
Abstract
Epinephrine (Epi), which initiates short-term responses to cope with stress, is, in part, stress-regulated via genetic control of its biosynthetic enzyme, phenylethanolamine N-methyltransferase (PNMT). In rats, immobilization (IMMO) stress activates the PNMT gene in the adrenal medulla via Egr-1 and Sp1 induction. Yet, elevated Epi induced by acute and chronic stress is associated with stress induced, chronic illnesses of cardiovascular, immune, cancerous, and behavioral etiologies. Major sources of Epi include the adrenal medulla and brainstem. Although catecholamines do not cross the blood–brain barrier, circulating Epi from the adrenal medulla may communicate with the central nervous system and stress circuitry by activating vagal nerve β-adrenergic receptors to release norepinephrine, which could then stimulate release of the same from the nucleus tractus solitarius and locus coeruleus. In turn, the basal lateral amygdala (BLA) may activate to stimulate afferents to the hypothalamus, neocortex, hippocampus, caudate nucleus, and other brain regions sequentially. Recently, we have shown that repeated IMMO or force swim stress may evoke stress resiliency, as suggested by changes in expression and extinction of fear memory in the fear-potentiated startle paradigm. However, concomitant adrenergic changes seem stressor dependent. Present studies aim to identify stressful conditions that elicit stress resiliency versus stress sensitivity, with the goal of developing a model to investigate the potential role of Epi in stress-associated illness. If chronic Epi over expression does elicit illness, possibilities for alternative therapeutics exist through regulating stress-induced Epi expression, adrenergic receptor function and/or corticosteroid effects on Epi, adrenergic receptors and the stress axis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Epinephrine (Epi) or adrenaline is a neurotransmitter and neurohormone rapidly released from the adrenal medulla into the portal circulation in response to stress. In contrast to corticosteroids (cortisol or corticosterone), it is thought to mediate short-term responses to stressors by initiating behavioral and physiological changes that permit an organism to confront the stressful stimulus and overcome it, the classic “fight or flight” mechanism described by Cannon and De La Paz in the early 1900s (Cannon and De La Paz 1911). Cannon introduced the notion of homeostasis, referring to mechanisms the body activates to maintain constancy, particularly on exposure to a stressor. Multiple cooperative feedback responses activate simultaneously or successively to ensure that constancy. Selye coined the term “general adaptation syndrome” as a further extension of events associated with stress and adaptation; the general syndrome evokes a systemic response to the particular stress while adaptation refers to the integrated responses of the biological system to counteract the stress. In current stress parlance, as proposed by McEwen, the stress responsiveness of an organism is considered in terms of allostasis and allostatic load (McEwen 2007). In contrast to homeostasis, which maintains physiological parameters within a well-defined, narrow range, allostasis refers to the broader range of variability in physiological parameters of the cardiovascular, metabolic, immune and central nervous systems, based on the circumstantial demands placed on them. Allostatic load, then, encompasses the price to be paid, whether real or perceived, beyond acceptable norms and as defined by McEwen and Gianaros, includes: (1) frequency and intensity of common stressor, (2) failure to habituate to a repeated single stressor leading to persistent elevation of a mediator of that stressor, (3) failure to terminate autonomic or neuroendocrine responses evoked to counter the stressor, and finally (4) failure to adequately meet the challenge of the stressor (McEwen and Gianaros 2010).
While Epi has been associated with short-term stress responses, cortisol is thought to mediate the long-term stress responses that can evoke illness when stress repeatedly occurs (Kvetnansky et al. 2009; McEwen and Gianaros 2010; Ulrich-Lai and Herman 2009). As such, its role in physiological and behavioral adaptation seems to be that of a modulator of the stress axis and its responses. Stress can vary in intensity and may be composed of physical, psychological, and environmental components. When an organism mounts the challenge to the stress, despite type or intensity, habituation can occur with repetition, and resiliency to the stress can ensue. However, if adaptation is inadequate to the challenge of the stress, increased stress sensitivity and chronic physical illness may occur instead.
Initially, the “stress axis” was equated with the “hypothalamic–pituitary–adrenal (HPA) axis.” From numerous investigations through the years to the present, it is clear that the former definition is far too limiting based on neuroanatomical, behavioral, and molecular evidence (Kvetnansky et al. 2009). Figure 1 shows a schematic of the “stress axis” as modified from Kvetnansky et al. 2009. Briefly, viscero- and somato-sensory stressful stimuli, via ascending spinal and medullary non-catecholaminergic and catecholaminergic neurons projecting to the brain integrative centers (limbic system, hypothalamus, and cerebral cortex) elicit activation of (1) the HPA cortical axis leading to the synthesis and release of cortisol or corticosterone from the adrenal cortex, (2) the HPA medullary axis leading to the synthesis and release of predominantly Epi and small amounts of norepinephrine (NE) into the portal circulation, (3) descending catecholaminergic neurons from the brainstem, which stimulate the pre-ganglionic sympathetic neurons-adrenal medulla leading again to Epi and NE release, and finally (4) pre-ganglionic sympathetic neurons-sympathetic ganglia (sympathetic nervous system, SNS), which pump large amounts of NE into the bloodstream. As described, the major source of peripheral Epi is the adrenal medulla and specifically, medullary chromaffin cells with large, medium-density vesicles containing the releasable pool of Epi. Chromaffin cells expressing NE can readily be distinguished from the latter by the presence of the NE containing small, dense core vesicles. Secondary to the chromaffin cells, Epi is synthesized in brainstem adrenergic neurons localized to the C1–C3 regions (Hokfelt et al. 1984).

Stress axis. Schematic of the central and peripheral components of the stress axis (see Kvetnansky et al. 2009, for comprehensive review)
Our interests focus on the potential role that the stress hormone Epi, which is itself regulated by stress via genetic control of its biosynthetic enzyme, phenylethanolamine N-methyltransferase (PNMT, EC 2.1.1.28) (Wong and Tank 2007; Wong et al. 2008, 2010), may play in initiating illness. While Epi historically has been associated with short-term stress responsiveness, it has also been linked to many long-term, stress-induced illnesses. What conditions and associated mechanisms of repeated, prolonged or unrelenting stress induce Epi expression to permit adaptation and provide resiliency to stress? To the contrary, how do those conditions and mechanisms differ from stressful conditions and mechanisms that lead to enduring elevation of Epi, maladaptation to stress, stress sensitivity, and illness? Understanding conditions and mechanisms distinguishing the latter from those of stress resiliency may provide inroads to therapeutics that minimize or prevent the potential for illness.
