, Lahav, Matta and Yechiam. This research is funded by the Fund of the Chief Scientist of the Israeli Ministry of Agriculture and Rural Development.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Online Resource 1
Physical and biotic characteristics of the study sites along the rainfall gradient, including long-term rainfall and temperature averages, as well as past and present grazing regimes. Temperature refers to annual means (mean minimum, mean, and mean maximum). Rainfall coefficient of variation (CV) is presented as percent. (PDF 34 kb)
Online Resource 2
Production to Rain Variability Ratio (PRVR) for ten sites along the rainfall gradient in Israel. Sites are ordered by PRVR. (PDF 14 kb)
Online Resource 3
Mean Annual Rainfall (MAR) and available soil phosphorus content (mean ± se) for six mesic sites in Israel representing 540–780 mm. Sites are ordered by geographical position from south to north. Data were not analyzed statistically due to large discrepancies in numbers of replicate soil samples at different sites, and differences in reliability of data. (PDF 19 kb)
Rights and permissions
About this article
Cite this article
Golodets, C., Sternberg, M., Kigel, J. et al. From desert to Mediterranean rangelands: will increasing drought and inter-annual rainfall variability affect herbaceous annual primary productivity?. Climatic Change 119, 785–798 (2013). https://doi.org/10.1007/s10584-013-0758-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10584-013-0758-8