
Abstract
The Slit-Robo GTPase-activating proteins (srGAPs) are important multifunctional adaptor proteins involved in various aspects of neuronal development, including axon guidance, neuronal migration, neurite outgrowth, dendritic morphology and synaptic plasticity. Among them, srGAP3, also named MEGAP (Mental disorder-associated GTPase-activating protein), plays a putative role in severe mental retardation. SrGAP3 expression in ventricular zones of neurogenesis indicates its involvement in early stage of neuronal development and differentiation. Here, we show that overexpression of srGAP3 inhibits VPA (valproic acid)-induced neurite initiation and neuronal differentiation in Neuro2A neuroblastoma cells, whereas knockdown of srGAP3 facilitates the neuronal differentiation in this cell line. In contrast to the wild type, overexpression of srGAP3 harboring an artificially mutation R542A within the functionally important RhoGAP domain does not exert a visible inhibitory effect on neuronal differentiation. The endogenous srGAP3 selectively binds to activated form of Rac1 in a RhoGAP pull-down assay. We also show that constitutively active (CA) Rac1 can rescue the effect of srGAP3 on attenuating neuronal differentiation. Furthermore, change in expression and localization of endogenous srGAP3 is observed in neuronal differentiated Neuro2A cells. Together, our data suggest that srGAP3 could regulate neuronal differentiation in a Rac1-dependent manner.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Slit-Robo GTPase-activating proteins (srGAPs) were first identified as a novel family of RhoGAP that transduce Slit-Robo activation to the intracellular signaling cascades (Wong et al. 2001). In mammals, the srGAP family consists of four members, the highly related srGAP1, srGAP2, and srGAP3, and the more distantly related ARHGAP4/RhoGAP C1 (Aspenstrom 2009). The srGAPs are multi-domain proteins: they all possess an N-terminal Fer/Fes CIP4 homology (FCH) domain, a RhoGAP domain, a Src homology 3 (SH3) domain, and two predicted coiled-coil motifs (Aspenstrom 2009). Structurally, the FCH domain and the adjacent coiled-coil motif are combined and renamed as the FCH-Bin/Amphiphysin/Rvs (F-BAR) domain (Itoh et al. 2005; Tsujita et al. 2006). All the three domains F-BAR, SH3, RhoGAP and coiled-coil motifs are able to mediate protein–protein interaction and signal transduction (Aspenstrom 2009).
The srGAP1, 2, and 3 have recently been reported to be expressed in multiple central nervous system tissues throughout embryonic, early postnatal, and adult brain development (Wong et al. 2001; Waltereit et al. 2008; Yao et al. 2008; Bacon et al. 2009; Guerrier et al. 2009). SrGAP1 directly binds to Robo1 intracelluar region and specifically down-regulates Cdc42 activation in migrating neurons (Wong et al. 2001). SrGAP2 induces filopodia formation through its F-BAR domain and affects neurite branching and neuronal migration (Guerrier et al. 2009). SrGAP3, which is also called MEGAP or WRP, has been shown to regulate Rac1 and Cdc42 and be able to mediate Rac-dependent neurite outgrowth (Soderling et al. 2002). Furthermore, SrGAP3 has been demonstrated to bind to the Arp2/3 activating protein WAVE-1, and the WAVE1-srGAP3 complex is involved in the regulation of dendritic morphology and synaptic plasticity (Soderling et al. 2002, 2007). Interestingly, it is reported that disruption of srGAP3 protein contributes to severe mental retardation in human (Endris et al. 2002). ARHGAP4 is highly expressed in hematopoietic cells and also expressed in the brain. ARHGAP4, which regulates Cdc42, Rac1, and RhoA, is involved in the regulation of cellular and axonal motility (Tribioli et al. 1996; Vogt et al. 2007).
It is notable that the expression pattern of srGAPs is well matched with their roles in the process of neuronal development. SrGAP2 expression in zones of neuronal differentiation and srGAP3 in ventricular zones of neurogenesis indicates that srGAPs are involved in the early stages of neuronal development (Wong et al. 2001; Bacon et al. 2009; Guerrier et al. 2009). Moreover, srGAP3 was also identified as a downstream effector of Neurogenin2, which has been demonstrated to participate in the specification of neural fates (Mattar et al. 2004). However, the exact roles of srGAPs in neuronal differentiation are still poorly understood.
In the present study, we are investigating the roles of srGAP3 during neuronal differentiation, using VPA-induced mouse Neuro2A neuroblastoma cells as a model. Neuro 2A is a mouse neural crest-derived cell line that has been extensively used to study neuronal differentiation and neurite growth. VPA (valproic acid), an inhibitor of histone deacetylase, is showed to induce neuronal differentiation of adult hippocampal neural progenitors (Hsieh et al. 2004). VPA can also induce neuronal differentiation of Neuro2A cells, and this model is used extensively as a cell culture model of neuronal differentiation (Regan 1985; Ito et al. 1997). We demonstrate that overexpression of srGAP3 inhibits VPA-induced neuronal differentiation of Neuro2A cells. Employment of RNA interference against srGAP3 results in an increase of differentiation rate. Furthermore, we also show that endogenous srGAP3 preferentially binds to activated form of Rac1 in a RhoGAP pull-down assay and the effect of srGAP3 on neuronal differentiation is through Rac1 signaling. In addition, endogenous srGAP3 expression and localization are changed during Neuro2A cell neuronal differentiation. These findings indicate that srGAP3 could play important roles in neuronal differentiation by transducing the signaling through Slit-Robo to Rac1.
Materials and Methods
DNA Constructs
The full-length of human srGAP3 cDNA sequence was constructed by the ligation of the PCR fragments encoding amino acids 1–425 to the KIAA0411 cDNA (gift from the Kazusa DNA Research Institute, Chiba, Japan). The other two human cDNA were also used, srGAP1 (IMAGE: 4153945, BC029919, PTG lab) and srGAP2 (gift from the Kazusa DNA Research Institute, Chiba, Japan). The full-length of srGAP1, srGAP2, and srGAP3 were amplified by LA-PCR (TAKARA). The primers used are listed below (5′–3′) and the restriction sites are underlined:
srGAP1 F: TCAAGCTTATGTCCACCCCGAGCCGATTC (HindIII site underlined);
srGAP1 R: GTGGATCCTTACATTGTGCATGACTTGTC (BamHI site underlined);
srGAP2 F: TCGAATTCACATGACGTCTCCAGCCAAATTC (EcoRI site underlined);
srGAP2 R: CAGTCGACTCAGACAGTACAAGACTTGTC (SalI site underlined);
srGAP3 F: TCAAGCTTATGTCATCTCAAACTAAGTTC (HindIII site underlined);
srGAP3 R: GTAGATCTCACATGGTGCCCGACTTGTCC (BglII site underlined).