Epi and Stress-Induced Illness
The examples presented here are not meant to be all inclusive but to provide a sense of the important role and contribution of Epi to illnesses that are associated with and elicited by stress and linked to cardiovascular, immune, tumorigenic, and behavioral abnormalities. Certainly, there are many examples linking cardiovascular disease to Epi and stress. Repeated, intermittent elevation of Epi by stress increases blood flow throughout the circulatory system and causes vasoconstriction, thereby elevating systolic blood pressure over the short-term (Hoffman 2001). Epi also lowers diastolic blood pressure and via β2-receptor-mediated vasodilation may decrease blood pressure. However, these responses occur at low concentrations of Epi. At high concentrations of Epi, α-adrenergic receptors, present in greater abundance than β-adrenergic receptors, are activated to cause vasoconstriction and marked blood pressure elevation. Long-term stress has indeed been linked to hypertension but the contributions attributable to Epi remain to be determined. A recent report showed that stress exercise produces variable circulating Epi levels in humans, which associates with specific polymorphisms in the PNMT gene (Ji et al. 2008). The latter points to the important potential role of genetics in Epi-related cardiovascular disease. Emotional, physiological, and physical stresses have also been implicated in acute and chronic cardiomyopathies that induce cerebrovascular changes that can lead to sudden death. While the underlying pathophysiology remains to be elucidated, it is known that adrenergic-mediated changes, e.g., epicardial vasospasm, microvascular dysfunction, hyperdynamic contraction, are due to the direct action of the catecholamines on cardiomyocytes (Abraham et al. 2009; Dimsdale 2008; Esler 2010; Fineschi et al. 2010; Khouri and Imran 2009; Mausbach et al. 2010; Zeb et al. 2011). Abraham et al. (2009) described a “syndrome of stress cardiomyopathy,” which is elicited by intense emotional or physical stress, produces transient cardiac dysfunction and is thought driven by excessive sympathetic stimulation. Using coronary angiography, they further showed that intravenous administration of Epi or the β-receptor antagonist, dobutamine, induced similar symptoms, cardiac enzyme elevation, QTc interval prolongation and rapidly reversible cardiac dysfunction (Abraham et al. 2009). Recently, great concern has arisen with regards to placing individuals with cardiac abnormalities at risk in the case of travel and participation in high altitude activities since physiological adaptation may be problematic (Higgins et al. 2010). High altitude exposes individuals to hypoxic conditions; Epi, myocardial function, and pulmonary arterial pressure all increase, placing a greater load on the cardiovascular system. Tillinger et al. (2006) showed that mild hypoxic stress (8%) selectively induces PNMT gene expression in heart atria in rats (Tillinger et al. 2006). As PNMT protein was not determined, it would be interesting to ascertain if enzyme follows suit as PNMT mRNA and protein can be differentially regulated (Wong et al. 1992). Cardiorespiratory function is very sensitive to small changes in arterial oxygen, the availability of which is regulated through a family of transcription factors, the hypoxia-inducible factors (Peng et al. 2011). It has been demonstrated that in heterozygous HIF2α+/− mice, which exhibit partial HIF2α deficiency, the carotid body shows extreme sensitivity to hypoxia with the consequence of adverse cardiorespiratory changes. SNS activity, activated reflexively by hyper-stimulation of the carotid body, produces high plasma levels of NE. Further, in the adrenal medullae, mitochondrial electron transport is altered, consistent with oxidative stress. We have previously demonstrated that Epi is up-regulated by moderate hypoxic stress (5% O2) through activation of PNMT gene transcription with HIF1α acting as a “master switch” (Tai et al. 2009, 2010). However, the study of Peng et al. (2011) does not identify the role of Epi in the described cardiovascular changes, address whether mitochondrial electron transport dysfunction is focal to the predominant Epi producing cells in the medullae or if plasma Epi levels are elevated.
Epi is also an important modulator of the immune system through its control of neuroendocrine and immune interaction. Well known is the use of Epi in the treatment of anaphylaxis, sepsis, and bronchodilation for asthma. Its utility in the treatment of anaphylaxis and asthma depend on its vasoconstrictive effects mediated by β1-adrenergic receptor activation (Zhang et al. 2011) and bronchodilatory effects mediated by β2-adrenergic receptor activation (Ameredes 2011), respectively. In the case of anaphylaxis, activation of β2-adrenergic receptors on the pulmonary artery by circulating endogenous Epi produced in the adrenal, in fact, may be associated with the increased incidence and severity of anaphylaxis in patients (Zhang et al. 2011). Stress exercise is known to activate both the neuroendocrine and immune systems as well. Epi and NE increase during stress exercise. Monocytes show a rise in β2-adrenergic receptor expression prior to exercise onset, which wanes in both monocytes and granulocytes during the course of exercise. In contrast, β2-receptor expression in lymphocytes increases during the recovery period. Thus, there appears to be a temporal interaction in β2-adrenergic receptor expression in the neuroendocrine and immune systems prior to, during and after the physical stress of exercise (Fragala et al. 2011). Interestingly, acute immobilization (IMMO) stress produces a significant rise in β2-adrenergic receptor mRNA in the spleen of rats but a marked decline in mRNA and protein when IMMO is repeated. An even greater decline is observed in α2a and α2b receptor mRNA and protein with repeated IMMO. In all cases, the rise in receptor indices evoked by repeated stress appears to correlate with an increase in interleukin-6 (IL-6) and decrease in IL-10 mRNA (Laukova et al. 2010). Similarly, mood or fatigue appears to produce a diurnal variation in immune parameters and catecholamines. For example, IL-6 shows lesser nocturnal decline when mood is more negative or fatigue greater, and the latter is paired with a reduction in Epi decline. In contrast, no significant associations are observed for C-reactive protein, soluble tumor necrosis factor-receptor I or NE under comparable psychological conditions (Gaykema and Goehler 2011). Finally, the regulatory link between neuroendocrine hormones, such as Epi, and the immune system place the elderly at risk for inflammatory disease. As neuroendocrine function changes with age, the regulation of immune function is altered. Notably, chronic stress adversely affects neuroendocrine modulation of the immune system to exacerbate the possibility of immune disease (Heffner 2011).