The PCR products were ligated into the pcDNA3.1(+) vector (Invitrogen) to yield un-tagged expression plasmids, designated pcDNA-3.1(+)-srGAP1, pcDNA-3.1(+)-srGAP2, pcDNA-3.1(+)-srGAP3, or into the pEGFP-C3 vector (Clontech) to yield GFP-tagged expression plasmids, designated pEGFP-C3-srGAP1, pEGFP-C3-srGAP2, and pEGFP-C3-srGAP3.
The cDNA that encoded an arginine to alanine substitution at amino 542 (R542A) in the full-length srGAP3 protein was generated using Stratagene QuikChange Multi Site-Directed Mutagenesis Kit (Stratagene) according to the manufacturer’s protocol. The following is the primer pair used (5′–3′, base pair changes shown in bold).
F: CTCCAGCAGCAGGGGATCTTCGCAGTGCCAGGATCTCAGG;
R: CCTGAGATCCTGGCACTGCGAAGATCCCCTGCTGCTGGAG.
Affinity Purification of srGAP3 Antibodies
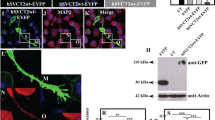
Two novel polyclonal antibodies against srGAP3, 3A1, and 3A3 were raised by immunization of rabbits with srGAP3 peptides. Two peptides, GDTHSPPRGLGPS (amino acids 870–882 of full-length srGAP3) and FPNSSADKSGTM (amino acids 1088–1099) were chosen from human srGAP3 sequence according to the hydrophobicity profile and antigenicity prediction analysis (Fig. 1a). The epitope peptide for 3A1 is conserved between human, rat, and mouse srGAP3, and for 3A3 there is only one mismatch. The antibodies were generated from rabbits immunized with keyhole limpet hemocyanin conjugated synthetic peptides. They were purified through protein-A agarose affinity columns, followed by peptide affinity purification.

The two srGAP3 polyclonal antibodies 3A1 and 3A3 specifically detect full-length srGAP3. a Schematic representation of the structure of full-length srGAP3 protein, which contains an FCH domain, a RhoGAP domain, a SH3 domain and two coiled-coil motifs. The FCH domain and the first coiled-coil motif assemble the F-BAR domain. The arrow indicates the arginine at the 542 residue. The arrowheads indicate the target regions of two srGAP3-specific polyclonal antibodies (3A1 and 3A3). b HEK293FT cells are transfected with mock vector or GFP-tagged full-length srGAP1, srGAP2, srGAP3. The lysates are detected by western blotting with two srGAP3 antibodies (3A1 and 3A3) and GFP antibody
RNA Interference
According to the targeting sequences of srGAP3, a pair of oligonucleotide encoding shRNA was designed using BLOCK-iT™ RNAi Designer. The oligonucleotides were annealed and cloned into the pcDNA6.2TM-GW/EmGFP vector (Invitrogen) to generate J33 shRNA expressing plasmid according to the instruction of the BLOCK-iTTM Lentiviral Pol II miR RNAi Expression Vector Kits (Invitrogen). The sequence of J33 shRNA are shown as below:
5′-TGCTGTGCAGTACCAGATACCAACAGGTTTTGGCCACTGACTGACCTGTTGGTCTGGTACTGCA-3’ (Sense); 5′-CCTGTGCAGTACCAGACCAACAGGTCA GTCAGTGGCCAAAACCTGTTG GTATCTGGTACTGCAC-3′ (Anti-sense).
Cell Culture, Transfection, and Differentiation Assay
HEK293FT cells (Invitrogen) were maintained in DMEM supplemented with 10% FBS, 100 U/ml penicillin, and 100 μg/ml streptomycin (Invitrogen). The mouse neuroblastoma cells, Neuro2A cells, were obtained from Institute of Biochemistry and Cell Biology, SIBS, CAS (Shanghai, China). They were cultured in DMEM containing 10% FBS, 1% non-essential amino acid (Invitrogen), 100 U/ml penicillin, and 100 μg/ml streptomycin. HEK293FT cells or Neuro2A cells were transfected using FuGENE HD Transfection Reagent (Roche) according to the manufacturer’s instructions. Transient transfection conditions were optimized for maximum expression and minimal toxicity. To induce neuronal differentiation, Neuro2A cells (at about 20% confluence) were transferred to serum-free optiMEM (Invitrogen) containing 1 mM VPA (Sigma) and allowed to extend neurites (Ito et al. 1997).
Statistical Analysis
For assessing differentiation, Neuro2A cells grown in 35-mm dishes were treated with optiMEM containing 1 mM VPA for 0, 24, or 48 h. Cells with neurite processes longer than two cell bodies were considered to be differentiated. Each group was analyzed by counting about 150–500 cells. Assessment of neurite outgrowth was performed by counting about 30 to 50 cells per condition. Neurite length and neurite number were quantified using Image Pro-Plus software. In each analysis, the data represent mean ± SEM of at least three experiments. For comparison, statistical significance was tested by one-way ANOVA or unpaired t test using Sigma Stat software.
Immunocytochemistry
Cells on Poly-l-Lysine-coated glass coverslips were fixed with 4% paraformaldehyde for 15 min at room temperature and then permeabilized by treatment with ice-cold methanol for 10 min. After being blocked by 20% normal donkey serum for 30 min, cells were incubated at 4°C overnight with primary 3A1 antibody (1:50) and GAP-43 antibody (Duan et al. 2003) (1:500) diluted in antibody buffer (50 mM Tris–HCl, pH 7.4, 150 mM NaCl, 100 mM l-Lysine, 1% BSA and 0.04% azide). After incubation with the primary antibodies, they were rinsed and incubated for 1 h with fluorescence-labeled secondary antibodies, following by treatment with Hoechst 33342 (10 μg/ml) for 10 min to stain the nuclei. After washing, the coverslips were mounted with PBS containing 50% glycerine.
Western Blotting
The western blotting assay had been previously described (Jin et al. 2003). In brief, whole-cell extracts were prepared in High KCl lysis buffer (10 mM Tris–HCl, pH 8.0, 140 mM NaCl, 300 mM KCl, 1 mM EDTA, 0.5% Triton X-100, and 0.5% sodium deoxycholate) with complete protease inhibitor cocktail (Roche). The protein concentration of the lysates was determined using BCA protein assay kit (Pierce, Rockford, USA). The lysates were then separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Roche). The membranes were then treated with 1% blocking solution in TBS, followed by incubation with primary antibodies and then POD-labeled secondary antibodies. The immunolabeled proteins were detected by BM Chemiluminescence Western Blotting kit (Roche). To assess the transfer of proteins, the membranes were stained with 0.1% naphthol blue black (Sigma) in 45% methanol and 10% acetic acid. Densitometry quantitation was acquired with Gel Doc 1000 system and analyzed using Quantity One software (BioRad). The antibodies we used were as following: 3A1, 1:4000; 3A3, 1:1000; β-actin antibody (AbMART, China), 1:1000; TuJ1 antibody (Sigma), 1:5000; GAP-43 antibody (Duan et al. 2003), 1:5000; GFP antibody (AbMART, China), 1:5000; GAPDH-HRP (KANGCHEN, China), 1:5000.
RhoGAP Pull-Down Assay
Affinity precipitation of RhoGAPs with activated mutants of RhoGTPase has been described previously (Garcia-Mata et al. 2006). Briefly, plasmids were transformed into DH5α cells and expression was induced by 100 μM isopropyl-β-d-thio-galactoside (TAKARA) overnight at room temperature. GST-fusion proteins were purified in batch on glutathione-agarose (Sigma). Neuro2A cells were lyzed in 20 mM HEPES (pH 7.5), 150 mM NaCl, 5 mM MgCl2, 1% TritonX-100, and 1 mM dithiothreitol with protease inhibitors. About 3,000 μg of protein lysates were incubated with 60 μg of purified RhoGTPase (CA) bound to glutathione-agarose for 2 h at 4°C, followed by washing three times with lysis buffer. Precipitated proteins were then solubilized in sample buffer and analyzed by western blotting.
RT-PCR Assay
To obtain total RNA, Neuro2A cells were extracted using Trizol Reagent (Invitrogen), following by treatment with RNase-free Dnase I (Roche). Reverse transcription reactions of total RNA and subsequent cDNA PCR were performed using BcaBestTM RNA PCR Kit (TAKARA) according to the manufacturer’s instruction. Gene-specific primer pairs for PCR are listed in Table 1. The PCR products were then subjected to 1% agarose gel electrophoresis and stained by ethidium bromide. The images were acquired in the Gel Doc 1000 system (BioRad) and analyzed using Quantity One software (BioRad).
Results
Characterization of Specific srGAP3 Antibodies
Our previous srGAP3 antibody is successfully applied for immunofluorescence staining experiments in tissue sections and primary culture cortical neurons (Yao et al. 2008). Two newly raised antigen-affinity purified polyclonal antibodies against srGAP3, 3A1, and 3A3 were used in our western blotting experiments. The epitope peptides of 3A1 and 3A3 antibodies are close to the C-terminal of human srGAP3 and do not locate in the F-BAR, SH3, RhoGAP domains, or coiled-coil motifs (Fig. 1a).
The specificity of these two srGAP3 polyclonal antibodies was evaluated by western blots. HEK293FT cells were transiently transfected with GFP-tagged srGAP1, srGAP2, and srGAP3. Western blotting of the cell lysates using GFP antibody demonstrated that all three transfected proteins were expressed (Fig. 1b, right panel). The same blots were then probed with 3A1 and 3A3 antibodies. Both antibodies specifically recognized the 170 kDa GFP-srGAP3 (150 kDa srGAP3 plus 20 kDa GFP), but not GFP-srGAP1 or GFP-srGAP2 (Fig. 1b, left panel and middle panel), which indicated that the two antibodies are able to specially blot srGAP3 and not shown to display cross reactivity with other srGAPs. In our experiments, these antibodies also recognize mouse and rat srGAP3 (data not shown).
In Neuro2A cells, the affinity purified antibody 3A1 or 3A3 also recognize endogenous srGAP3 protein as shown by immnuofluoresence staining: in differentiated cells, the endogenous srGAP3 was accumulated in the nucleus and present on small protrusion and neurites (see below). Given the complexity of the staining pattern, it is essential to exclude antibody artifacts. Three lines of evidence validate the specificity of the staining. First, the anti-srGAP3 antibody recognized single bands corresponding to the predicted protein size. Second, the membrane, neurite and nuclear staining pattern were consistent with those of a commercial antibody (WRP: S-16, Santa Cruz) and our previous srGAP3 antibody (Yao et al. 2008), and the immunofluorescence staining was abolished upon preincubation of the antibodies with the respective antigen peptide (data not shown). Third, knockdown of srGAP3 by shRNA drastically reduced both immunofluorescence and Western blot signals (see below).
SrGAP3 Inhibits Neuronal Differentiation of VPA-Induced Neuro2A Cells
Neuro2A cells were transiently co-transfected with an EGFP expression vector and a srGAP3 expression vector, pcDNA3.1(+)-srGAP3. The transfection efficiency was around 70–80% and the transfected cells were labeled by GFP fluorescence (Fig. 2a). Western blots have demonstrated that srGAP3 level was significantly increased compared to mock transfection (Fig. 2b). Similar bands could be observed from endogenous mouse srGAP3 and overexpressed human srGAP3 (Fig. 2b). In the blot, srGAP3 antibody detected additional bands at 130 and 95 kDa from extracts of Neuro2A cells, whereas bands at 100 and 70 kDa were detected from lysis of GFP-srGAP3 expressed HEK293FT cells (Figs. 1b, 2b).