Recently, Epi and stress have also been linked to cancer with demonstration that they facilitate progression of this disease, leading to poor prognosis. Cancer patients are exposed to physiological and psychological stress, which elevate corticosteroid and Epi secretion. It has been shown in F344 rats infected with CRNK-16 leukemia cells, followed by forced swim (FS) stress or Epi, corticosteroid or prostaglandin-E2 administration, that leukemia progression is enhanced, possibly through suppression of cell-mediated immunity. Moreover, β-adrenergic receptor antagonists or COX inhibitors slow progression and increase survival (Inbar et al. 2011). It has also been demonstrated that adrenergic modulation may facilitate tumor consolidation and growth to accelerate the progression of ovarian tumor cancer. Using a mouse model of human ovarian cancer, it was shown that restraint stress, under conditions that increase both Epi and NE, prevent cancerous cells from becoming apoptotic. Consequently, tumor growth increases, with concurrent activation of focal adhesion kinase (FAK). Stress hormones are thought to modulate FAK, which, in turn, facilitates tumor formation (Sood et al. 2010).
Finally, Epi and stress contribute to the development of psychiatric disorders. It has been hypothesized that Epi-expressing cells in the C1–C3 regions of the brainstem (Hokfelt et al. 1974) send afferent projections to pre-frontal cortex, midbrain and locus coeruleus (LC), releasing Epi, which can then interact with α1b-adrenergic receptors to modulate dopaminergic, noradrenergic, and serotonergic neurons. These regions have been linked to behavioral disorders, e.g., schizophrenia, bipolar disorder, depression, and are the targets for drugs efficacious in their treatment (Stone et al. 2003). Clinical studies have long associated elevated catecholamine levels with post-traumatic stress syndrome (PTSD). In a cohort of young adults examined over a 10-year period for trauma and PTSD, it was demonstrated that urinary Epi was significantly elevated in individuals with PTSD in the absence of major depressive disorder. In contrast, cortisol levels were unaltered and trauma alone did not lead to sustained cortisol and Epi elevation (Young and Breslau 2004). Moreover, co-morbid PTSD and major depression did present with higher cortisol levels. Thus, in this subset of PTSD patients, Epi, rather than corticosteroids, appears to associate with the illness. Recent studies are consistent with this earlier finding. Patients hospitalized with chronic PTSD were shown to have elevated levels of the Epi and NE metabolite, vanillylmandelic acid (VMA), decreased cortisol and elevated mean blood flow velocity in the circle of Willis vesicales (Dikanovic et al. 2011). Three weeks of treatment with anti-depressants or anxiolytics, however, significantly reduced the number of patients with elevated VMA and cerebral vasospasm. Interestingly, Debiec et al. (2011), using auditory fear conditioning in rats, showed that infusion of the β-adrenergic receptor agonist, isoproterenol, into the amygdala produced noradrenergic augmentation during retrieval of fear memories. By contrast, the antagonist, propranolol, blocked the enhancing effects of isopreterenol. As described earlier, peripheral Epi, via β-adrenergic receptors, activates NE release from the vagus, LC, and nucleus tracts solitarius (NTS) to stimulate the basal lateral amygdala (BLA). Thus, consistent with our hypothesis, Epi and noradrenergic impairment of extinction of conditioned fear may provide an inroad to understanding PTSD and alternative approaches for treatment (Debiec et al. 2011). However, the use of propanolol in treating PTSD patients appears controversial (Donovan 2010). Early research, which focused on memory acquisition and consolidation, showed inconsistent results with this drug. More recent work, including the aforementioned, which focuses on the role of propanolol on reconsolidation, suggests PTSD patients should not be withheld access from the palliative effects of Epi therapeutics. Finally, Epi abnormalities have also been detected in developmental disorders, such as autism, sampling from blood, plasma, or urine. Consistently, these studies demonstrate that Epi is significantly elevated in autistic children by comparison to their normal cohorts (Herault et al. 1993; Launay et al. 1987).
Collectively, these examples suggest a role of stress-induced Epi contributing to long-term stress-associated illness. Considering the overlap of stress and adrenergic circuitry, perhaps these occurrences seem a logical extension, rather than simply unrelated serendipity.
Epi Regulation by Stress
Steady-state levels of Epi, in the tissues where it is expressed, are governed by its relative rates of synthesis and degradation. This review on stress regulation of Epi focuses on the former, the effects of stress on its biosynthesis. Epi, the final catecholamine in the catecholamine biosynthetic pathway, is produced by N-methylation of the primary amino group on the aliphatic chain of NE (Fig. 2). S-adenosylmethionine is the methyl donor with methylation catalyzed by PNMT. Early studies in rats and mice demonstrated that Epi is regulated in vivo by stress through genetic control of its biosynthesis by PNMT (reviewed in Kvetnansky et al. 2009; Wong et al. 2010). In the rat adrenal medulla, IMMO stress increases both PNMT mRNA and protein expression with similar patterns and extent of change (Tai et al. 2007; Viskupic et al. 1994; Wong et al. 2002, 2004, 2008). mRNA and protein rise rapidly with 30 min of IMMO as effective as 2 h in maximizing their elevation. Repeated IMMO does not dampen the magnitude of induction of either PNMT mRNA or protein indicating that at least through 6 days of daily IMMO for 30 min or 2 h, the animals do not habituate to the stress. The latter is consistent with the earlier findings of Kvetnansky et al. (2002), which demonstrated that desensitization to 2 h of daily IMMO had not occurred even after 42 days of stress exposure (Kvetnansky et al. 2002).