SrGAP3 inhibits VPA-induced neuronal differentiation. a The transfected cells are recognized by EGFP expression. Cells with processes longer than two cell bodies are considered those differentiated (marked by arrow). Scale bar = 50 μm. b Western blotting analysis Neuro2A cells lysates by anti-srGAP3 (3A3) and anti-β-actin antibodies. Neuro2A cells are co-transfected along with empty vector plus the EGFP expression plasmid (lane 1) or the pcDNA3.1(+)-srGAP3 expression plasmid plus the EGFP expression plasmid (lane 2). The expression of β-actin is used as a loading control. Left panel low exposure; right panel: high exposure. c Quantification of the differentiation rates from Neuro2A cells in the absent of VPA, or under VPA stimulation for 24 or 48 h. Values are means ± SEM, ** P < 0.01. d Assessment of neurite outgrowth of Neuro2A cells by VPA stimulation for 24 h. Three endpoints are quantified: total neurite length (upper panel), longest neurite length (middle panel), and neurite number per cell (lower panel). Values are means ± SEM
The effects of srGAP3 on Neuro2A cells differentiation were analyzed. Cells with neurite processes longer than two cell bodies were counted as differentiated cells. In the absent of VPA stimulation, overexpression of srGAP3 did not affect the differentiation rate (mock transfection: 3.66 ± 0.45%; srGAP3 overexpression: 4.66 ± 0.98%) (Fig. 2c). However, overexpression of srGAP3 in the Neuro2A cells had an approximately 40% inhibitory effect on neuronal differentiation by VPA stimulation for 24 h (Fig. 2c). The differentiation rate was reduced to 14.17 ± 1.38% compared to mock transfection, which was 22.13 ± 1.36%. When the Neuro2A cells were induced for 48 h, overexpression of srGAP3 also resulted in a remarkable reduce in differentiation rates, from 42.76 ± 3.02% to 28.67 ± 2.59% (Fig. 2c). Results remained consistent and statistically significant even if cells with at least one neurite process longer than one half of the cell body diameter were defined as differentiated cells, which indicated that our grouping criteria did not introduce bias into the analysis (data not shown).
We next tested whether srGAP3 could affect neurite outgrowth of differentiated Neuro2A cells that were exposed to VPA for 24 h. The results showed that overexpression of srGAP3 did not affect total neurite length (srGAP3, 148.718 ± 7.616 μm; mock, 146.001 ± 5.185 μm) (Fig. 2d) or the longest neurite length (srGAP3, 92.043 ± 6.710 μm; mock, 88.910 ± 3.810 μm) (Fig. 2d). Similarly, the average neurite number did not show significant difference (srGAP3, 2.642 ± 0.076; mock, 2.626 ± 0.159) (Fig. 2d). Our results showed that srGAP3 did not alter neurite outgrowth of differentiated Neuro2A cells.
Knockdown of Endogenous srGAP3 Promotes Neuronal Differentiation of Neuro2A Cells
SrGAP3 mRNA is highly expressed in Neuro2A cells (Supplementary Fig. 1). As indicated in our western blotting, the endogenous expression of srGAP3 protein could also be detected (Fig. 2b, left lane under long exposure). To assess whether endogenously expressed srGAP3 is involved in the regulation of VPA-induced neuronal differentiation of Neuro2A cells, we examined the effects of srGAP3 depletion from Neuro2A cells by shRNA-based knockdown method. We constructed plasmids that express different small hairpin RNAs against srGAP3 and examined their knockdown efficiency on srGAP3 expression in HEK293FT cells. One of these shRNA, J33 shRNA, which was demonstrated to knockdown srGAP3 expression dramatically, was isolated (data not shown). Neuro2A cells were transfected with either negative control or J33 shRNA expressing vectors. The results of the western blotting proved that transfection of J33 shRNA caused a significant reduction in expression of endogenous srGAP3 (Fig. 3a). In addition, the immunostaining results also confirmed that endogenous srGAP3 level was knockdown by J33 shRNA transfection (Fig. 3b).

SrGAP3 knockdown increase the neuronal differentiation rate of Neuro2A cells. a Western blotting probe with anti-srGAP3 (3A3) and anti-β-actin antibodies from Neuro2A cells transfected with either control shRNA (lane 1) or J33 shRNA (lane 2). The expression of β-actin is used as loading control. b Neuro2A cells transfected with J33 shRNA are fixed and stained for srGAP3 and DNA. Note that in srGAP3 knockdown cells (labeled by GFP), the signals at the cell periphery and nuclear (indicated by arrowheads) can be mostly eliminated. Scale bar = 10 μm. c Quantification of the differentiation rates of control or srGAP3-knockdown Neuro2A cells. Values are means ± SEM, ** P < 0.01
Next, we asked whether transfection of J33 shRNA could affect neuronal differentiation. In the absence of VPA treatment, Neuro2A cell line transfected with J33 shRNA did not induce visible neuron-like differentiation (control shRNA: 4.18 ± 0.98%; J33 shRNA: 3.09 ± 0.55%) (Fig. 3c). After 24 h of VPA induction, the differentiation rate of srGAP3 knockdown cells was altered. Compared with the control shRNA transfected cells, J33 shRNA transfected cells exhibited a marked increase of neuronal differentiation upon VPA stimulation. The differentiation rate of the srGAP3 knockdown group was 29.29 ± 0.17% compared to control group which was 22.57 ± 0.93% (Fig. 3c). We also tested whether knockdown of srGAP3 could have similar effect when Neuro2A cells neuronal differentiation was induced by other treatment. Indeed, when Neuro2A cells were cultured in DMEM containing 2% FBS and 20 μM Retinoic Acid (Zeng and Zhou 2008), transfection of J33 shRNA also caused an increase in neuronal differentiation rates (control shRNA: 22.56 ± 1.25%; J33 shRNA: 27.47 ± 1.19%) (Supplementary Fig. 2). These data confirmed that endogenous srGAP3 is a negative regulator in neuronal differentiation of Neuro2A cells.
The Effect of srGAP3 on Neuronal Differentiation is Rac1-Dependent
SrGAP3 is a Rac1 and Cdc42 selective GTPase-activating protein (Endris et al. 2002; Soderling et al. 2002; Yang et al. 2006). We asked whether the role of srGAP3 in regulating neuronal differentiation of Neuro2A cells is related to the RhoGTPase signaling. A GST-pull-down assay by GST-fusion constitutively active (CA) Cdc42, Rac1, and RhoA was carried out with Neuro2A cell lysates. Our result demonstrated that endogenous srGAP3 selectively binds to activated form of Rac1, but not Cdc42 or RhoA (Fig. 4a). It suggested that Rac1 could serve as a downstream effector for srGAP3.

The effects of srGAP3 on Neuro2A differentiation are through the regulation of Rac1 activities. a Lysates of Neuro2A cells are incubated with control GST protein or GST-fusion CA-Cdc42, CA-Rac1, CA-RhoA in a pull-down assay. The precipitates are solubilized and processed to western blotting, using srGAP3 antibody (3A3). The amount of GST-fusion protein used in the assay is revealed by naphthol blue black staining. GST-fusion proteins are indicated by asterisks. b Quantification of the differentiation rates of Neuro2A cells expressing control vectors, srGAP3, srGAP3R542A, srGAP3 plus CA-Rac1, srGAP3 plus CA-Cdc42, srGAP3 plus CA-RhoA, CA-Rac1 and DN-Rac1, respectively. Values are means ± SEM, ** P < 0.01, *** P < 0.001
The activation of Rac1 GTPase, which has been shown to be important for the neuronal differentiation of N1E-115 neuroblasmtoma cells, is negatively regulated by RhoGAP (Kozma et al. 1997). To test the hypothesis that srGAP3 regulates neuronal differentiation through Rac1 signaling, we transfected constitutively active Rac1 Q61L (CA-Rac1) and dominant negative Rac1 T17N (DN-Rac1) (Yuan et al. 2003) into the Neuro2A cells, respectively. The CA-Rac1 transfected cells showed slightly increase in neuronal differentiation rate (28.37 ± 1.05%), whereas DN-Rac1 transfected cells had an approximately 40% inhibitory activity on neuronal differentiation (12.68 ± 1.56%), similar to the effect of srGAP3 overexpression (Fig. 4b). Since the RhoGAP domain of srGAP3 could be the direct inhibitor for Rac1, we then examined whether the RhoGAP domain of srGAP3 is required for its function. A mutant form of srGAP3 (srGAP3R542A), which lost the RhoGAP activity, was generated by replacing the conserved amino acid Arg542 with alanine in the RhoGAP domain (Fig. 1a). As expected, Neuro2A cells transfected with srGAP3R542A mutant did not show an inhibitory effect on VPA-induced differentiation. The differentiation rate in cells expressing srGAP3R542A was 22.64 ± 0.09%, compared to control level which was 21.53 ± 1.32%, while the differentiation rate of wild-type srGAP3 over-expressing cells reduced to 13.91 ± 1.22% (Fig. 4b). In addition, overexpression of the F-BAR domain of srGAP3 (deletion of the RhoGAP domain and Robo-interacting SH3 region) did not show any inhibitory effect on VPA-induced differentiation (data not shown).
We further test if activation of Rac1 could rescue the negative effect of srGAP3. Indeed, the differentiation rate was restored on co-transfection of srGAP3 with CA-Rac1. Compared with cells transfected with srGAP3 alone, co-expression of CA-Rac1 resulted in a significant increase in differentiation rate, from 13.91 ± 1.22% to 24.54 ± 1.15% (Fig. 4b), which is similar to the differentiation rate of control level. On the other hand, the active form of the other two RhoGTPase, Cdc42 Q61L (CA-Cdc42) or RhoA Q63L (CA-RhoA), failed to counteract the inhibitory effect of srGAP3 (srGAP3 + CA-Cdc42: 9.12 ± 1.06%; srGAP3 + CA-RhoA: 10.91 ± 0.75%) (Fig. 4b). Taken together, our observation indicated that srGAP3 functions through Rac1 signaling to regulate neuronal differentiation of Neuro2A cells.
Up-Regulation and Altered Localization of Endogenous srGAP3 During Differentiation of Neuro2A Cells
Since srGAP3 overexpression inhibits neuronal differentiation of Neuro2A cells, we next checked the expression level of endogenous srGAP3 in this cell line. Photomicrographs revealed that treatment with VPA-induced processes within 8 h, and additional incubation caused the formation of networks with long and bold neurites at 24 h. After 24 h of incubation with 1 mM VPA, around 20% of cells were exhibiting neurite growing (Fig. 5a).

The expression and subcellular localization of endogenous srGAP3 in Neuro2A cells are changed during VPA-induced neuronal differentiation. a Photomicrographs of morphological changes of Neuro2A cells, which are exposed to VPA for 0 or 24 h. The differentiation rates of different VPA stimulation time points are counted and showed in the histogram. b Neuro2A cells cultured in DMEM with 10% FBS (UD, upper panel) or opti-MEM plus 1 mM VPA (VPA, lower panel) are fixed and stained for srGAP3, GAP-43 and DNA. Nuclear-localized srGAP3 is detected both in un-differentiated (UD) and VPA-induced neuronal differentiated Neuro2A cells (indicated by arrowheads). SrGAP3 is also localized at the filopodia and lamellipodia structures before differentiation (indicated by arrows) and at the neurite structures and plasma membrane after Neuro2A differentiation (indicated by open arrowhead). Scale bar = 20 μm. c Lysates from Neuro2A cells exposed to VPA for the indicated times are subjected to western blot analysis. The expression level of srGAP3, together with two other neuronal markers, GAP-43 and TuJ1, is detected. The β-actin is used as a loading control and the relative expression ratio of srGAP3/β-actin, GAP-43/β-actin and TuJ1/β-actin are reported under each lane with 0 h expression level set to 1.0
The expression levels of srGAP3, together with two other neuronal markers, GAP-43 and TuJ1 (class III β-tubulin), were analyzed by western blotting. Upon VPA induction for 24 h, GAP-43 and TuJ1 were shown to increase mostly between 2.0- and 2.7-fold, respectively (Fig. 5c). As shown, the TuJ1 expression increased strikingly and reached a plateau in a short time, whereas the expression of GAP-43 increased gradually and relatively slower (Fig. 5c). During early differentiation phase, the expression of endogenous srGAP3 level in Neuro2A cells was up-regulated. The relative level has reached about 2-fold within 12 h and upon VPA induction for 48 h, the level was 2.7-fold compare to that of the un-differentiated cells (Fig. 5c). Interestingly, we observed a decline of srGAP3 expression in differentiated cells from 48 to 72 h (Fig. 5c). After exposed to VPA for 72 h, the expression level of srGAP3 was still 2-fold as high as control level (Fig. 5c).
Next, we examined the localization of endogenous srGAP3 protein during neuronal differentiation by immunofluorescence staining. Neuro2A cells were co-stained with 3A1 antibody and GAP-43 antibody. The results showed that in undifferentiated cells, srGAP3 was localized in the cytoplasm and the nucleus. In some fillopodia and lamellipodia structures at the cell periphery, the srGAP3 immunoreactive was also detected (Fig. 5b, upper panel). In differentiated cells which were exposed to 1 mM VPA for 24 h, the srGAP3 immunoreactivity was co-localized at the cytoplasma membrane with GAP-43 and was also accumulated in the neurite structures, while nuclear srGAP3 immunoreactive remained strong (Fig. 5b, lower panel). The highly dynamic pattern of endogenous srGAP3 localization in the process of neuronal differentiation suggests that membrane re-localization of srGAP3 might be functionally important.