Epinephrine biosynthesis from norepinephrine. Phenylethanolamine N-methyltransferase converts norepinephrine to epinephrine via transfer of a methyl group from S-adenosylmethionine (AdoMet) to the primary amino group on the aliphatic chain substituted onto the benzene ring
Research in rats indicates that critical transcriptional activators of PNMT (see Wong et al. 2010, for review) include early growth response gene-1 (Egr-1), specificity protein 1 (Sp1), glucocorticoid receptor (GR), glial cell missing-like factor (GCMl), activating protein 2 (AP-2), and cMyc-activating factor (MAZ) with response elements (REs) for all residing within the proximal −893 bp of the gene promoter sequence upstream of the site of transcription initiation (+1, Fig. 3) (Ebert et al. 1994, 1998; Ebert and Wong 1995; Her et al. 1999, 2003; Ross et al. 1990; Tai et al. 2002; Tai and Wong 2003; Wong et al. 1998). There are two REs for the immediate early gene transcription factor Egr-1 at −45 and −165 bp, two REs for the ubiquitous factor and transcription initiator of housekeeping genes and other genes lacking TATA and CAAT boxes, Sp1, at −48 and −168 bp, three REs for AP-2, a developmental factor critical for differentiation of neural crest-derived tissue, e.g., adrenal medulla, at −105, −587, and −674 bp, three glucocorticoid receptor REs (GREs) at −539, −759, and −773 bp (latter two overlap), one RE for a GCMl at −40 bp and an RE for the tumorigenic factor MAZ at −48 bp. While hypoxia did induce PNMT promoter activity as described earlier, a hypoxia response element (HRE), identified at −282 bp, showed no detectable interaction with HIF1α, even when levels of the latter where extremely elevated (see below). Stimulus-induced activation of all identified PNMT transcription activators, followed by their interaction with their cognate REs permits them to act independently and cooperatively to control PNMT gene expression. IMMO stress, in particular, transcriptionally activates the rat PNMT gene by inducing Egr-1 and Sp1 (Tai et al. 2002). Furthermore, stress induction of Egr-1 and Sp1 mRNA and protein (nuclear pool) and formation of protein–DNA binding complexes mirror the pattern of induction observed for PNMT mRNA and protein in the adrenal medulla.

Rat PNMT promoter. Schematic depicting binding sites for PNMT transcriptional activators
Other types of stress also activate PNMT, including cold (4°C), exercise and hypoglycemia (see Kvetnansky et al. 2009; Wong et al. 2010). However, the magnitude of change in PNMT expression is highly dependent on the type of stress (psychological, physical, or physiological) and its intensity. IMMO stress, a very robust physical and psychological stressor in rats, evokes marked changes in both adrenal PNMT mRNA and protein. In contrast, rats habituate to chronic cold stress, particularly when intermittent, with a loss of adrenal medullary PNMT responsiveness to the stress.
In addition, stress-induced changes in PNMT mRNA and protein, particularly in the case of repeated stress, may not necessarily be coordinated. Acute IMMO stress in rats produces similar patterns of change in adrenal PNMT mRNA and protein, irrespective of stress duration or time to kill. In contrast, if IMMO is repeated for 7 days, PNMT protein induction declines whereas mRNA elevation remains marked and sustained (Tai et al. 2007). The same inverse pattern of change in adrenal medullary mRNA and protein is evoked by glucocorticoid administration (Wong et al. 1992). Dexamethasone and the type II-specific glucocorticoid agonist, RU28362, induce PNMT mRNA as much as 10- to 20-fold while enzyme activity remains low or shows a rapid dose-dependent reduction. These findings indicate that PNMT can be both transcriptionally and post-transcriptionally regulated, and both types of control may contribute to stress responsiveness.
To delineate molecular mechanisms underlying stress-induced regulation of Epi through genetic control of PNMT, cell culture models have been invoked. For the most part, the model of choice are the PC12 cells, clonally established from a rat pheochromocytoma or adrenal medullary tumor (Greene and Tischler 1976). These cells are highly mutable, providing a composite, rather than single genotype/phenotype of cell, but are most akin to rat adrenal medullary chromaffin cells in response. Much information on stress regulatory mechanisms for PNMT has been garnered from examination of the effects of hypoxic stress on these cells. We have demonstrated that moderate hypoxia (5%) activates PNMT promoter-driven gene expression and induces endogenous PNMT mRNA and protein in these cells (Tai et al. 2002, 2009, 2010). Moderate hypoxia stresses the cells while further reduction in O2 switches cells toward the apoptotic pathway and certain cell death (Hopfl et al. 2004; Santra et al. 2008). The same two transcriptional activators of PNMT, Egr-1 and Sp1, which induced PNMT gene expression in the adrenal medulla when rats were exposed to IMMO stress, contribute to the rise in PNMT promoter-driven gene expression in response to hypoxic stress. As described above, hypoxia also induces a family of proteins called the hypoxia inducible factors. Mentioned earlier, HIF1α, an important member of that family, appears to act as a “regulatory on–off” switch for the proximal −893 bp of rat promoter based on the following evidence (Tai et al. 2009): (1) The HRE at −282 bp upstream of the transcription initiation site appears to be non-functional, (2) transfection studies with Egr-1 and Sp1 mutant constructs under conditions of hypoxia or over expression of HIF1α and Egr-1 and Sp1 siRNA knock-down do not result in activation of PNMT promoter-driven gene expression, and (3) HIF1α increases Egr-1 and Sp1 mRNA and protein expression.
In contrast to other reports, we have shown that in the PC12 cells, hypoxia activates the HIF1α gene to increase HIF1α protein. In rats, IMMO and the stress axis neurotransmitter, pituitary adenylate cyclase activating polypeptide (PACAP) also elevate HIF1α. In general, HIF1α is considered post-translationally regulated via protein degradation (Hopfl et al. 2004). The protein is constitutively expressed, and is functionally controlled through hydroxylation. Prolyl-hydroxylation in the oxygen-dependent domain of the protein leads to its ubiquitinylation and shunting to the proteosomes where it is degraded. Hydroxylation of an asparagine residue prevents interaction of HIF1α with cyclic AMP RE binding protein/p300 so that HIF1α is not activated and thus, cannot serve as a transcriptional regulator.
Signaling mechanisms underlying hypoxic activation of the PNMT gene have also been identified by examining the effects of selective signaling inhibitors on hypoxic activation of PNMT promoter-driven reporter gene expression in the PC12 cells and/or protein kinase A (PKA)- or phospholipase C (PLC)-deficient PC12 cells (Tai et al. 2010; Wong et al. 2010). Hypoxia initiates signaling through the two major bifurcating pathways, cAMP-PKA and PLC. Downstream, cAMP-PKA stimulates ERK1/2 mitogen-activated protein kinase (ERK1/2 MAPK) and p38 mitogen-activated protein kinase (p38 MAPK) while PLC induces phosphoinositide 3-kinase (PI3K) signaling. Further, we have evidence that protein kinase C (PKC) may interconnect the cAMP/PKA and PLC signaling pathways.