Discussion
SrGAP3 has been implicated in the regulation of neuronal development and migration. However, its role in early stage of neuronal differentiation remains largely unexplored. In the present study, we have discovered that overexpression of srGAP3 reduced VPA-induced neuronal differentiation rate in Neuro2A cells. On the other hand, knockdown of endogenous srGAP3 by transfection with shRNA promoted neurite initiation and differentiation. SrGAP3 expression was also observed to be up-regulated during early phase of VPA-induced Neuro2A cells neuronal differentiation. These data imply that the mental retardation associated protein, srGAP3, may function as a negative regulator of neuronal differentiation.
In developing nervous system, srGAP3 is found to be expressed in ventricular zones in very early stages (Bacon et al. 2009). Similarly, in VPA-induced neuronal differentiated Neuro2A cells, endogenous srGAP3 expression level was up-regulated during early stages. Our hypothesis that srGAP3 could be involved in the mediation of neuronal differentiation is strongly strengthened by the expression pattern. Moreover, overexpression of srGAP3 in Neuro2A cells resulted in a reduction in differentiation rate, which supported that srGAP3 has an opposite effect on the VPA-induced neuronal differentiation. Our findings extend to indicate that srGAP3 may also be involved in the fate specification of adult neural progenitors or neural stem cells which could contribute to the higher brain functions, such as learning and emotional disabilities associated with schizophrenia, mental retardation, and autism spectrum disorders.
Besides srGAP3, the roles of other srGAP family members during neuronal differentiation are poorly understood. In Neuro2A cells, srGAP2, but not srGAP1, is expressed according to our RT-PCR results (Supplementary Fig. 1). We attempted to test if srGAP2 is involved in regulating Neuro2A cells differentiation, by transfecting srGAP2 in this cell line. However, overexpression of srGAP2 failed to show any visible effect on VPA-induced neuronal differentiation (data not shown). Meanwhile, in our GST-pull-down experiments, the endogenous srGAP2 protein from Neuro2A cells could not be precipitated by CA-Cdc42, CA-Rac1, or CA-RhoA. Indeed, this was in consistency with our previous study that the RhoGAP domain in srGAP2 has a relative weak hydrolysis activity toward GTP-Rac1 (Guerrier et al. 2009). We speculate that srGAP2 may not function in affecting VPA-induced neuronal differentiation of Neuro2A cells, at least not through regulating of RhoGTPase activities.
Although we observed an inhibitory effect of srGAP3 on neuronal differentiation, we did not notice a corresponding change in neurite extension in differentiated Neuro2A cells. These results are not in agreement with previous findings since transfection of WRP (srGAP3) is shown to reduce neurite length of cerebellar granular neurons (Soderling et al. 2002). This seemingly contradiction could be due to the different cell types and the difference of developmental stages in these studies. In our experiments, the Neuro2A cell line is used as an in vitro system. Neuro2A cells can differentiate into neurons upon serum starvation or treatment with reagents like retinoic acid or VPA (Schubert et al. 1969; Regan 1985; Wu et al. 1990). It is a reliable model to study the process of neuronal differentiation and neurite outgrowth. The upstream regulators of the srGAP family, Slit2 and Robo1, are both found to be highly expressed in Neuro2A cells (Supplementary Fig. 1), whereas Slit1, Slit2, or Robo1 is not expressed in the post-mitotic cerebellar granular neurons (Marillat et al. 2001; Hivert et al. 2002). The Slit-Robo signaling has been shown to be involved in the proliferation and morphological differentiation of cortical interneurons (Andrews et al. 2008). Thus, it is tempting to entertain the possibility that the effect of srGAP3 in Neuro2A cells is controlled by the upstream Slit-Robo signaling. Further studies are required to clarify this issue.
As a GTPase-activating protein, srGAP3 has been showed to act toward Rac1 and Cdc42 both in vitro and in vivo (Endris et al. 2002; Soderling et al. 2002; Yang et al. 2006). Numerous reports have documented the Rho family GTPases, typical RhoA, Rac1, and Cdc42 as important regulators involved in various aspects of neurite outgrowth during neuronal differentiation (Govek et al. 2005; Koh 2006). The activity of Rho GTPases is negatively regulated by Rho GTPases-activating proteins (RhoGAPs) (Brouns et al. 2001; Moon et al. 2003; Koh 2006). Our data demonstrated that the endogenous active srGAP3 in Neuro2A cells selectively binds to activated form of Rac1 in a RhoGAP pull-down assay. Interestingly, Rac1 is proved to induce membrane ruffling and lamellipodium formation (Ridley et al. 1992; Nobes and Hall 1995) and in neuroblastoma cells, activated Rac1 promotes neuronal differentiation and neurite extension (Kozma et al. 1997). In the Neuro2A cell line model, we observed that DN-Rac1 exhibited similar inhibitory effect with forced expressed srGAP3, and expression of a CA-Rac1 completely blocked the ability of srGAP3 to suppress neuronal differentiation. Importantly, the mutant form of srGAP3 (srGAP3R542A) failed to inhibit differentiation in this cell line. All data support the hypothesis that Rac1 could be a downstream effector for srGAP3 in the signaling cascade that mediates neurite initiation and neuronal differentiation.
We previously observed that srGAPs showed distinct nucleocytoplasmic localizations at different developmental stages (Yao et al. 2008). In differentiated Neuro2A cells, srGAP3 also shows nuclear localization. Indeed, the nuclear localization of srGAP3 is not unusual, since accumulating data have showed RhoGAPs and their downstream effectors like RhoGTPases are present in nucleus (Lanning et al. 2004; Yuan et al. 2007; Dingova et al. 2009). However, the detailed function of the nuclear-localized RhoGAP is largely unknown. One of the previous studies about nuclear-localized RhoGAP had caught our attention: a chromosome 13q12 encoded RhoGAP (ARHGAP1), which is predicted to have a chance of 65% to localize in the nucleus, is found to interact with one of the subunits of the SWI/SNF-like chromatin remodeling complex, SWI/SNF (SMARCD3/BAF60C) (Nagaraja and Kandpal 2004). Accumulated evidence has shown that the SWI/SNF-like chromatin remodeling complexes (BAF complexes) are playing important roles in post-mitotic neuronal development (Lessard et al. 2007; Wu et al. 2007; Yoo and Crabtree 2009), and the core subunit of the BAF complex, Brg1, is required for neuronal differentiation of P19 cells (Seo et al. 2005). Thus, it is possible that nuclear-localized Rho GAP, including srGAP3, may play important roles in nuclear signaling. At the mean time, we are actively investigating if nuclear-localized srGAP3 would participate in nuclear signaling in regulating neuronal differentiation.
Based on our observation of the Neuro2A differentiation studies, we conclude that srGAP3 negatively regulates the neurite initiation and neuronal differentiation. To our knowledge, it is the first time to report the direct involvement of srGAPs in neuronal differentiation. Further experiments are required to dissect the roles of srGAP3 in different compartments of the neuronal cell, for example, the nuclear-localized srGAP3 versus neurite-localized srGAP3. The role of srGAP3 at different phases of neuronal differentiation is another aspect of future studies.
Abbreviations
- srGAP:
-
Slit-Robo GTPase-activating protein
- VPA:
-
Valproic acid
- CA:
-
Constitutively active
- MEGAP:
-
Mental disorder-associated GAP
- GAP-43:
-
Growth associated protein-43
- FCH:
-
Fer/Fes CIP4 homology
- SH3:
-
Src homology 3
- F-BAR:
-
FCH-bin/amphiphysin/Rvs
- CC:
-
Coiled-coil
- WRP:
-
WAVE-associated Rac GAP
- GST:
-
Glutathione S-transferase
- GAPDH:
-
Glyceraldehyde 3-phosphate dehydrogenase
- Rac1:
-
Ras-related C3 botulinum toxin substrate 1
- RhoA:
-
Ras homolog gene family, member A
- Cdc42:
-
Cell division control protein 42 homolog
References
Andrews W, Barber M, Hernadez-Miranda LR, Xian J, Rakic S, Sundaresan V, Rabbitts TH, Pannell R, Rabbitts P, Thompson H, Erskine L, Murakami F, Parnavelas JG (2008) The role of Slit-Robo signaling in the generation, migration and morphological differentiation of cortical interneurons. Dev Biol 313:648–658
Aspenstrom P (2009) Roles of F-BAR/PCH proteins in the regulation of membrane dynamics and actin reorganization. Int Rev Cell Mol Biol 272:1–31
Bacon C, Endris V, Rappold G (2009) Dynamic expression of the Slit-Robo GTPase activating protein genes during development of the murine nervous system. J Comp Neurol 513:224–236
Brouns MR, Matheson SF, Settleman J (2001) p190 RhoGAP is the principal Src substrate in brain and regulates axon outgrowth, guidance and fasciculation. Nat Cell Biol 3:361–367
Dingova H, Fukalova J, Maninova M, Philimonenko VV, Hozak P (2009) Ultrastructural localization of actin and actin-binding proteins in the nucleus. Histochem Cell Biol 131:425–434
Duan XL, Huang WJ, Wang BR, Song JF, Jin WL, Guo X, Ju G (2003) Construction and expression of prokaryotic expression vector of GAP-43 and preparation of monoclonal antibody against GAP-43. Chin J Cell Mol Immunol 19:480–483
Endris V, Wogatzky B, Leimer U, Bartsch D, Zatyka M, Latif F, Maher ER, Tariverdian G, Kirsch S, Karch D, Rappold GA (2002) The novel Rho-GTPase activating gene MEGAP/srGAP3 has a putative role in severe mental retardation. Proc Natl Acad Sci USA 99:11754–11759
Garcia-Mata R, Wennerberg K, Arthur WT, Noren NK, Ellerbroek SM, Burridge K (2006) Analysis of activated GAPs and GEFs in cell lysates. Methods Enzymol 406:425–437
Govek EE, Newey SE, Van Aelst L (2005) The role of the Rho GTPases in neuronal development. Gene Dev 19:1–49
Guerrier S, Coutinho-Budd J, Sassa T, Gresset A, Jordan NV, Chen K, Jin WL, Frost A, Polleux F (2009) The F-BAR domain of srGAP2 induces membrane protrusions required for neuronal migration and morphogenesis. Cell 138:990–1004
Hivert B, Liu Z, Chuang CY, Doherty P, Sundaresan V (2002) Robo1 and Robo2 are homophilic binding molecules that promote axonal growth. Mol Cell Neurosci 21:534–545
Hsieh J, Nakashima K, Kuwabara T, Mejia E, Gage FH (2004) Histone deacetylase inhibition-mediated neuronal differentiation of multipotent adult neural progenitor cells. Proc Natl Acad Sci USA 101:16659–16664
Ito M, Jameson JL, Ito M (1997) Molecular basis of autosomal dominant neurohypophyseal diabetes insipidus. Cellular toxicity caused by the accumulation of mutant vasopressin precursors within the endoplasmic reticulum. J Clin Invest 99:1897–1905
Itoh T, Erdmann KS, Roux A, Habermann B, Werner H, De Camilli P (2005) Dynamin and the actin cytoskeleton cooperatively regulate plasma membrane invagination by BAR and F-BAR proteins. Dev Cell 9:791–804
Jin WL, Liu YY, Liu HL, Yang H, Wang Y, Jiao XY, Ju G (2003) Intraneuronal localization of Nogo-A in the rat. J Comp Neurol 458:1–10
Koh CG (2006) Rho GTPases and their regulators in neuronal functions and development. Neurosignals 15:228–237
Kozma R, Sarner S, Ahmed S, Lim L (1997) Rho family GTPases and neuronal growth cone remodelling: relationship between increased complexity induced by Cdc42Hs, Rac1, and acetylcholine and collapse induced by RhoA and lysophosphatidic acid. Mol Cell Biol 17:1201
Lanning CC, Daddona JL, Ruiz-Velasco R, Shafer SH, Williams CL (2004) The Rac1 C-terminal polybasic region regulates the nuclear localization and protein degradation of Rac1. J Biol Chem 279:44197
Lessard J, Wu JI, Ranish JA, Wan M, Winslow MM, Staahl BT, Wu H, Aebersold R, Graef IA, Crabtree GR (2007) An essential switch in subunit composition of a chromatin remodeling complex during neural development. Neuron 55:201–215
Marillat V, Cases O, Nguyenf-Ba-Charvet KT, Tessier-Lavigne M, Sotelo C, Chedotal A (2001) Spatiotemporal expression patterns of slit and robo genes in the rat brain. J Comp Neurol 442:130–155
Mattar P, Britz O, Johannes C, Nieto M, Ma L, Rebeyka A, Klenin N, Polleux F, Guillemot F, Schuurmans C (2004) A screen for downstream effectors of Neurogenin2 in the embryonic neocortex. Dev Biol 273:373–389
Moon SY, Zang H, Zheng Y (2003) Characterization of a brain-specific Rho GTPase-activating protein, p200RhoGAP. J Biol Chem 278:4151–4159