Molecular events associated with Epi regulation by stress reviewed in this section identify molecular targets, transcription factor mRNA and protein, PNMT mRNA and protein, signaling proteins/pathways, which can be used to distinguish how regulation of Epi biosynthesis, adrenergic receptors, and glucocorticoids may contribute to Epi’s role in stress-related illness as further elaborated below.
Stress Circuitry and Adrenergic Circuitry
The C1–C3 adrenergic nuclei in the brainstem and the chromaffin cells in the adrenal medulla are the major sources of Epi within the stress axis. Ascending and descending projections regulate CNS Epi production and mobilization, while adrenal medullary chromaffin cells regulate peripheral Epi production and mobilization.
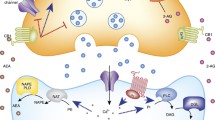
Circulating Epi is largely derived from the adrenal gland. It is established that catecholamines, for the most part, do not cross the blood–brain barrier (BBB) although limited passage of peripheral catecholamines to the CNS can occur at circumventricular sites, where the BBB is absent or structural deficiencies permit “leakage” of the neurotransmitters across the BBB (Kostrzewa 2007). Nonetheless, one route by which Epi can exert its effects centrally has been elegantly mapped out from evidence derived from behavioral studies in animal models and human testing (Cahill and Alkire 2003; Kvetnansky et al. 2009; McGaugh 2004; Mravec 2011; Roozendaal et al. 2009; Schreurs et al. 1986; Ulrich-Lai and Herman 2009). Briefly, Epi released from the adrenal medulla, can activate β-adrenergic receptors on vagal afferents to stimulate NE release from the vagus nerve (Fig. 4, as modified from Roozendaal et al. 2009). In turn, NE can activate the NTS and LC leading to NE release from the NTS and LC and stimulation of the BLA (Roozendaal et al. 2009). Together, adrenal gland, vagus nerve, NTS, LC, and BLA comprise the learning experience circuitry. BLA further activates the neocortex and hippocampus, caudate and other brain regions (memory consolidation circuitry). Reciprocal interaction between neocortex and the latter brain regions permits modulation of memory consolidation. Glucocorticoids, synthesized and released from the adrenal cortex, further modulate the activity of NTS, LC, BLA, neocortex, hippocampus, caudate nucleus and these other brain regions associated with memory consolidation and thereby the learning and memory circuitry.

Overlay of Epi, stress and learning and memory circuitry. This consolidated figure of the adrenergic, stress and learning and memory circuitry demonstrates the extensive overlap of these circuitries. Catecholamines do not, in general, cross the blood–brain barrier. However, Epi activation of vagal β-adrenergic receptors permits Epi to communicate with the CNS and thereby central components of the stress axis as well as learning and memory centers
This potential overlap in the adrenergic circuitry with the learning/memory circuitry may provide the opportunity to track stress recollection or delayed stress memory extinction. If stress is prolonged or repeated, we hypothesize that it may evoke long-term changes in Epi expression and the mechanisms by which Epi may be regulated by stress. Selective agonists or inhibitors of β-adrenergic receptors, e.g., isoprenaline and Epi or sotalol, alone or in combination, inhibitors of PNMT, which cross the BBB, e.g., 2,3-dichloro-α-methylbenzylamine (DCMB) and glucocorticoid antagonists, e.g., RU38486, could be used to part and parcel out if and how changes in Epi elicited by stress, adrenergic receptor function or changes in glucocorticoid responses may alter stress memory and responses to manifest in long-term illness.
Stress-Induced Changes in Epi and Potential for Stress-Induced Illness
Information from the preceding sections demonstrate the following: (1) Epi is a short-term neuromodulator of behavioral and physiological responses activated by stress to counter the stress and restore homeostasis, (2) Epi itself is genetically regulated by stress but the underlying mechanisms by which this occurs remain largely unknown, (3) up-regulation of Epi, as occurs during stress, is associated with a variety of stress-induced illnesses of cardiovascular, immune, cancer-related or behavioral etiologies, and (4) Epi plays an important role in learning and memory consolidation. Why would one neurohormone subsume multiple varied and at times, conflicting roles?
We hypothesize that Epi can serve as dual adaptive and maladaptive modulator of the stress response, the “good” guy as well as the “bad guy.” Stress-induced, short-term, rapid up- and down-regulation of Epi may lend to adaptation while long-term, sustained elevation may precipitate maladaptation and illness. By understanding the molecular mechanisms by which Epi responses evoke adaptation or maladaptation, respectively, through stress resiliency or increased stress sensitivity, we may be able to provide alternative therapeutics to manage stress and hence, stress-associated illness.
As a first approach, we are examining conditions which engender stress resiliency versus facilitate stress sensitivity. The goal is to develop a model whereby a stressor is identified, which on repeated exposure, evokes long-term, enduring changes in Epi expression to alter physiological function(s), e.g., immune, cardiovascular, nervous system, in a detrimental way lending to illness. Adverse, sustained stress effects will be detected through delayed extinction of stress memory. We are currently using fear-potentiated startle (FPS), a behavioral measure of conditioned fear, to investigate the effect of chronic stress in rats (Davis and Astrachan 1978). Rats are permitted to acquire conditioned fear by exposure to a neutral cue (light) and aversive event (foot shock), followed by testing with a series of startle-eliciting noise bursts (NBs), in the presence or absence of light. Conditioned fear to the light is defined as the FPS magnitude, the percentage increase in startle magnitude in the presence versus the absence of the light (Walker and Davis 2002).
Conditioned fear can be reduced through extinction, a secondary learning process where the fear-conditioned rat is exposed to repeated light in the absence of foot shock (Myers and Davis 2007). The rat learns that light no longer predicts foot shock and its FPS response to the light declines in magnitude in successive test sessions. We are interested in the effect of repeated stress on FPS extinction because extinction is malleable and stress-sensitive (Izquierdo et al. 2006). In addition, extinction rate serves as a measure of the strength of fear conditioning memory as stronger memories resist extinction (Gwinn 1951). Details of our FPS paradigm are provided in Fig. 5 and references cited in the legend. As stressors, we have used IMMO stress or the milder, FS stress.