Nagaraja GM, Kandpal RP (2004) Chromosome 13q12 encoded Rho GTPase activating protein suppresses growth of breast carcinoma cells, and yeast two-hybrid screen shows its interaction with several proteins. Biochem Biophys Res Commun 313:654–665
Nobes CD, Hall A (1995) Rho, rac, and cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia. Cell 81:53–62
Regan CM (1985) Therapeutic levels of sodium valproate inhibit mitotic indices in cells of neural origin. Brain Res 347:394–398
Ridley AJ, Paterson HF, Johnston CL, Diekmann D, Hall A (1992) The small GTP-binding protein rac regulates growth factor-induced membrane ruffling. Cell 70:401–410
Schubert D, Humphreys S, Baroni C, Cohn M (1969) In vitro differentiation of a mouse neuroblastoma. Proc Natl Acad Sci USA 64:316–323
Seo S, Richardson GA, Kroll KL (2005) The SWI/SNF chromatin remodeling protein Brg1 is required for vertebrate neurogenesis and mediates transactivation of Ngn and NeuroD. Development 132:105–115
Soderling SH, Binns KL, Wayman GA, Davee SM, Ong SH, Pawson T, Scott JD (2002) The WRP component of the WAVE-1 complex attenuates Rac-mediated signalling. Nat Cell Biol 4:970–975
Soderling SH, Guire ES, Kaech S, White J, Zhang F, Schutz K, Langeberg LK, Banker G, Raber J, Scott JD (2007) A WAVE-1 and WRP signaling complex regulates spine density, synaptic plasticity, and memory. J Neurosci 27:355–365
Tribioli C, Droetto S, Bione S, Cesareni G, Torrisi MR, Lotti LV, Lanfrancone L, Toniolo D, Pelicci P (1996) An X chromosome-linked gene encoding a protein with characteristics of a rhoGAP predominantly expressed in hematopoietic cells. Proc Natl Acad Sci USA 93:695–699
Tsujita K, Suetsugu S, Sasaki N, Furutani M, Oikawa T, Takenawa T (2006) Coordination between the actin cytoskeleton and membrane deformation by a novel membrane tubulation domain of PCH proteins is involved in endocytosis. J Cell Biol 172:269–279
Vogt DL, Gray CD, Young WS III, Orellana SA, Malouf AT (2007) ARHGAP4 is a novel RhoGAP that mediates inhibition of cell motility and axon outgrowth. Mol Cell Neurosci 36:332–342
Waltereit R, Kautt S, Bartsch D (2008) Expression of MEGAP mRNA during embryonic development. Gene Expr Patterns 8:307–310
Wong K, Ren XR, Huang YZ, Xie Y, Liu G, Saito H, Tang H, Wen L, Brady-Kalnay SM, Mei L, Wu JY, Xiong WC, Rao Y (2001) Signal transduction in neuronal migration roles of GTPase activating proteins and the small GTPase Cdc42 in the Slit-Robo pathway. Cell 107:209–221
Wu G, Vaswani KK, Lu ZH, Ledeen RW (1990) Gangliosides stimulate calcium flux in neuro 2A cells and require exogenous calcium for neuritogenesis. J Neurochem 55:484–491
Wu J, Lessard J, Olave IA, Qiu Z, Ghosh A, Graef IA, Crabtree GR (2007) Regulation of dendritic development by neuron-specific chromatin remodeling complexes. Neuron 56:94–108
Yang Y, Marcello M, Endris V, Saffrich R, Fischer R, Trendelenburg MF, Sprengel R, Rappold G (2006) MEGAP impedes cell migration via regulating actin and microtubule dynamics and focal complex formation. Exp Cell Res 312:2379–2393
Yao Q, Jin WL, Wang Y, Ju G (2008) Regulated shuttling of Slit-Robo-GTPase activating proteins between nucleus and cytoplasm during brain development. Cell Mol Neurobiol 28:205–221
Yoo AS, Crabtree GR (2009) ATP-dependent chromatin remodeling in neural development. Curr Opin Neurobiol 19:120–126
Yuan X, Jin M, Xu X, Song Y, Wu C, Poo M, Duan S (2003) Signalling and crosstalk of Rho GTPases in mediating axon guidance. Nat Cell Biol 5:1–8
Yuan BZ, Jefferson AM, Millecchia L, Popescu NC, Reynolds SH (2007) Morphological changes and nuclear translocation of DLC1 tumor suppressor protein precede apoptosis in human non-small cell lung carcinoma cells. Exp Cell Res 313:3868–3880
Zeng M, Zhou JN (2008) Roles of autophagy and mTOR signaling in neuronal differentiation of mouse neuroblastoma cells. Cell Signal 20:659–665
Acknowledgments
We thank Dr. Yuan XB (Institute of Neuroscience, SIBS, CAS, Shanghai, China) for the gift of GFP-tagged CA-Rac Q61L, CA-Rho Q63L, CA-Cdc42 Q61L, and DN-Rac T17N constructs and Dr. Burridge K (UNC, Chapel Hill, NC) for the gift of GST-Rho GTPases (CA-Rac Q61L, CA-Rho Q63L, and CA-Cdc42 Q61L) constructs. We are grateful for the generous donation of the human KIAA0411 and KIAA 0456 clones from the Kazusa DNA Research Institute (Chiba, Japan). This work was supported by National Natural Science Foundation of China (No. 30770671 and No. 309700936) and Shanghai Leading Academic Discipline Project (No. B205).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
10571_2011_9664_MOESM1_ESM.tif
Supplementary Fig. 1. Expression of srGAPs and their upstream regulators in Neuro2A cells. RT–PCR is performed on total RNA from Neuro2A cells. The specific primers used to detect expression of Slit2, Robo1, Robo2, Robo3, srGAP1, srGAP2, srGAP3, GAPDH and β-actin in Neuro2A cells, the corresponding product size are listed in Table 1. The PCR products are subjected to a 1.5% agarose gel and visualized by ethidium bromide staining. (TIFF 1050 kb)
10571_2011_9664_MOESM2_ESM.tif
Supplementary Fig. 2. Quantification of the differentiation rates of control or srGAP3-knockdown Neuro2A cells cultured in DMEM containing 2% FBS and 20 μM Retinoic Acid. Values are means ± SEM, * p < 0.05. (TIFF 1088 kb)
Rights and permissions
About this article
Cite this article
Chen, K., Mi, YJ., Ma, Y. et al. The Mental Retardation Associated Protein, srGAP3 Negatively Regulates VPA-Induced Neuronal Differentiation of Neuro2A Cells. Cell Mol Neurobiol 31, 675–686 (2011). https://doi.org/10.1007/s10571-011-9664-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10571-011-9664-7