Stress resiliency. To define conditions of stress that may lead to stress resiliency, rather than stress sensitivity, fear-potentiated-startle was used as a read-out of stress memory (Carlezon Jr et al. 2005; Meloni et al. 2006; Myers and Davis 2004) for immobilization stress and forced swim stress. Male Sprague–Dawley rats were tested for startle to cull outliers and separated into control and experimental groups (n = 8/group). a Protocol for FPS training/testing, repeated stress and FPS extinction/testing. b Behavioral data for IMMO (2 h daily for 7 days). c Behavioral data for FS (5 min daily for 7 days)
Our hypothesis was that Epi elevation, induced by repeated stress, if of greater and/or sustained magnitude, might delay extinction of the memory of the stress, given Epi’s role in memory consolidation. However, delayed extinction was not observed for either type of stress. In the case of IMMO stress, controls showed an apparent increase in the strength of fear conditioning memory between Days 3 and 11, whereas this effect was blunted in the stressed group. Rates of extinction in the two groups were similar. For FS stress, control and experimental rats showed similar extent of fear on Day 11 but diverged during the extinction phase, with the stressed group exhibiting more rapid extinction than the control group. Hence, contrary to our hypothesis that stress would delay extinction, the opposite emerged. One possible interpretation of this finding is that the previously stressed rats showed resiliency to subsequent stressors.
Curiously, the molecular markers, Egr-1, Sp1, HIF1α, and PNMT, showed the same elevated pattern of response described in our earlier IMMO studies, although the magnitude of response varied in two experiments of nearly identical design. In general, HIF1α mRNA was robustly stimulated and experimental animals showed similar induction of HIF1α protein, Egr-1 mRNA and protein but not as marked as previously. Sp1 mRNA was weakly stimulated at best while its protein was markedly elevated, and PNMT mRNA was highly elevated but only experimental animals showed the same in PNMT protein. In contrast to IMMO stress, for FS stress, molecular indices showed no consistent induction in stressed versus control rats with the exception of PNMT mRNA and protein, where both were significantly elevated. At present, we are repeating the FS stress experiment, except that the FS will be extended to 15 min to determine if duration of FS stress affects adrenergic molecular indices as with IMMO.
The studies to date clearly raise many questions, an important one being whether our behavioral read-out for stress, FPS, is a stressor itself. The key will be to identify the appropriate paradigms for examination of resiliency versus facilitation of stress sensitivity. Little is known about biological mechanisms associated with either. That knowledge will facilitate the design of strategies to understand how the stress hormone epinephrine contributes to both and provide avenues for alternative treatment strategies for stress-related illness.
References
Abraham J, Mudd JO, Kapur N, Klein K, Champion HC, Wittstein IS (2009) Stress cardiomyopathy after intravenous administration of catecholamines and beta-receptor agonists. J Am Coll Cardiol 53:1320–1325
Ameredes BT (2011) Beta-2-receptor regulation of immunomodulatory proteins in airway smooth muscle. Front Biosci (Schol Ed) 3:643–654
Cahill L, Alkire MT (2003) Epinephrine enhancement of human memory consolidation: interaction with arousal at encoding. Neurobiol Learn Mem 79:194–198
Cannon WG, De La Paz D (1911) Emotional stimulation of adrenal secretion. Am J Physiol 28:64–70
Carlezon WA Jr, Rohan ML, Mague SD, Meloni EG, Cayetano K, Tomasiewicz HC, Rouse ED, Cohen BM, Renshaw PF (2005) Antidepressant-like effects of cranial stimulation within a low energy magnetic field in rats. Biol Psychiatry 57:571–576
Davis M, Astrachan DI (1978) Conditioned fear and startle magnitude: effects of different footshock or backshock intensities used in training. J Exp Psychol Anim Behav Process 4:95–103
Debiec J, Bush DEA, LeDoux JE (2011) Noradrenergic enhancement of reconsolidation in the amygdala impairs extinction of conditioned fear in rats—a possible mechanism for the persistence of traumatic memories in PTSD. Depress Anxiety 28:186–193
Dikanović M, Demarin V, Kadojić D, Kadojić M, Trkanjec Z, Titlić M, Bitunjac M, Soldo-Butković S (2011) Effect of elevated catecholamine levels on cerebral hemodynamics in patients with chronic post-traumatic stress disorder. Coll Antropol 35:471–475
Dimsdale JE (2008) Psychological stress and cardiovascular disease. J Am Coll Cardiol 51:1237–1246
Donovan E (2010) Propranolol use in the prevention and treatment of posttraumatic stress disorder in military veterans: forgetting therapy revisited. Prespect Biol Med 53:61–74
Ebert SN, Wong DL (1995) Differential activation of the rat phenylethanolamine N-methyltransferase gene by Sp1 and Egr-1. J Biol Chem 270:17299–17305
Ebert SN, Balt SL, Hunter JPB, Gashler A, Sukhatme V, Wong DL (1994) Egr-1 activation of rat adrenal phenylethanolamine N-methyltransferase gene. J Biol Chem 269:20885–20898
Ebert SN, Ficklin MB, Her S, Siddall BJ, Bell RA, Morita K, Ganguly K, Wong DL (1998) Glucocorticoid-dependent action of neural crest factor AP-2: stimulation of phenylethanolamine N-methyltransferase gene expression. J Neurochem 70:2286–2295
Esler M (2010) The 2009 Carl Ludwig Lecture: pathophysiology of the human sympathetic nervous system in cardiovascular diseases: the transition from mechanisms to medical management. J Appl Physiol 108:227–237
Fineschi V, Michalodimitrakis M, D’Errico S, Neri M, Pomara C, Piezzo I, Turillazzi E (2010) Insight into stress-induced cardiomyopathy and sudden cardiac death due to stress. A forensic cardio-pathologist point of view. Forensic Sci Int 194:1–8
Fragala MS, Kraemer WJ, Mastro AM, Denegar CR, Volek JS, Hakkinen K, Anderson JM, Lee EC, Maresh CM (2011) Leukocyte b2-adrenergic receptor expression in response to resistance exercise. Med Sci Sports Exerc 43:1422–1432
Gaykema RP, Goehler LE (2011) Ascending caudal medullary catecholamine pathways drive sickness-induced deficits in exploratory behaviors: brain substrates for fatigue. Brain Behav Immun 25:443–460
Greene LA, Tischler AS (1976) Establishment of a noradrenergic clonal line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc Natl Acad Sci USA 73:2424–2428
Gwinn GT (1951) Resistance to extinction of learned fear-drives. J Exp Psychol 42:6–12
Heffner KL (2011) Neuroendocrine effects of stress on immunity in the elderly: implications for inflammatory disease. Immunol Allergy Clin North Am 31:95–108
Her S, Bell RA, Bloom AK, Siddall BJ, Wong DL (1999) Phenylethanolamine N-methyltransferase gene expression: Sp1 and MAZ potential for tissue specific expression. J Biol Chem 274:8698–8707
Her S, Claycomb R, Tai TC, Wong DL (2003) Regulation of the rat phenylethanolamine N-methyltransferase gene by transcription factors Sp1 and MAZ. Mol Pharmacol 64:1180–1188
Herault J, Perrot A, Barthelemy C, Buchler M, Cherpi C, Leboyer M, Sauvage D, Lelord G, Mallet J, Muh J (1993) Possible association of c-Harvey-Ras-1 (HRAS-1) marker with autism. Psychiatry Res 46:261–267
Higgins JP, Tuttle T, Higgins JA (2010) Altitude and the heart: is going high safe for your cardiac patient. Am Heart J 159:25–32
Hoffman BB (2001) Catecholamines, sympathomimetic drugs, and adrenergic receptor antagonists. In: Hardman JG, Limbird LE (eds) Goodman & Gilman’s the pharmacological basis of therapeutics, vol 10. McGraw Hill, New York, pp 215–268
Hokfelt T, Fuxe K, Goldstein M, Johansson O (1974) Immunohistochemical evidence for the existence of adrenaline neurons in the rat brain. Brain Res 66:235–251
Hokfelt T, Johansson O, Goldstein M (1984) Central catecholamine neurons as revealed by immunochemistry with special reference to the adrenaline neurons. In: Bjorklund A, Hokfelt T (eds) Handbook of neuroanatomy, vol 2. Elsevier, Amsterdam, pp 157–276
Hopfl G, Ogunshola O, Gassmann M (2004) HIFs and tumors—causes and consequences. Am J Physiol Regul Integr Comp Physiol 286:R608–R623
Inbar S, Neeman E, Avraham R, Benish M, Rosenne E, Ben-Eliyahu S (2011) Do stress responses promote leukemia progression? An animal study suggesting a role for epinephrine and prostaglandin-E2 through reduced NK activity. PLoS One 6:1–11
Izquierdo A, Wellman CL, Holmes A (2006) Brief uncontrollable stress causes dendritic retraction in infralimbic cortex and resistance to fear extinction in mice. J Neurosci 26:5733–5738
Ji Y, Snyder EM, Fridley BL, Salavaggione OE, Moon I et al (2008) Human phenylethanolamine N-methyltransferase genetic polymorphisms and exercise-induced epinephrine release. Physiol Genomics 33:323–332
Khouri S, Imran N (2009) Stress cardiomyopathy (Takotsubo cardiomyopathy). Clin Med Cardiol 3:93–99
Kostrzewa RM (2007) The blood–brain barrier for catecholamines—revisited. Neurotoxic Res 11:261–272
Kvetnansky R, Nankova B, Rusnak M, Micutkova L, Kubovcakova L, Dronjak SK, Krizanova O, Sabban EL (2002) Differential gene expression of tyrosine hydroxylase in rats exposed long-term to various stressors. In: Nagatsu T, Nabeshima T, McCarty R, Goldstein DS (eds) Catecholamine research: from molecular insights to clinical medicine vol. Plenum, New York, pp 317–320
Kvetnansky R, Sabban EL, Palkovits M (2009) Catecholaminergic systems in stress: structural and molecular genetic approaches. Physiol Rev 89:535–606
Laukova M, Vargovic P, Krizanova O, Kvetnansky R (2010) Repeated stress down-regulates β2- and α2C-adrenergic receptors and up-regulates gene expression of IL-6 in the rat spleen. Cell Mol Neurobiol 30:1077–1087
Launay JM, Bursztein C, Ferrari P (1987) Catecholamine metabolism in infantile autism: a controlled study of 22 autistic children. J Autism Dev Disord 17:333–347
Mausbach BT, Roepke SK, Ziegler MG, Milic M, von Kanel R, Dimsdale JE, Mills PJ, Patterson TL, Allison MA, Ancoli-Israel S, Grant I (2010) Association between chronic caregiving stress and impaired endothelial function in the elderly. J Am Coll Cardiol 55:2599–2606
McEwen BS (2007) Physiology and neurobiology of stress. Physiol Rev 87:873–904
McEwen BS, Gianaros PT (2010) Central role of the brain in stress and adaptation: links to socioeconomic status, health, and disease. Ann NY Acad Sci 1186:190–222
McGaugh JL (2004) The amygdala modulates the consolidation of memories of emotionally arousing experiences. Annu Rev Neurosci 27:1–28
Meloni EG, Gerety LP, Knoll AT, Cohen BM, Carlezon WA Jr (2006) Behavioral and anatomical interactions between dopamine and corticotropin-releasing factor in the rat. J Neurosci 26:3855–3863
Mravec B (2011) Role of catecholamine-induced activation of vagal afferent pathways in regulation of sympathoadrenal system activity: negative feedback loop of stress response. Endocr Regul 45:37–41
Myers KM, Davis M (2004) AX+, BX- discrimination learning in the fear-potentiated startle paradigm: possible relevance to inhibitor fear learning in extinction. Learn Mem 11:464–475
Myers KM, Davis M (2007) Mechanisms of fear extinction. Mol Psychiatry 12:120–150
Peng Y-J, Nanduri J, Khan SA, Yuan G, Wang N, Kinsman B, Vaddi DR, Kumar GK, Garcia JA, Semenza GL, Prabhakar NR (2011) Hypoxia-inducible factor 2a (HIF-2a) heterozygous-null mice exhibit exaggerated carotid body sensitivity to hypoxia, breathing instability, and hypertension. Proc Natl Acad Sci USA 108:3065–3070
Roozendaal B, McEwen BS, Chattarji S (2009) Stress, memory and the amygdala. Nat Rev Neurosci 10:423–433
Ross ME, Evinger MJ, Hyman SE, Carroll JM, Mucke L, Comb M, Reis DJ, Joh TH, Goodman HM (1990) Identification of a functional glucocorticoid response element in the phenylethanolamine N-methyltransferase promoter using fusion genes introduced into chromaffin cells in primary culture. J Neurosci 10:520–530
Santra M, Santara S, Zhang J, Chopp M (2008) Ectopic decorin expression up-regulates VEGF expression in mouse cerebral endothelial cells via activation of the transcription factors Sp1, HIF1α, and Stat3. J Neurochem 105:324–337
Schreurs J, Seelig T, Schulman H (1986) Beta 2-adrenergic receptors on peripheral nerves. J Neurochem 46:294–296
Sood AK, Armaiz-Pena GN, Halder J, Nick AM, Stone RL, Hu W, Carroll AR, Spannuth WA, Deavers MT, Allen JK, Han LY, Kamat AA, Shahzad MMK, McIntyre BW, Diaz-Montero CM, Jennings NB, Lin YG, Merritt WM, DeGeest K, Vivas-Mejia PE, Lopez-Berestein G, Schaller MD, Cole SW, Lutgendorf SK (2010) Adrenergic modulation of focal adhesion kinase protects human ovarian cancer cells from anoikis. J Clin Invest 120:1515–1523
Stone EA, Lin Y, Rosengarten H, Kramer HK, Quartermain D (2003) Emerging evidence for a central epinephrine-innervated α1-adrenergic system that regulates behavioral activation and is impaired in depression. Neuropsychopharmacology 28:1387–1399
Tai TC, Wong DL (2003) Protein kinase A and protein kinase C signaling pathway interaction in phenylethanolamine N-methytransferase gene regulation. J Neurochem 85:816–829
Tai TC, Claycomb R, Her S, Bloom AK, Wong DL (2002) Glucocorticoid responsiveness of the rat phenylethanolamine N-methyltransferase gene. Mol Pharmacol 61:1385–1392
Tai TC, Claycomb R, Siddall BJ, Bell RA, Kvetnansky R, Wong DL (2007) Stress-induced changes in epinephrine expression in the adrenal medulla in vivo. J Neurochem 101:1108–1118
Tai TC, Wong-Faull DC, Claycomb R, Wong DL (2009) Hypoxic stress-induced changes in adrenergic function: role of HIF1α. J Neurochem 109:513–524
Tai TC, Wong-Faull DC, Claycomb R, Wong DL (2010) Hypoxia and adrenergic function: molecular mechanisms related to Egr-1 and Sp1 activation. Brain Res 1353:14–27
Tillinger A, Bruderova V, Kubovcakova L, Zeman M, Kopacek J, Novakova M, Kvetnansky R, Krizanova O (2006) Gene expression of the phenylethanolamine N-methyltransferase is differently modulatedin cardiac atria and ventricles. Gen Physiol Biophys 25:355–364
Ulrich-Lai YM, Herman JP (2009) Neural regulation of endocrine and autonomic stress responses. Nat Rev Neurosci 10:397–409
Viskupic E, Kvetnansky R, Sabban EL, Fukuhara K, Weise VK, Kopin IJ, Schwartz JP (1994) Increase in rat adrenal phenylethanolamine N-methyltransferase mRNA level caused by immobilization stress depends on intact pituitary-adrenocortical axis. J Neurochem 63:808–814
Walker DL, Davis M (2002) Quantifying fear potentiated startle using absolute versus proportional increase scoring methods: implications for the neurocircuitry of fear and anxiety. Psychopharmacology 164:318–328
Wong DL, Tank AW (2007) Stress-induced catecholaminergic function: transcriptional and post-transcriptional control. Stress 10:121–130
Wong DL, Lesage A, Siddall B, Funder JW (1992) Glucocorticoid regulation of phenylethanolamine N-methyltransferase in vivo. FASEB J 6:3310–3315
Wong DL, Siddall BJ, Ebert SN, Bell RA, Her S (1998) Phenylethanolamine N-methyltransferase gene expression: synergistic activation by Egr-1, AP-2 and the glucocorticoid receptor. Mol Brain Res 61:154–161
Wong DL, Her S, Tai TC, Bell RA, Rusnak M, Farkas R, Kvetnansky R, Shih J (2002) Stress-induced expression of phenylethanolamine N-methyltransferase: normal and knock out animals. In: McCarty R, Aguilera G, Sabban EL, Kvetnansky R (eds) Stress: neural, endocrine and molecular studies. Taylor and Francis, London, pp 129–135
Wong DL, Tai TC, Wong-Faull DC, Claycomb R, Kvetnansky R (2004) Genetic mechanisms for adrenergic control during stress. In: Pacak K, Aguilera G, Sabban EL, Kvetnansky R (eds) Stress: current neuroendocrine and genetic approaches, vol 1018. Ann. N.Y. Acad. Sci, New York, pp 387–397
Wong DL, Tai TC, Wong-Faull DC, Claycomb R, Kvetnansky R (2008) Adrenergic responses to stress: transcriptional and post-transcriptional changes. Ann NY Acad Sci 1148:249–256
Wong DL, Tai TC, Wong-Faull DC, Claycomb R, Siddall BJ, Bell RA, Kvetnansky R (2010) Stress and adrenergic function: HIF1α, a potential regulatory switch. Cell Mol Neurobiol 30:1451–1457
Young EA, Breslau N (2004) Cortisol and catecholamines in posttraumatic stress disorder. Arch Gen Psychiatry 61:394–401
Zeb M, Sambu N, Scott P, Curzen N (2011) Takotsubo cardiomyopathy: a diagnostic challenge. Postgrad Med J 87:51–59
Zhang W, Shibamoto T, Kuda Y, Ohmukai C, Kurata Y (2011) Pulmonary vasoconstrictive and bronchoconstrictive responses to anaphylaxis are weakened via b2-adrenoceptor activation by endogenous epinephrine in anesthetized rats. Anesthesiology 114:614–623
Acknowledgments
This study was supported by The Spunk Fund, Inc., the Sobel-Keller Research Fund, McLean Hospital, and the Emerald Foundation, Inc. (DLW), NSERC and CIHR grants (TCT), MH063266 (WAC) and Slovak Grants, APVV-0088-10 and VEGA 2/0036-11 and 2/0188-09 (RK).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wong, D.L., Tai, T.C., Wong-Faull, D.C. et al. Epinephrine: A Short- and Long-Term Regulator of Stress and Development of Illness. Cell Mol Neurobiol 32, 737–748 (2012). https://doi.org/10.1007/s10571-011-9768-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10571-011-9768-0